
Abstract
There are three isoforms of arachidonate 12-lipoxygenase in mammals: platelet, leukocyte, and epidermal types. We found in this study that the leukocyte-type enzyme was present in rat pineal gland, lung, spleen, aorta, adrenal gland, spinal cord, and pancreas, as assessed by RT-PCR. Immunohistochemical analysis showed that the enzyme was localized in macrophages in lung and spleen, α-cells of pancreatic islet, zona glomerulosa cells of adrenal cortex, and neuronal cells of spinal cord and superior cervical ganglion. The presence of the 12-lipoxygenase in pancreatic α-cells was confirmed by glucagon staining in a consecutive section. We overexpressed the leukocyte-type 12-lipoxygenase cDNA in a glucagon-secreting αTC clone 6 cell line that had been established from a transgenic mouse. Glucagon secretion was stimulated by approximately twofold in the 12-lipoxygenase-expressing cells compared to the mock-transfected and original cells. The results suggest that the 12-lipoxygenase of the leukocyte type augments glucagon secretion from pancreatic islets.
12-L
Materials and Methods
RNA Isolation
RNA from various rat tissues was isolated using ISOGEN (Nippon Gene; Osaka, Japan) according to the manufacturer's instructions. The tissues (0.5 g), except for pancreas, were homogenized in 5 ml of ISOGEN using a Potter homogenizer at 4C. To prevent RNA degradation, pancreas was frozen in liquid N2 immediately after dissection, crushed with a Polytron homogenizer (Kinematica; Lucerne, Switzerland), and mixed with 10 volumes of ISOGEN. The homogenate was mixed with 1 ml of chloroform, and RNA in the aqueous phase was precipitated with the same volume of isopropyl alcohol.
RT-PCR
To remove contaminating chromosomal DNA, the RNA preparation was incubated at room temperature (RT) for 15 min with 2 U of deoxyribonuclease I (GIBCO BRL; Gaithersburg, MD). DNase was inactivated by heating at 65C for 15 min. The RNA (5 μg) was incubated at 42C for 50 min in a reaction mixture of 20 μl containing 100 U of reverse transcriptase (Super Script II; GIBCO BRL), 20 pmol of oligo (dT)12–18, 50 mM Tris-HCl, pH 8.3, 75 mM KCl, 3 mM MgCl2, 0.5 mM dNTP, 10 mM dithiothreitol, and 10 U RNase inhibitor (Boehringer Mannheim; Mannheim, Germany). PCR primers were designed according to the reported sequence of rat pineal 12-lipoxygenase (Hada et al. 1994): sense 5′-AGCTGGAGCCTTTCTGACCTATTG-3′ and antisense 5′-ACTGATTAGGGTTGGGCAGTGTAG-3′. They were labeled with [γ-32P]-ATP (Amersham Pharmacia Biotech; Uppsala, Sweden) by T4 polynucleotide kinase (Toyobo; Osaka, Japan). PCR was performed in a mixture of 20 μl containing 2 μl of the first reaction mixture, 0.5 U of Ex-Taq DNA polymerase (Takara; Kyoto, Japan), 0.2 mM dNTP, and 0.2 μM each of the radiolabeled primers (104 cpm). Amplification was carried out on a DNA thermal cycler (Perkin-Elmer biosystems; Foster City, CA): denaturation (94C, 30 sec), annealing (65C, 1 min), and extension (72C, 1 min) for 21 cycles. Half of the reaction mixture was electrophoresed on a 7% polyacrylamide gel, and radioactivity was quantified by a Fujix BAS1000 imaging analyzer (Fujifilm; Tokyo, Japan). Quantification was performed in the log-linear phase of the PCR.
Western Blotting for 12-Lipoxygenase
Pineal glands of adult male Wistar rats were homogenized by a Potter homogenizer and platelets were sonicated in 20 mM Tris-HCl buffer, pH 7.4 containing 1 mM EDTA. After centrifugation at 10,000 × g for 10 min, supernatant (10 μg of protein) was subjected to SDS-PAGE followed by the electrophoretic transfer of proteins to a nitrocellulose membrane. The membrane was incubated with an antiserum (1:2000 dilution) that had been raised in rabbit with purified recombinant rat pineal 12-lipoxygenase expressed in E. coli (Kawajiri et al. 1997). After incubation of the membrane with peroxidase-labeled goat anti-rabbit IgG (1:2000 dilution; Vector, Burlingame, CA), 12-lipoxygenase bands were visualized by an ECL system as described previously (Kawajiri et al. 1997). Protein concentration was determined by the method of Lowry et al. (1951) using bovine serum albumin as a standard.
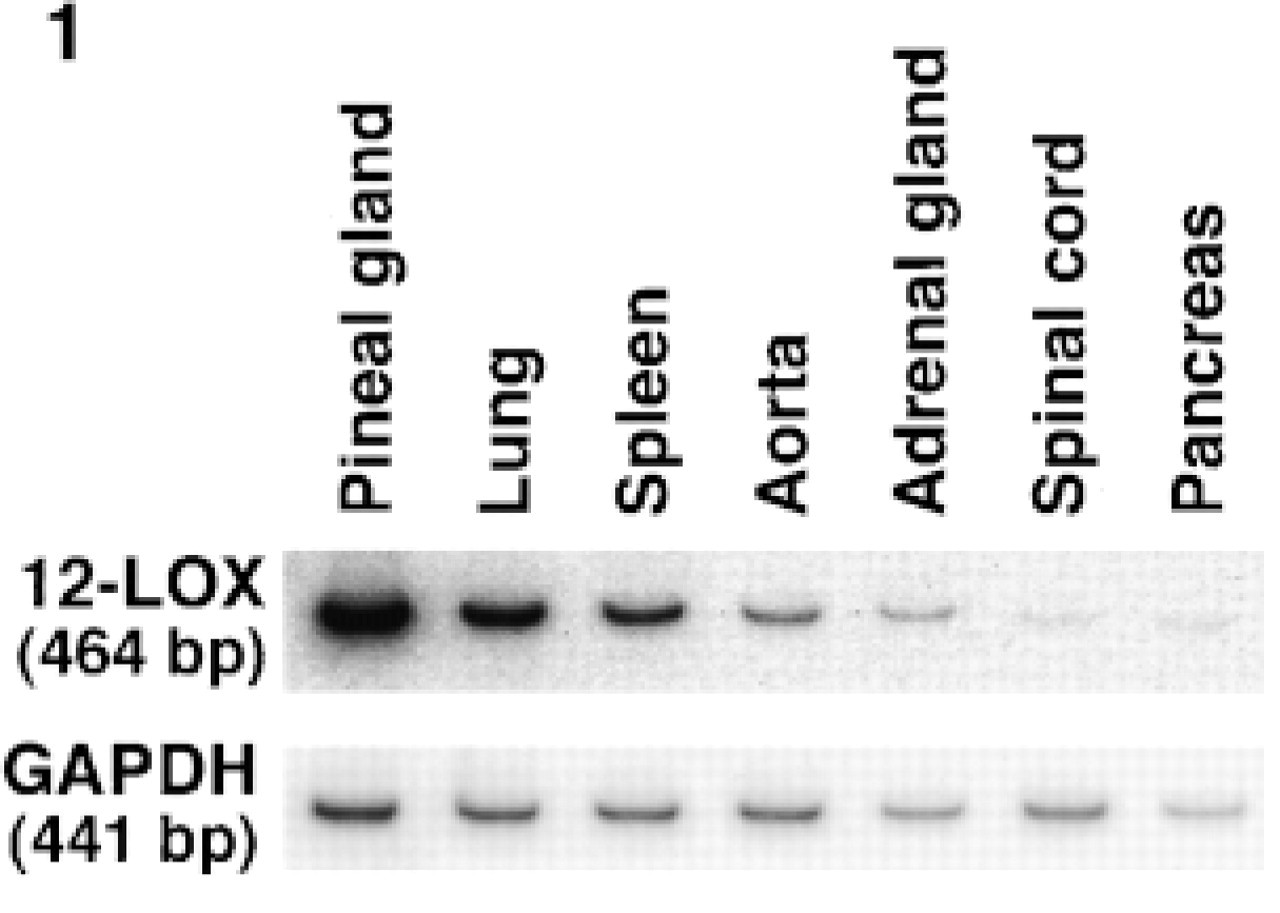
Expression of 12-lipoxygenase mRNA in various rat tissues. Total RNA (2 μg) was subjected to RT-PCR with 32P-labeled primers for rat pineal 12-lipoxygenase (12-LOX) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Amplified DNA resolved by polyacrylamide gel electrophoresis was analyzed by a Fujix BAS1000.

Crossreactivity of anti-rat pineal 12-lipoxygenase antibody. Homogenate (10 μg of protein) of rat pineal glands (Lane b) and platelets (Lane c) was subjected to Western blotting with an ECL detection system. Purified recombinant rat 12-lipoxygenase (0.1 μg of protein) was also run as a control (Lane a) and was slightly retarded due to N-terminal histidines in the expression vector (Kawajiri et al. 1997).
Immunohistochemistry
Adult male Wistar rats were fixed by transcardial perfusion with 4% paraformaldehyde in 0.1 M phosphate buffer, pH 7.2. Various organs and tissues were excised, and immersed in the same fixative at 4C for 4 hr. To make paraffin sections for light microscopy, the specimens were dehydrated in a graded ethanol–xylene series and embedded in paraffin at 60C. To make cryosections for light microscopy and preembedding immunoelectron microscopy, the specimens were rinsed at 4C overnight with 30% sucrose-phosphate buffer, and then frozen at −30C. For postembedding immunoelectron microscopy, the specimens were cut into small pieces, dehydrated in a graded ethanol series, and embedded in LR White resin (London Resin; London, UK) in the presence of 1% benzoyl peroxide. They were then polymerized at RT for 1–2 days with UV polymerizer.
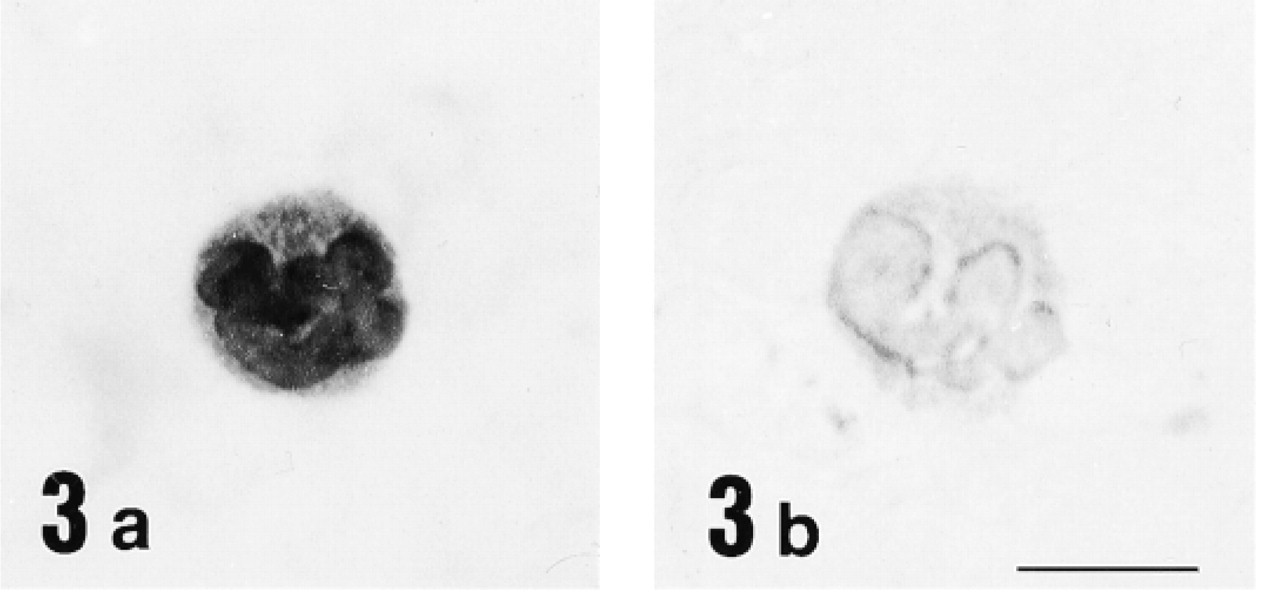
Light micrographs of the peripheral blood immunostained with anti-12-lipoxygenase antiserum (
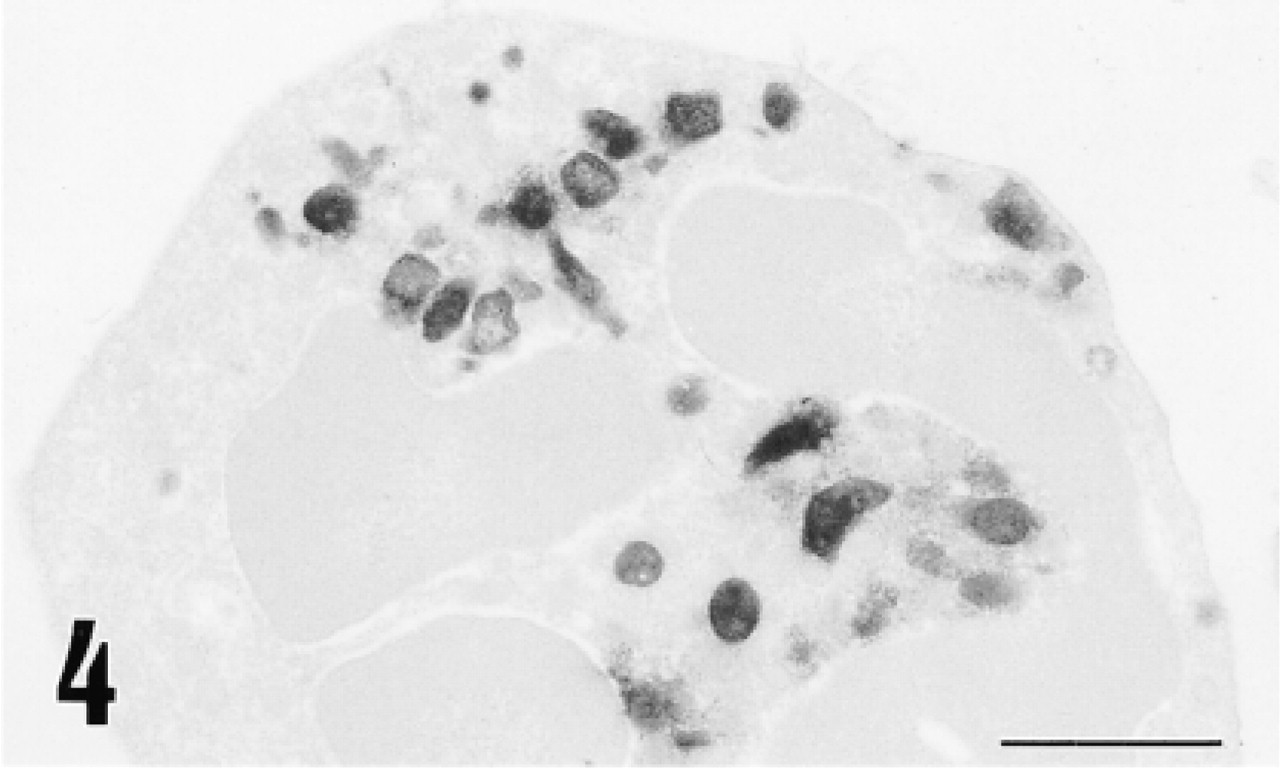
Electron micrograph of a neutrophil immunostained for 12-lipoxygenase by the preembedding (immunoperoxidase) method. Immunoreactive material is localized primarily in the granules. Bar = 1 μm.
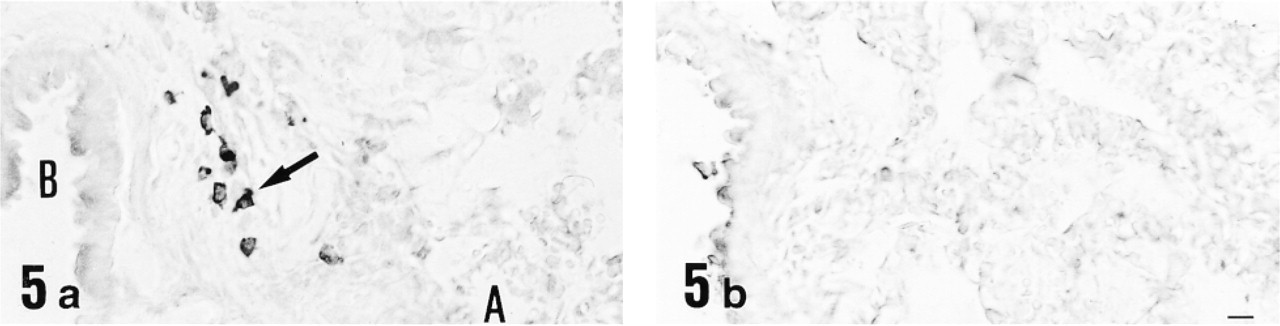
Light micrographs of lung immunostained with anti-12-lipoxygenase antiserum (
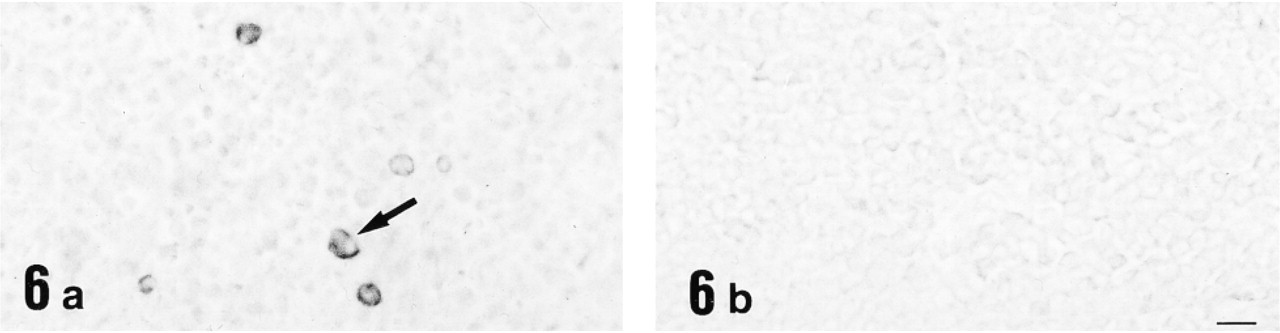
Light micrographs of spleen immunostained with anti-12-lipoxygenase antiserum (
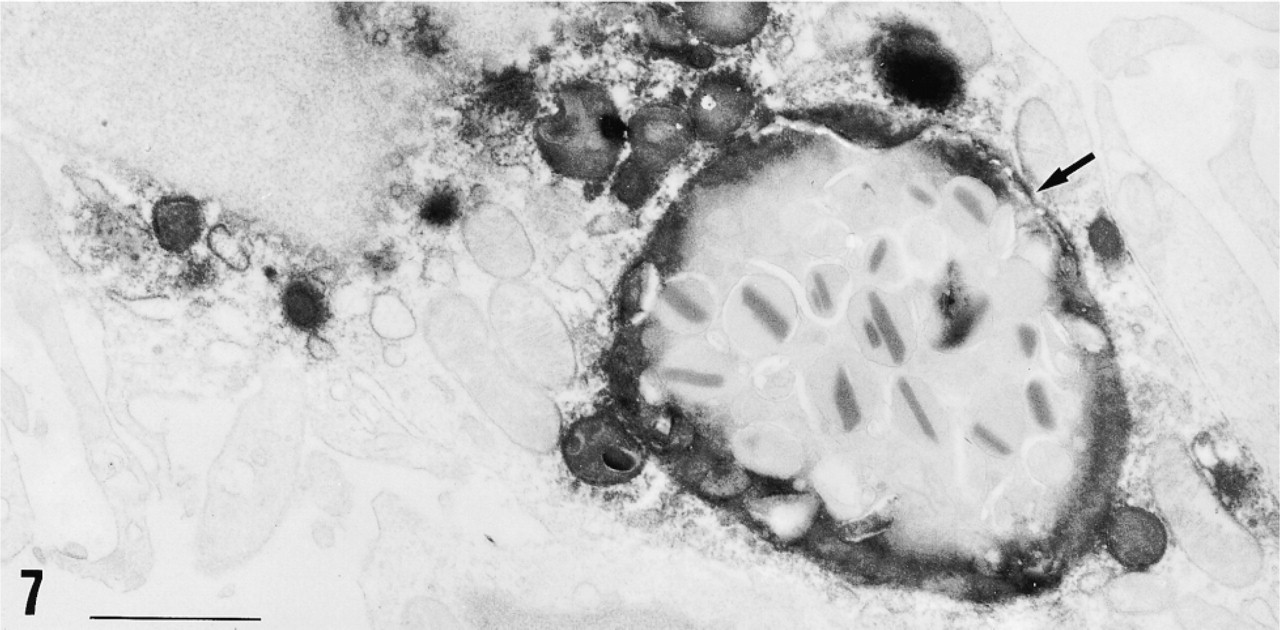
Electron micrograph of the red pulp of spleen immunostained for 12-lipoxygenase by the preembedding (immunoperoxidase) method. The immunopositive cell is identified as a macrophage by a large phagosome (arrow) that is digesting an eosinophil in this case. The immunoreactive material is present in the cytoplasm and is localized primarily in the lysosomes and phagosomes. Bar = 1 μm.
For light microscopic immunohistochemistry, the paraffin-embedded specimens were cut into 4-μm sections with a microtome, mounted on gelatin-coated slides, and deparaffinized in xylene-graded ethanol series. The frozen specimens were cut into 15-μm sections with a cryostat, mounted on slides, and air-dried. Peripheral blood films were also made and fixed with 4% paraformaldehyde. The tissue sections and blood films were treated successively with 0.3% Triton X-100 in 0.1 M PBS, pH 7.2, for 1 hr, with 0.3% hydrogen peroxide in methanol for 10 min, and with 5% normal swine serum (Dakopatts; Glostrup, Denmark) for 30 min. They were then incubated at RT overnight with either the anti-12-lipoxygenase antibody (1:2000 dilution) or rabbit polyclonal anti-glucagon antibody (1:4000 dilution) purchased from Affiniti Research Products (Mamhead, UK). After washing with PBS, the immunoreaction was visualized by incubating the sections successively with biotinylated anti-rabbit IgG (1:200 dilution; Vector Laboratories) for 1 hr, with streptavidin-conjugated horseradish peroxidase (1:500 dilution; Dakopatts) for 1 hr, and finally with a mixture of 0.01% 3′-3′-diaminobenzidine tetrahydrochloride and 0.02% hydrogen peroxide in 50 mM Tris-HCl buffer, pH 7.5. Comparison of 12-lipoxygenase-immunoreactive cells with glucagon-immunoreactive cells was performed in consecutive paraffin sections. For negative controls, either pre- immune rabbit serum or anti-12-lipoxygenase antiserum that had been absorbed overnight with purified 12-lipoxygenase at 50 μg/ml was used as the primary antibody. In some experiments, the antiserum was absorbed with glucagon (Calviochem-Novabiochem; Laufelfingen, Switzerland) at 50 μg/ml.
For electron microscopic immunocytochemistry with the preembedding (immunoperoxidase) method, the immunostained cryosections were postfixed with 0.5% osmium tetraoxide for 20 min, stained with 0.5% uranyl acetate for 30 min, dehydrated in a graded ethanol series, and embedded in epoxy resin (Glicidether 100; Selva Feinbiochemica, Heidelberg, Germany). Ultrathin sections were made and examined with a Hitachi H-700 electron microscope (Tokyo, Japan). For electron microscopic immunocytochemistry with the postembedding (immunogold) method, ultrathin sections were cut from the LR White-embedded specimens and collected on nickel grids. They were first treated with 2% bovine serum albumin in PBS for 15 min and then incubated with anti-12-lipoxygenase antiserum at RT overnight. After washing with PBS, the sections were subjected to the reaction with colloidal gold (20 nm)-conjugated anti-rabbit IgG (British BioCell; Cardiff, UK) at 1:20 dilution in PBS containing 0.1% bovine serum albumin at RT for 2 hr. They were then contrasted with urayl acetate and examined with the electron microscope.
Expression of 12-Lipoxygenase in αTC Clone 6 Cell Line
The full-length cDNA encoding rat pineal 12-lipoxygenase (Hada et al. 1994) was ligated to a eukaryotic pRC/CMV expression vector (Invitrogen; Carlsbad, CA). The expression vector was introduced by the Lipofectin method (GIBCO BRL) into αTC clone 6 cells that had been established from a glucagonoma arising in transgenic mouse (Hamaguchi and Leiter 1990; Powers et al. 1990). The transfected cells were cultured at 37C for 48 hr in Dulbecco's modified minimal essential medium supplemented with 5% fetal bovine serum and 16.5 mM glucose in an atmosphere of 5% CO2. The cells were kept for 14 days in the medium containing 800 μg/ml geneticin. About 100 clones were examined by Western blotting for 12-lipoxygenase expression. Five clones were used in subsequent experiments that had shown the highest enzymatic activity as determined by a method described elsewhere (Hada et al. 1994). Mock-transfected cells were established by transfection of parental plasmids.
Glucagon Secretion
The 12-lipoxygenase-expressing αTC clone 6 cells were cultured for 3 days at an initial density of 107/10 ml in 100-mm dishes. After washing the cells with Dulbecco's PBS, they were cultured in medium containing 80 μg/ml aprotinin to prevent possible degradation of secreted glucagon. Aliquots of 200 μl at various time intervals were subjected to double-antibody radioimmunoassay using a glucagon kit (Daiichi Radioisotope Lab; Tokyo, Japan). Statistical analysis was performed by Student's t-test and a value of p < 0.05 was considered significant.
Results
Expression of 12-Lipoxygenase mRNA in Various Rat Tissues
We examined expression of leukocyte-type 12-lipoxygenase mRNA in various rat tissues by semiquantitative RT-PCR with glyceraldehyde 3-phosphate dehydrogenase as an internal standard. As shown in Figure 1, pineal gland showed by far the highest expression of the enzyme, followed by lung, spleen, aorta and adrenal gland. Faint bands were observed in spinal cord and pancreas. These bands were not detected in the absence of reverse transcriptase, and the primers were designed to span three introns of the enzyme, indicating that only 12-lipoxygenase cDNA was amplified under our experimental conditions.
Specificity of Antibody against Rat Pineal 12-Lipoxygenase
Before immunohistochemistry, the specificity of anti-rat pineal 12-lipoxygenase antibody was examined by Western blotting analysis, as shown in Figure 2. Rat pineal glands gave a dense band of about 74 kD. In contrast, rat platelets that showed 12-lipoxygenase activity (Chang et al. 1982) did not give any detectable bands. The result indicates that this antibody specifically binds to the pineal 12-lipoxygenase, which is of the leukocyte type based on amino acid sequence (Hada et al. 1994), but not to the platelet enzyme.
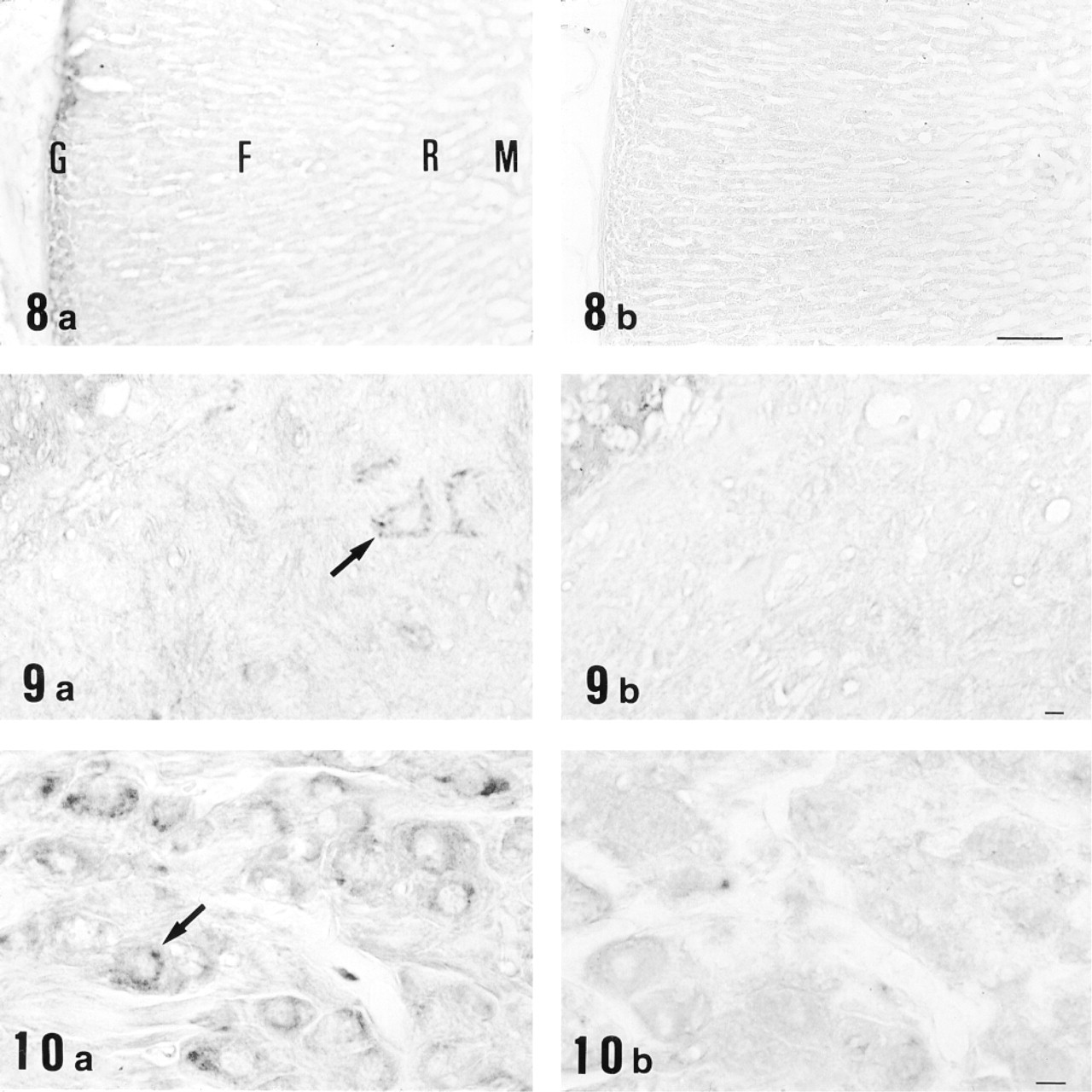
Light micrographs of cryosections of adrenal gland (Figure 8), spinal cord (Figure 9), and superior cervical ganglion (Figure 10) immunostained with anti-12-lipoxygenase antiserum (
Immunohistochemistry for 12-Lipoxygenase
Immunohistochemical study for 12-lipoxygenase was carried out in a variety of rat organs and tissues. In the blood film, neutrophils were strongly immunostained with anti-12-lipoxygenase antibody (Figure 3a). On electron microscopy of neutrophils, the immunoreactive material was present in the cytoplasm and preferentially was localized to the granules (Figure 4). In the cryosections, intense immunoreactivity was also recognized in a subset of connective tissue cells scattered at various locations, such as the walls of bronchi and blood vessels of lung (Figure 5a) and the red pulp of spleen (Figure 6a). When examined with electron microscopy, these immunopositive cells were identified as macrophages, with their typical large phagosomes (Figure 7). The immunoreactive material was present in the cytoplasm of macrophages and preferentially was localized to the lysosomes and phagosomes. In the adrenal cortex, shown in Figure 8a, zona glomerulosa cells were immunostained, whereas the cells of zona fasciculata and reticularis of cortex and those of medulla were not stained. We also examined the immunoreactivity for 12-lipoxygenase in neuronal tissues. The pineal gland was densely immunostained (data not shown). Some neurons, such as anterior horn cells of the spinal cord (Figure 9a) and the ganglion cells of the superior cervical ganglion (Figure 10a), exhibited intense cytoplasmic immunoreactivity that was granular. Incubation with preimmune rabbit serum did not show any staining in these tissues (Figures 3b, 5b, 6b, 8b, 9b, and 10b). Moreover, preabsorption of the antibody with an excess amount of antigen (50 μg/ml) abolished the positive immunostaining (data not shown), indicating the specificity of our anti-12-lipoxygenase antibody.
Light micrographs of serial sections of pancreas immunostained with anti-12-lipoxygenase antiserum (
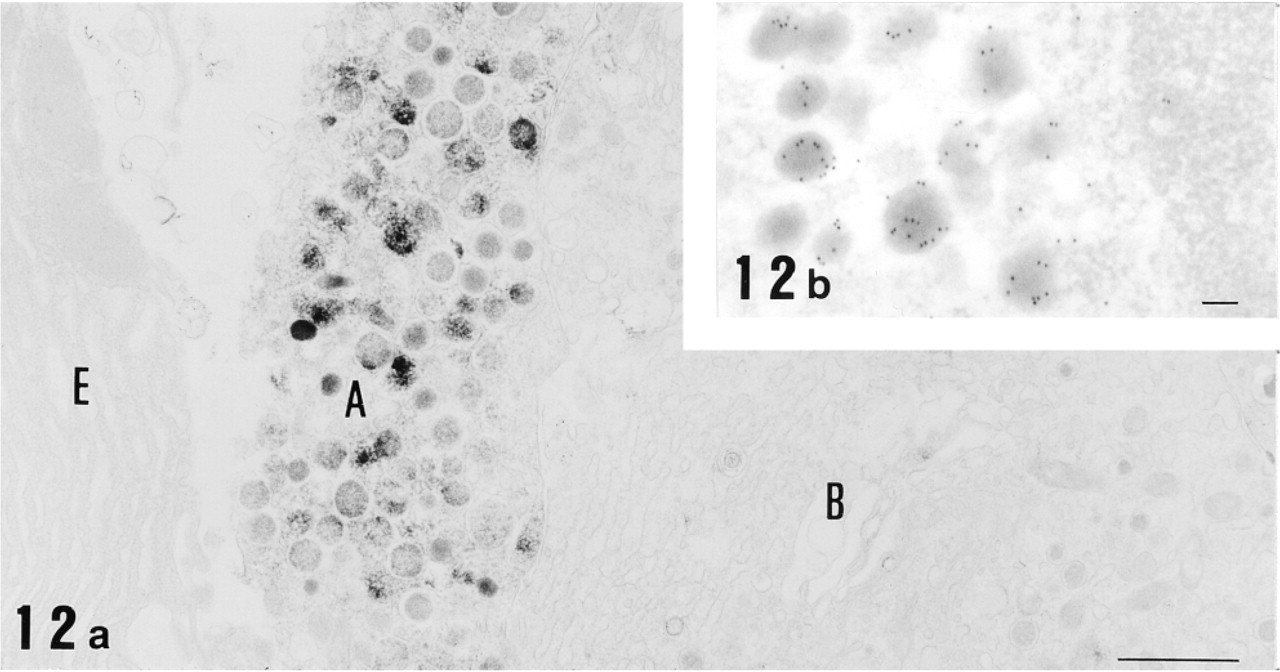
Electron micrographs of pancreas immunostained for 12-lipoxygenase by the preembedding (immunoperoxidase) method (
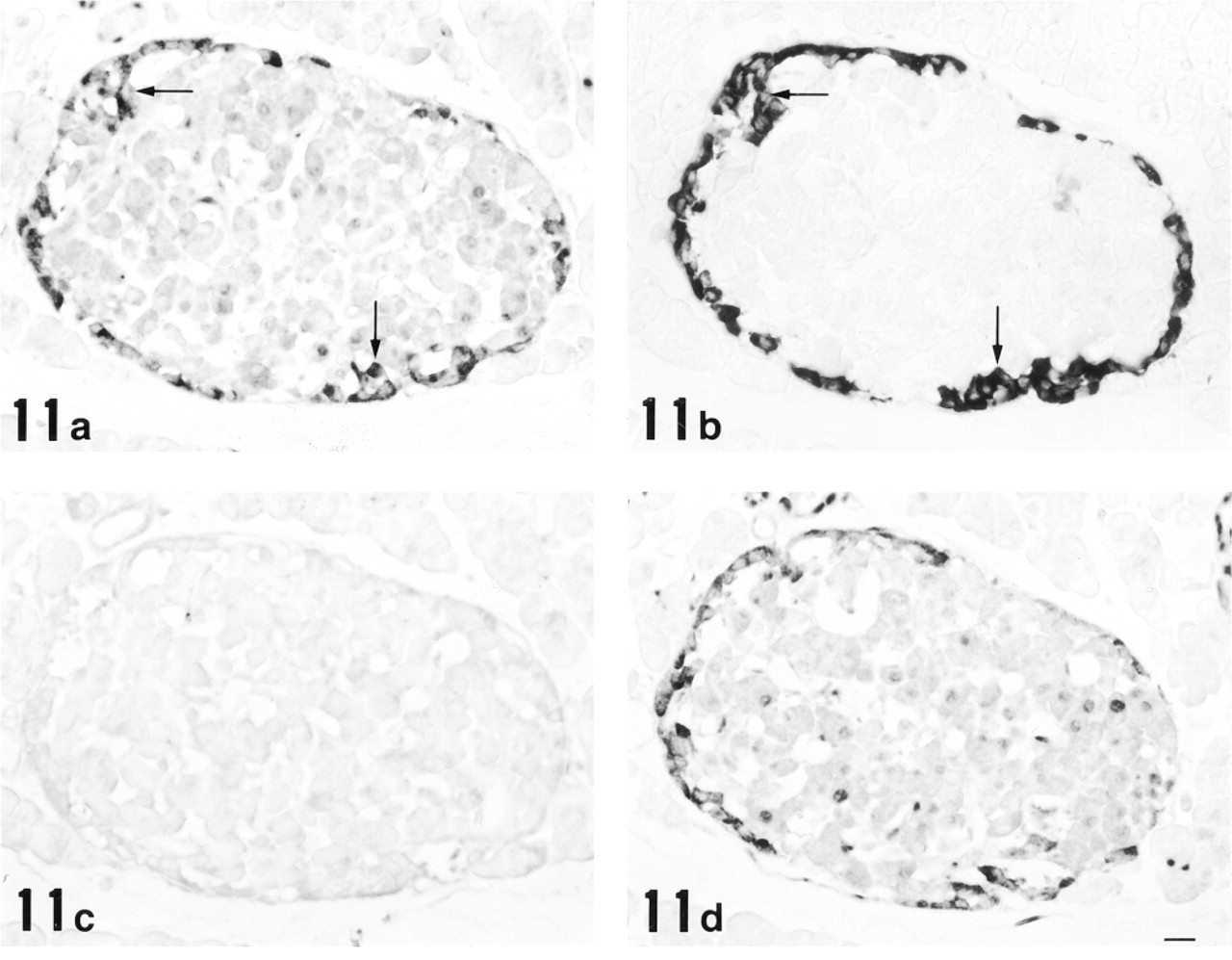
Finally, we found strong immunoreactivity in a subset of endocrine cells preferentially located at the periphery of pancreatic islets. Consecutive paraffin sections incubated with anti-12-lipoxygenase and anti-glucagon antisera clearly demonstrated the staining of identical cell populations (Figures 11a and 11b), indicating that 12-lipoxygenase immunoreactivity belongs to glucagon-secreting α-cells. All α-cells appeared to be immunopositive for 12-lipoxygenase. To further confirm the antigen specificity of α-cell staining, preabsorption of the antisera with purified antigens was performed. As shown in Figure 11c, absorption of anti-12-lipoxygenase antiserum with 12-lipoxygenase completely abolished the immunoreaction. In contrast, absorption of anti-12-lipoxygenase antiserum with glucagon caused no suppression of the immunoreaction (Figure 11d), despite the fact that the same concentration of glucagon completely abolished the immunoreaction for anti-glucagon antiserum (data not shown). These results indicate that our anti-12-lipoxygenase antiserum specifically recognizes 12-lipoxygenase and does not crossreact with glucagon in α-cells. Subcellular localization of 12-lipoxygenase was then examined with immunoelectron microscopy. Using the preembedding (immunoperoxidase) method, the immunoreactive material appeared to be present in the cytoplasm of α-cells and localized primarily in the endocrine granules (Figure 12a). With the postembedding method, the immunoreactivity was further localized preferentially in the periphery of each endocrine granule (Figure 12b).
Glucagon Secretion from an α-like Cell Line
Because 12-lipoxygenase was present in pancreatic α-cells, we examined whether the enzyme would affect glucagon secretion. We transfected the leukocyte-type 12-lipoxygenase cDNA into a glucagon-secreting αTC clone 6 cell line. Five 12-lipoxygenase-expressing clones were selected, which produced 0.2–0.6 nmol of 12-hydroxy eicosatetraenoic acid/107 cells/10 min from arachidonic acid. There were no detectable enzyme activities in the original and mock-transfected cells. As shown in Figure 13a, the glucagon secretion from original αTC clone 6 cells markedly increased for the first 30–60 min and gradually increased thereafter. The 12-lipoxygenase-expressing cells released approximately twice as much glucagon as original and mock-transfected cells. As shown in Figure 13b (open bars), cumulative glucagon secretion in the 12-lipoxygenase-expressing cells was significantly greater (1.3 ± 0.3 ng/105 cells/30 min) than that in the mock-transfected cells (0.5 ± 0.2 ng/105 cells/30 min). Arachidonic acid as a substrate of 12-lipoxygenase appeared to increase the glucagon secretion, but the increase in both 12-lipoxygenase and mock-transfected cells was not statistically significant (shaded bars in Figure 13b). These results suggest that the leukocyte-type 12-lipoxygenase per se enhances glucagon secretion by pancreatic α-cells.
Discussion
RT-PCR analysis of leukocyte-type 12-lipoxygenase mRNA showed the predominant presence of the enzyme in pineal gland, followed by lung, spleen, aorta, and adrenal gland. This finding was consistent with previous observations by Northern blotting (Hada et al. 1994) and enzyme activity (Yoshimoto et al. 1984). In addition, the sensitive RT-PCR revealed the enzyme mRNA expression in hitherto unidentified tissues, such as pancreas and spinal cord. We found that 12-lipoxygenase protein was markedly expressed in macrophages in rat lung and spleen (Figures 5 and 6). It should be mentioned that 12-lipoxygenase expression was detected in about 30–40% of resident peritoneal macrophages of mice (Sun and Funk 1996). With regard to the functional significance of the 12-lipoxygenase in macrophages, lipoxygenase-mediated oxidation of low-density lipoprotein has been proposed as a contributing factor to atherogenesis (Parthasarathy et al. 1989; Kuhn et al. 1994). Enhanced oxidation of low-density lipoprotein by zymosan-stimulated macrophages was not observed in 12-lipoxygenase-deficient macrophages, as measured as thiobarbituric-acid reactive substances (Sun and Funk 1996). An essential role of leukocyte-type 12-lipoxygenase in the development of atherosclerosis was also demonstrated in double-knockout mice for the enzyme and apo E (Cyrus et al. 1999).
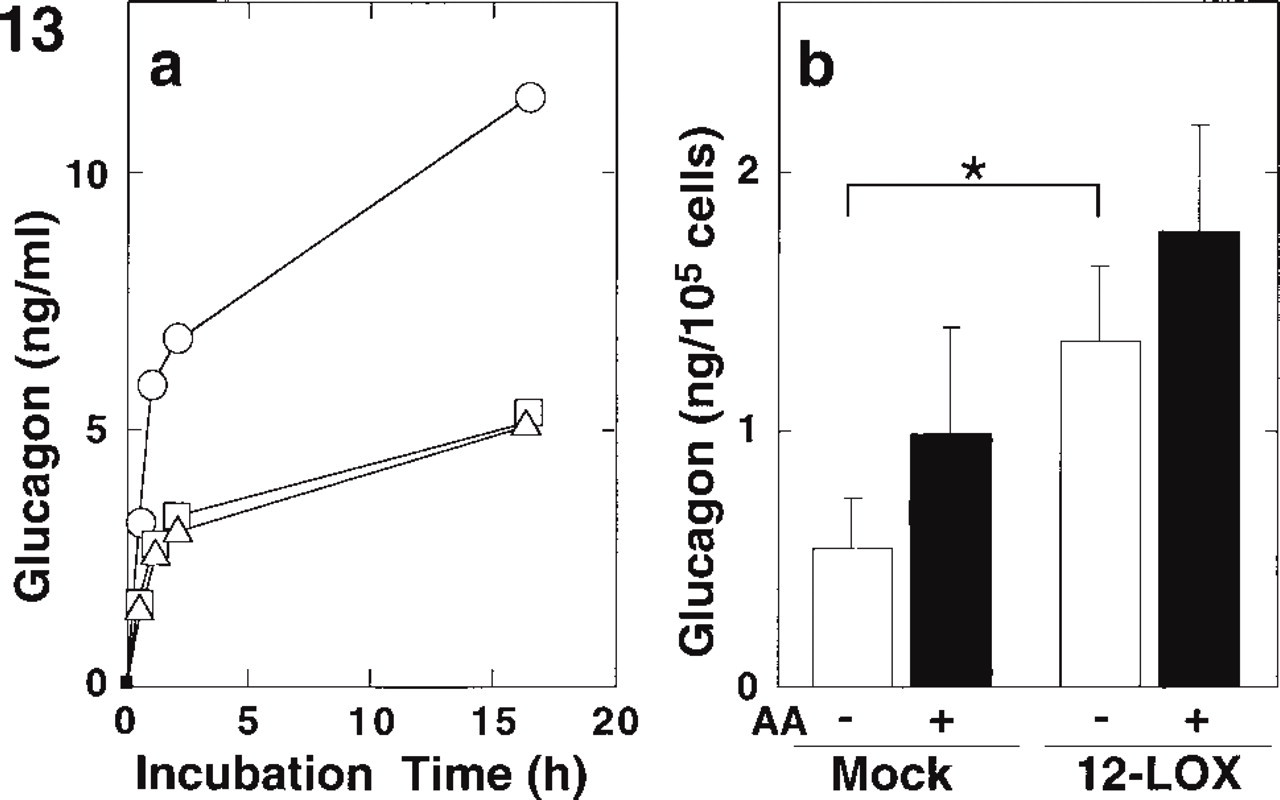
Glucagon secretion of αTC clone 6 cells expressing leukocyte-type 12-lipoxygenase. (
The leukocyte-type 12-lipoxygenase was clearly detected in zona glomerulosa cells of rat adrenal cortex (Figure 8). The presence of the enzyme in human adrenal glomerulosa cells was also demonstrated by in situ hybridization using an antisense riboprobe (Gu et al. 1994). Inhibition of angiotensin II-mediated aldosterone secretion by a selective 12-lipoxygenase blocker suggests that this enzyme plays a key role in mediating aldosterone secretion from adrenal cortex (Nadler et al. 1987; Natarajan et al. 1988).
Interestingly, the leukocyte-type 12-lipoxygenase was detected in nerve cells in spinal cord and superior cervical ganglion (Figures 9 and 10). It was previously shown that the 12-lipoxygenase was found in neurons and glial cells in canine cerebrum, basal ganglia, and hippocampus (Nishiyama et al. 1993). However, we could not detect any immunoreactive cells in rat cerebrum and cerebellum (data not shown). Inconsistency of enzyme localization in the nervous system might be attributable to species differences, which were often seen in this class of enzyme (Yoshimoto and Yamamoto 1995). It is of interest that the 12-lipoxygenase metabolites might act as a second messenger in Aplysia neurons (Piomelli et al. 1987; Buttner et al. 1989). Moreover, 12-lipoxygenase was shown to modulate glutamate release in hippocampal mossy fiber terminals (Freeman et al. 1991; Dorman et al. 1992). Our recent experiments demonstrated that the 12-lipoxy-genase and/or its metabolites upregulated membrane excitability by targeting K+ channels in NG108–15 cells (Takahashi et al. 1999). Therefore, it is envisaged that the leukocyte-type 12-lipoxygenase present in the nerve cells modulates neuronal signaling.
Our immunohistochemical study revealed the presence of 12-lipoxygenase in α-cells of pancreatic islets (Figure 11a). This finding was confirmed by glucagon staining of consecutive sections (Figure 11b). Previous reports showed that a 12-lipoxygenase was present in β-cells of rat pancreatic islet using polyclonal antibodies against porcine leukocyte 12-lipoxygenase (Shannon et al. 1992) and a partial peptide with sequence homology to amino acids 39–55 of the porcine enzyme (Bleich et al. 1995). There are several isoforms of 12-lipoxygenase, and the enzyme localization is different from species to species (Yoshimoto and Yamamoto 1995). Therefore, we raised the antibody against rat 12-lipoxygenase, and examined the tissue distribution in the same species. The difference of 12-lipoxygenase localization in pancreatic islets may be attributable to crossreactivity of the antibodies, which were raised with the purified enzyme of different origins; porcine (previous studies) and rat (this study). A notable finding in this study was the enhanced glucagon secretion in the αTC clone 6 cells overexpressing the leukocyte-type 12-lipoxygenase (Figure 13). However, the addition of 12-hydroxy eicosatetraenoic acid at 3 μM did not increase glucagon secretion from original αTC clone 6 cells under our experimental conditions (unpublished data). It was previously shown that synthetic 12-hydroxy eicosatetraenoic acid at 1 μM increased glucagon release from isolated pancreatic islets by approximately 220% (Falck et al. 1983). Ineffectiveness of the 12-hydroxy acid in our study might be explained by the changes of phenotype, including glucagon secretion, during transformation of the αTC clone 6 cells. Alternatively, because hepoxilin as a metabolite of 12-lipoxygenase pathway was reported to raise insulin levels (Pace-Asciak et al. 1999), other metabolites than 12-hydroperoxy or 12-hydroxy acids might regulate glucagon secretion. The specific localization of the leukocyte-type 12-lipoxygenase in particular cell types of pancreatic islets, adrenal gland, and neural tissues, all of which release hormones or neurotransmitters, suggests that the enzyme plays a certain role in the secretion of these compounds. Further studies to address the specific role of 12-lipoxygenase in glucagon secretion will allow a more comprehensive understanding of the role of this enzyme in hormone secretion.
Footnotes
Acknowledgements
Supported by grants-in-aid from the Ministry of Education, Science and Culture of Japan.