
Abstract
We report the isolation and characterization of a serine/threonine kinase expressed during rat pancreas development. This kinase was cloned as part of a general screen using degenerate oligonucleotides to map expression of kinases and receptors during the course of pancreatic development. Sequence analysis showed it to be a member of the ste20-like serine/threonine kinase family. Northern blotting analysis against both fetal and adult tissues showed two transcripts, one of 2 kb and the other of 4 kb. The ratio of transcript expression varied with the tissue. In situ hybridization analysis showed that this gene is expressed in the early gut and pancreatic epithelium. By embryonic Day 15, the transcript is localized to cells that will eventually become exocrine in nature. In situ hybridization analysis also demonstrated high levels of expression in the choroid plexus, the developing myocardium, kidney, CNS, dorsal root ganglia, and testes. In addition, a search of the EST database revealed a related human kinase not previously described.
P
A number of soluble growth factors can influence the survival, growth, and differentiation of pancreatic epithelium grown in explant culture (Sanvito et al. 1994; Gittes et al. 1996; Miralles et al. 1998). TGF-β1 has been shown to inhibit acinar formation and to increase the number of insulin- and pancreatic polypeptide-expressing cells (Sanvito et al. 1994). In contrast, follistatin has been shown to have an opposite effect, decreasing the number of insulin-positive cells and increasing the number of amylase-expressing exocrine cells (Miralles et al. 1998). EGF has been shown to increase epithelial growth in vitro (Githens et al. 1989), and overexpression of TGFα in transgenic mice has been shown to result in duct hyperplasia (Wang et al. 1993). Despite these studies, little is known about how the pancreas develops and differentiates and how determinations of cell fate are made. Even less is known regarding the role of receptors and signaling molecules during pancreatic development.
To better understand the process of pancreatic development and how choices might be made with regard to endocrine vs exocrine differentiation, we have begun mapping the expression patterns of signaling molecules and receptors during development. As part of this approach, we have used degenerate PCR to detect and map the expression of members of the receptor tyrosine kinase (RTK) and serine/threonine kinase (STK) signaling molecule families. RNA from discrete stages of pancreatic development was prepared and subjected to analysis. In our first series of experiments, a number of known signaling molecules previously unmapped to the pancreas were detected.
We describe here the identification of an STK that is expressed during pancreatic development. This STK is identical to the gene PASK identified by Ushiro et al. (1998). Our work shows that PASK is expressed both during development and in the adult in a tissue-specific manner as two species, a 2-kb transcript and a 4-kb transcript, as determined by Northern blotting analysis. In the adult, this gene is abundantly expressed in testes, kidney, heart, and pancreas, and less abundantly in brain, but is not detected in muscle. In situ hybridization of early embryos demonstrates expression in pancreatic epithelium, myocardium, and choroid plexus, in sensory ganglia such as the dorsal root and cervical ganglia, and in the mesonephros. A time course analysis of expression in the pancreas shows that during fetal development PASK is not expressed in insulin-positive cells but instead localizes to the exocrine parenchyma. Therefore, in the fetal pancreas PASK may be a marker of exocrine differentiation.
Materials and Methods
Cloning and Sequencing of cDNAs
Timed-pregnant Sprague-Dawley rats were obtained from Taconic Labs (Germantown, NY). Appropriately staged embryos were dissected and the pancreases collected on dry ice for RNA and either 2% or 4% paraformaldehyde for histology. Total RNA was obtained by Trizol (Gibco BRL; Rockville, MD) extraction. Each RNA sample was quantitated and stored as an EtOH precipitate at −80C until further use. Reverse transcription was performed using SuperScript II (Gibco). The resulting single-stranded cDNA was then amplified using degenerate primers to the conserved IHRDL and WMAPE kinase motifs (Lai and Lemke 1991). Primer sequences used were as follows: 5′, 5′-GTTAGGATCCACMGNGAYYT3′; 3′: 5′-AGGAATTCCRWAGGACCASACRTC3′. PCR cycling parameters were five cycles of 94C for 1 min, 37C for 2 min, 72C for 1 min, followed by 25 cycles of 94C for 1 min, 50C for 2 min, and 72C for 1 min. The resulting band of 220 bp obtained at all stages of development was cloned into the TA vector system (Clontech; Palo Alto, CA). Plasmid sequencing was performed by Seqwright Technology (Houston, TX). Sequences were assembled from rat sequences and related human EST sequences using the phred/phrad/consed package (Ewing and Green 1998; Gordon et al. 1998). Translated protein sequences were analysed for motifs (Genetics Computer Group; Madison, WI) and aligned with clustalw (Thompson et al. 1998). The 220-bp insert of a novel clone later shown to be PASK was isolated and used to screen rat testis and pancreas cDNA libraries (Stratagene; La Jolla, CA).
Northern Blotting Analysis of PASK mRNA Expression
RNA was isolated from e18 fetal and adult pancreas using the guanidinium isothiocyanate method (Cathala 1983). A multiple-tissue Northern blot was purchased from Clontech. Both the 220-bp fragment encoding pstk1 and a larger 3′ fragment from nucleotides 652–1843 were used as probes for the Northern blotting, with identical results. Probe synthesis using [32P]-dCTP (Amersham; Arlington Heights, IL) and random primer labeling were performed as described by the manufacturer (Gibco). Total RNA run on formaldehyde gels was transferred onto charged nylon membranes (Boehringer Mannheim; Indianapolis, IN) and hybridized in Xpress Hyb solution (Clontech) as described by Dulac and Axel (1995).
Localization of PSTK1 mRNA by Whole-mount and Section In Situ Hybridization Analysis
In situ probes were made using a digoxigenin RNA synthesis kit (Boehringer). Digoxigenin-based whole-mount in situ hybridization was performed as described previously (Miao et al. 1997). A fragment from nucleotides 652–1843 was cloned into the Bluescript vector system (Stratagene) and sense and antisense RNA probes were made by transcribing with T3 and T7 RNA polymerases and labeling with digoxigenin (Boehringer). Digoxigenin-labeled probes were used for both whole-mount and section analysis of staged embryos and tissues. Tissue sections were in some cases counterstained with hematoxylin and/or eosin or, alternatively, photographed with Nomarski optics.
Immunohistochemistry
Guinea pig anti-insulin antibodies were purchased from Linco Research (St Charles, MO). The antibodies were used at a final concentration of 1:2000 on paraffin sections in a PBS-Triton (0.3%) buffer using 1% BSA and 5% normal donkey serum as blocking agent. Secondary antibody was a biotinylated donkey anti-guinea pig antibody purchased from Jackson Immunoresearch (West Grove, PA) and used at a final concentration of 1:500, followed by avidin D-HRP (1:200) from Vector Labs (Burlingame, CA). Antibody binding was visualized using a diaminobenzidine substrate kit (Zymed; South San Francisco, CA).
Results
Isolation and Nucleotide Sequence Analysis of PASK cDNAs
Kinase members were identified by RT-PCR amplification of total RNA isolated from four different stages of rat embryonic pancreatic development spanning e12–e18. Degenerate primers were chosen from conserved nucleotide (nt) sequences between Domains VIII and IX of receptor tyrosine kinases (RTK) (Lai and Lemke 1991). A single band of approximately 220 bp was obtained from each stage studied. Each band was then isolated, cloned, and sequenced. The nt sequence analysis of subcloned RT-PCR products demonstrated that the products encoded a wide array of kinase subfamilies (Table 1).
Twelve different kinases were detected in our study, and the frequency of their detection varied with development (Table 1). However, it is noteworthy that the sample size of the analyzed populations was small and therefore the detected differences in clone frequency observed might be as much due to relative stage-specific cloning efficiencies as reflective of relative expression levels. Nevertheless, we include Table 1 as an example of signaling molecules detected by this methodology and heretofore unmapped to pancreatic development. The stages chosen for analysis span both the early expansion phase of pancreatic development (e12–e14) and the time period between e15 and e18, a time in which the pancreas is undergoing differentiation (Pictet and Rutter 1972).
Interestingly, PDGFR was detected most often in the e12–e14 period, whereas c-abl was found more often in the later differentiative phase. Molecules such as Flk1 showed some differences in detected levels and a few, such as the IGF-1 receptor and JAK1, showed low detectable levels.
One of the cloned RT-PCR products detected in our analysis of e18 cDNA encoded what appeared to be a novel kinase on database comparison using BLAST (Altschul et al. 1990), which subsequently turned out to be the PASK gene published by Ushiro et al. (1998). However, at that time, and on the basis of our discovery of a second related human homologue (see below), our gene sequence was deposited in Genbank as PSTK1 (for pancreas serine threonine kinase 1; accession number AF068261). Subsequent RT-PCR analysis showed that this transcript is present even in the early pancreatic bud (data not shown) but was not pulled out of our stage-specific screen, perhaps because of its much lower relative expression at earlier stages. A multiple-tissue Northern blot of adult rat also showed two transcripts of approximately 2 kb and 4 kb distributed among several tissues (Figure 1A). The Northern blot demonstrated strong expression in the testes and low expression in the kidney, heart, and brain. Expression in, spleen, lung, liver, and skeletal muscle was undetectable. Expression in the testes showed a predominance of the 2-kb band, whereas expression in the heart, brain, and kidney was found predominantly as the 4-kb transcript. Hybridization with an actin probe showed equal amounts of RNA per lane (not shown). To examine pancreatic expression, RNA was collected from both fetal and adult pancreas. Northern blotting analysis showed that both the fetal and adult pancreas express both transcripts in approximately equal ratio (Figure 1B).
Detection of kinases expressed during pancreatic development a
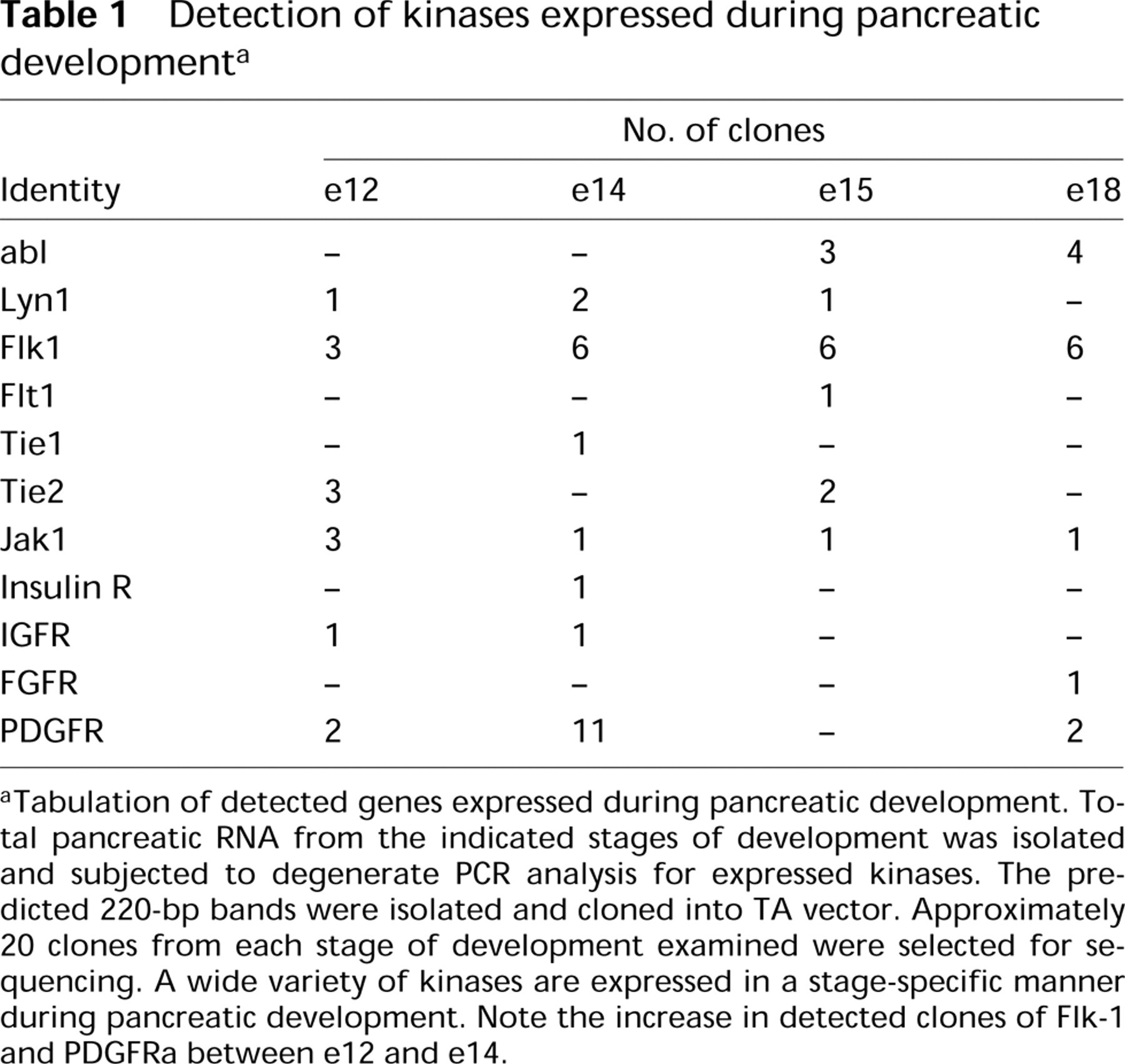
aTabulation of detected genes expressed during pancreatic development. Total pancreatic RNA from the indicated stages of development was isolated and subjected to degenerate PCR analysis for expressed kinases. The predicted 220-bp bands were isolated and cloned into TA vector. Approximately 20 clones from each stage of development examined were selected for sequencing. A wide variety of kinases are expressed in a stage-specific manner during pancreatic development. Note the increase in detected clones of Flk-1 and PDGFRa between e12 and e14.
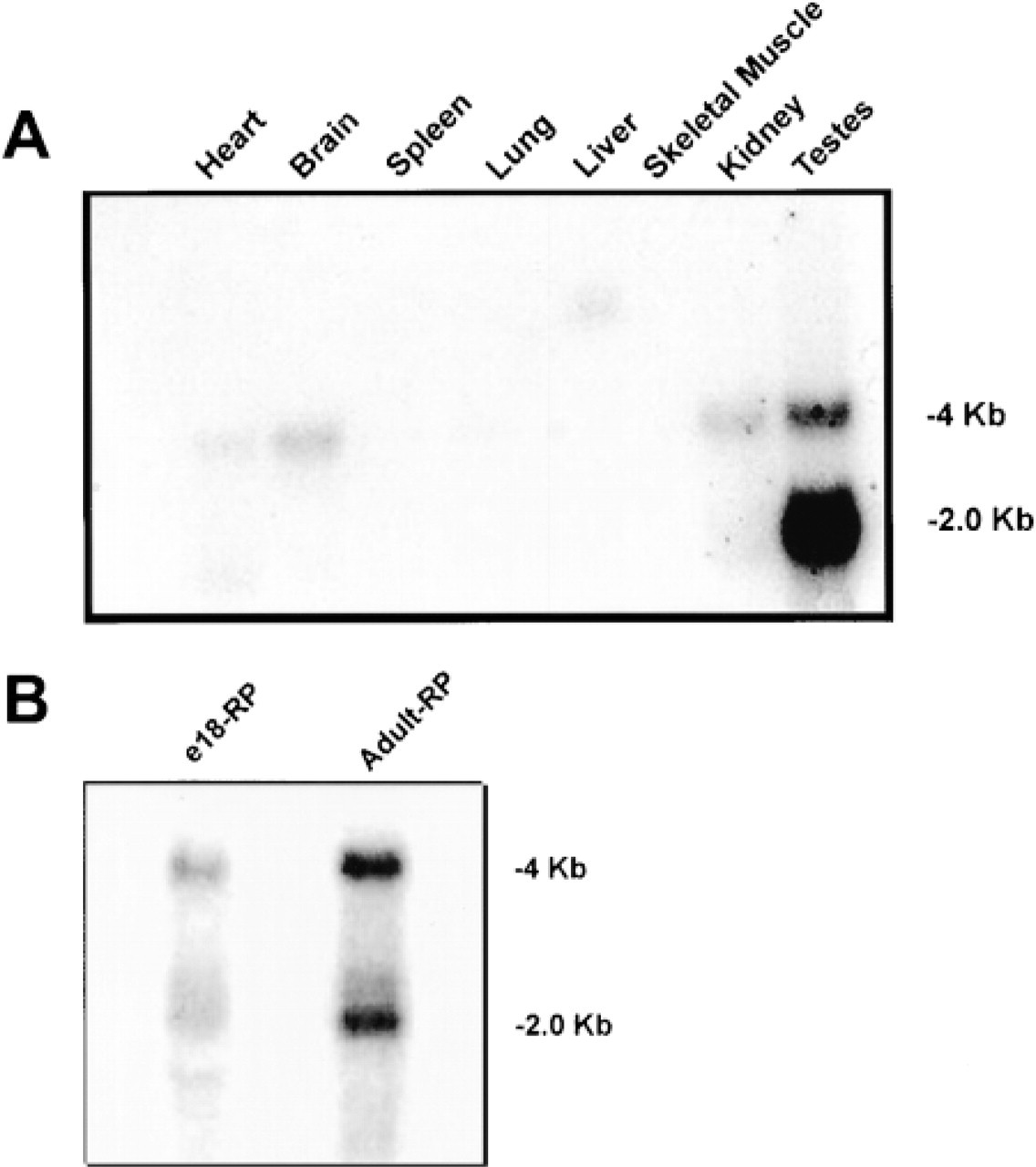
Northern blotting analysis of pask expression. (
On the basis of the expression pattern, we screened both a pancreas and a testis cDNA library and isolated five independent clones, apparently derived from the same mRNA species because no alternative splice forms were found. The longest cDNA clone was 1.86 kb and was designated pstk1, for pancreas serine/threonine kinase one. It contains a predicted 181-bp 5′-untranslated region followed by a 1659-bp open reading frame encoding a protein of 563 amino acids plus a stop codon, leaving a predicted 3′-untranslated region of 104 bp. The polyadenylation site was not found in any of the clones examined. The predicted amino acid sequence of the protein encoded by this clone is shown in Figure 2. Comparison of this sequence to the human EST database revealed the presence of a homologue that is 97.6% identical to the rat protein. Our analysis also discovered a similar but not identical kinase, which we termed human PSTK-2, which is 77.6% identical to the rat PASK/PSTK1 protein. We believe that these two genes might be part of a separate kinase subgroup. Sequence alignments and cladistic analysis (not shown) revealed pask/pstk-1 to be most similar to the Ste20-like family of kinases (Creasy et al. 1996).
Expression in the e14 Rat Embryo
Figure 3 shows a parasagittal section of an e14 rat embryo hybridized with antisense digoxigenin-labeled pask probe. Hybridization was visualized using an alkaline phosphatase-labeled anti-digoxigenin antibody and BM purple (Boehringer). The sections were then counterstained with eosin. Hybridization with sense control showed no staining expression (data not shown). There was a high level of pstk1 expression in the choroid plexus, myocardium, mesonephros, and in the dorsal root ganglia. The choroid plexus, myocardium, and mesonephros, respectively, are shown under higher magnification from top to bottom in the accompanying inset. Expression in the choroid plexus was observed throughout the anterior CNS from telencephalon to myelencephalon. Whole-mount staining also showed strong expression throughout the developing adrenal gland (data not shown).
Expression in the e15 Embryo
Figure 4 is a more medial parasagittal section of an e15 rat embryo. There was continued strong expression of pask in the choroid plexus. In addition, a number of large ganglia in the CNS now showed strong pask expression. Froriep's ganglion, which is a vestigial DRG whose components later break down to give rise to the motor neuron elements of the hypoglossal (XII) ganglion (Kuratani et al. 1988), the vestibulocochlear (VIII) ganglion, and the glossopharyngeal (IX) ganglion, all expressed pstk1. There was also continued expression of pstk1 in the myocardium and dorsal root ganglia.
Pancreatic Expression
Whole-mount in situ hybridization of e12 rat gut tubes showed distinct expression of pstk1 in the early pancreatic epithelium (Figure 5A) and in the epithelium of the developing gut tube (Figure 5B, arrows). The surrounding mesenchyme was negative for pstk1 expression and at this stage the entire pancreatic epithelium appeared to be pask-positive. Figure 5C shows an e12 gut tube hybridized with sense control. Little expression was found in the stomach anlage and no detectable expression was found in the lung. Expression in the gut epithelium could be detected as early as e11 and increased throughout the gut tube at e13, but subsequently decreased with age in an anterior to posterior fashion (data not shown).
Figure 5A shows whole-mount in situ hybridizations of the foregut region encompassing the stomach and pancreas at e15 and e18 of development. The age-matched sense controls are placed above and the anti-sense hybridized tissue below. Note that the stomach and the attached spleen are devoid of signal at all stages examined. Expression in the e15 pancreas is most intense along the distal edge of the developing pancreas, where acinus formation begins. As the pancreas develops, there is increased expression and signal strength of pask along the more distal aspects. The proximal portion of the pancreas, in which the larger ducts reside closest to the duodenal origin of the pancreas, is negative for pask expression. The signal for pancreatic expression of pask1 increased significantly by e18, accompanied by the explosion in formation of exocrine epithelium.
As early as e15, exocrine structures could be seen in the developing pancreas. Figure 5B shows a 5-μm paraffin section of an e15 foregut hybridized with pask. The stomach was devoid of pstk1 signal but the nascent pancreatic acini had strong expression. Pask was not seen in the mesenchyme. Note that the duct bed that gives rise to the branching acini (arrows) has little pask signal. The inset shows a higher-magnification photo detailing the strong acinar expression. Figure 5C is a 5-μm paraffin section of an e17 pancreas hybridized for pask (purple) and immunohistochemically stained for insulin (brown). The exocrine acini were again strongly positive for pask hybridization. The duct bed had very low levels of pask expression and was giving rise to strongly insulin-immunoreactive cells. The expression of PSTK1 mRNA and insulin protein appeared mutually exclusive. Further in situ studies in the adult pancreas showed continued extensive exocrine acinar expression (data not shown).
Discussion
Our screen for kinases expressed in the developing pancreas uncovered a number of molecules whose expression levels varied with stage of development. Of particular interest are the PDGF-α and Flk-1 receptors, which bind PDGF and VEGF (Terman et al. 1992), respectively. The levels of expression of these receptors, as determined by the number of clones detected, appear upregulated between e12 and e14 in rat. Expression of Flk-1 was previously mapped to fetal islets (Oberg et al. 1994). Our results indicate a role for Flk-1 much earlier in pancreatic development, before the formation of islet tissue. Another observation of interest is the upregulation of c-Abl between e14 and e15. c-Abl is associated with a number of key cellular processes, including apoptosis and cell-cycle regulation (Wong et al. 1995; Theis and Roemer 1998). Expression of many of the detected kinases and receptors has been localized to discrete cell types using immunohisto-chemistry and will be described elsewhere.

Alignment of pask/pstk-1 related proteins demonstrates homology to the Ste-20-like family of kinases. Shown are the rat pask and human pstk protein sequences as well as the discovered human homologue PSTK2 described here and aligned with the related human protein Ste20-like kinase (Genbank AAD42039) and MST1 (Genbank AAA83254). This alignment was produced with clustalw (Thompson et al. 1998) using the default parameters. Amino acids identical in all five proteins are shaded with black background and positions at which the majority of amino acids are identical or chemically similar are shaded in gray. The kinase domain, as defined by Pfam (Bateman et al., 2000; http://pfam.wustl.edu), extends from amino acid 72 to 346. Within this domain, these five proteins are 34% identical, with extensive conserved substitutions
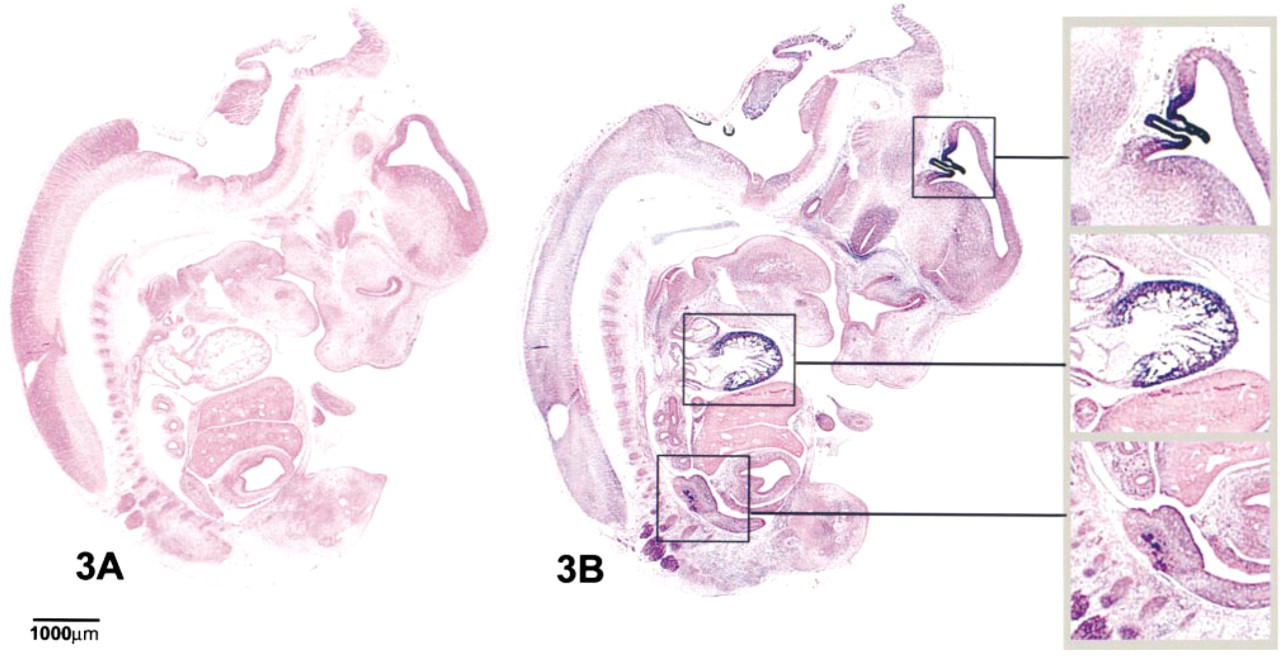
Expression of pstk1 mRNA in a parasagittal section of an e14 rat embryo. Digoxigenin staining is shown in blue and the embryo is counterstained with eosin. (
During the course of our mapping studies, expression of pask was discovered and studied. The expression pattern of this gene suggests that it may be important in the early formation of a variety of tissues, including the pancreas. The amino acid sequence of pask indicates that it is most closely related to the OPK Group III family of protein kinases as defined by Hanks and Quinn (1991), which also includes the PAK kinase and the yeast Ste20 kinase. This similarity appears to be restricted to the catalytic domain of the protein. On identification of this rat sequence, we searched the human EST database to determine whether sequences from human homologues were present. A total of 60 EST sequences were detected. These sequences appear to be encoded by two separate genes, which we refer to here as human pstk1 and human pstk2. Human pstk1 encodes a protein of at least 520 amino acids. However, the available EST sequences do not appear to contain the initiation codon. The available 520-amino-acid polypeptide is 97.6% identical to the rat pask protein. During the course of our database searches, we discovered a second human kinase. This second human protein appears to be encoded by a related but distinct mRNA sequence derived from seven EST's. This nucleotide sequence is 1778 nt in length and is referred to as human pstk2. It does not appear to contain the initiator start codon and does not terminate in polyA. The nucleotide sequence encodes a 522-amino-acid protein which is 77.6% identical to the rat pstk protein and 78.1% identical to the human pstk1 protein.
Rat pask transcripts were detectable in the embryo by in situ hybridization as early as e9 (data not shown). Expression was strong in the heart, telencephalic neuroepithelium, and the first two aortic arches, and was weak in the region of the developing gut endoderm. This pattern of expression was observed at all stages studied, with spreading of signal observed in tissues and organs such as the pancreas, adrenal medulla, kidney, dorsal root ganglia, heart, and choroid plexus. CNS expression appeared primarily in the cochlear ganglion at e15, and was not detectable in any other neural structures. Expression of pask increased with time in most positive tissues studied, an exception being the early gut epithelium in which signal progressively decreased in an anterior to posterior fashion, thus indicating that pask is a marker of early gut epithelium but is subsequently lost with differentiation.
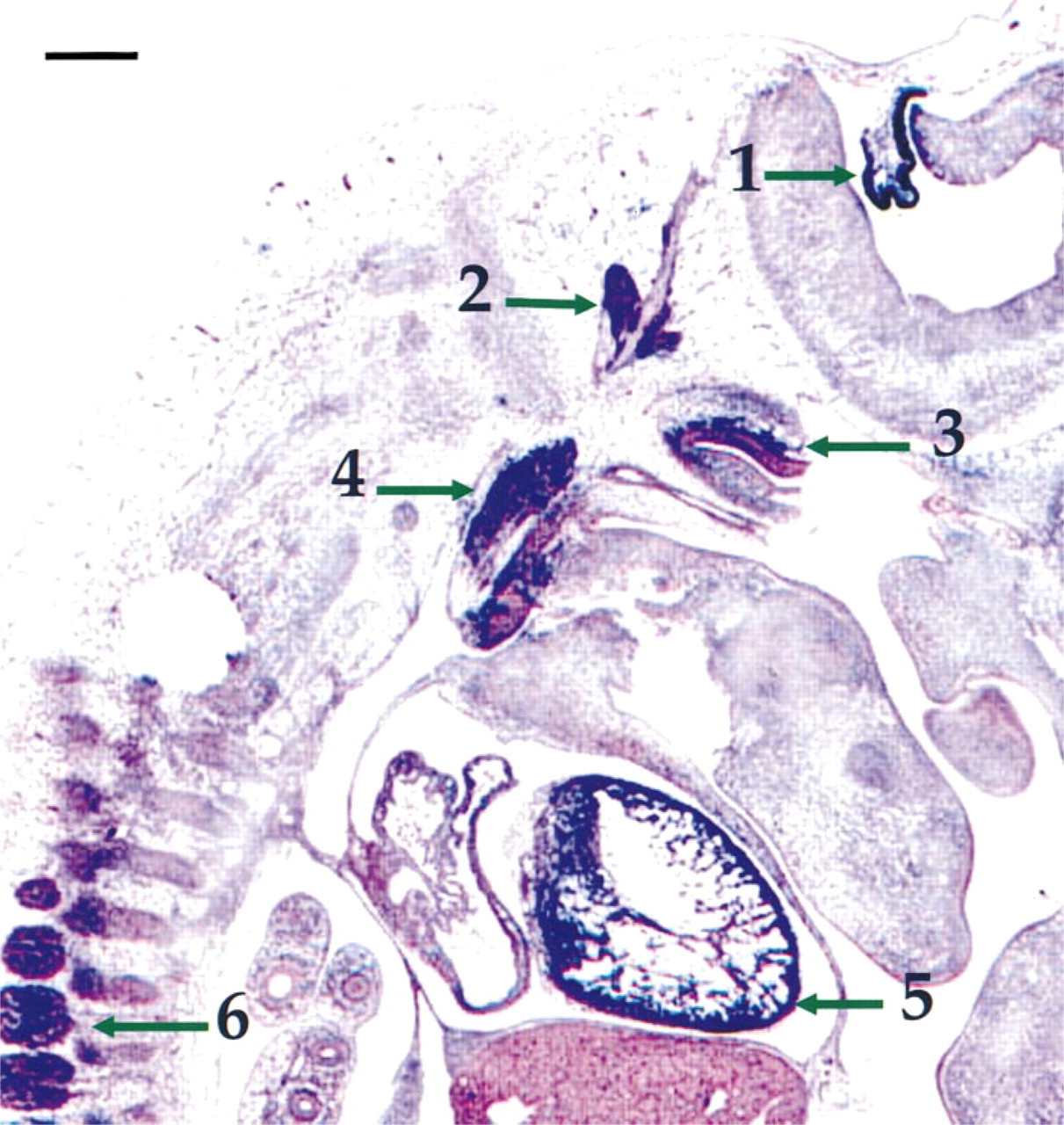
Enlarged section of an e15 rat parasagittal section shows extensive pask expression in the heart and nervous system. Arrow 1 shows the continued expression in the choroid plexus. Arrows 2, 3, and 4 indicate Froriep's ganglion, the vestibulocochlear ganglion, and glossopharyngeal ganglion, respectively, which are positive for pstk1 expression. Arrow 5 shows the extended expression of pstk1 in the myocardium, and arrow 6 shows that the anterior dorsal root ganglia also express pask. Endodermal organs such as the lung and liver, are negative for expression. Bar = 250 μm.
In contrast, the pancreas exhibits early expression in the entire bud epithelium, which then progressively localizes to the exocrine acini. In the e12 rat pancreas, expression of pask is found in the early epithelial bud, but by e14/15 expression is predominantly found in the distal portions of the pancreas, where the bulk of the acini are budding off the main epithelial trunk. By e17, there is clear exclusion of pask expression from the main branch of epithelium that gives rise to the insulin-positive β-cells and clear localization to the branching exocrine acini. Studies of the adult pancreas show continued high expression of pask in the exocrine tissue (data not shown).
The role of PASK in vertebrate development is not yet known. However, our spatial expression studies in the embryo might provide insight as to its role in early tissue formation. The expression of pask in early organogenesis of a number of tissues indicates a possible early role. It is interesting to speculate that many of the tissues that express pstk1 also express, and in some cases are known to be regulated by, members of the bone morphogenetic protein (BMP) gene family, a branch of the TGF-β superfamily, whose signal is mediated by activating serine/threonine receptor kinases (Takeda et al. 1994; Rosenzweig et al. 1995). For example, the BMPs 4, 5, 6, and 7 are expressed in the choroid plexus (Furuta et al. 1997), BMP 7 has been mapped to the developing kidney epithelium, heart, and intestinal epithelium (Helder et al. 1995), BMP 3 (Matsui 1998) and BMP-3B (Hino et al., 1996) are expressed in the testis, and BMP 4 is expressed in a subregion of the developing heart (Eisenberg and Markwald 1995). Interestingly, mapping of the CNS and PNS expression of BMP receptors shows strong CNS and dorsal root ganglia expression (Zhang et al. 1998). In addition, BMPs have been shown to regulate the survival of a sympathoadrenal cell line (Song et al. 1998) and to regulate renal branching morphogenesis (Piscione et al. 1997). The expression of pstk1 in early kidney, heart, choroid plexus, and dorsal root ganglia, correlates well with the BMP expression studies. Interestingly, the role of BMPs in pancreatic development has not been extensively investigated, although recent reports map BMP expression to the adult pancreas (Hino et al. 1996; Paralkar et al. 1998).
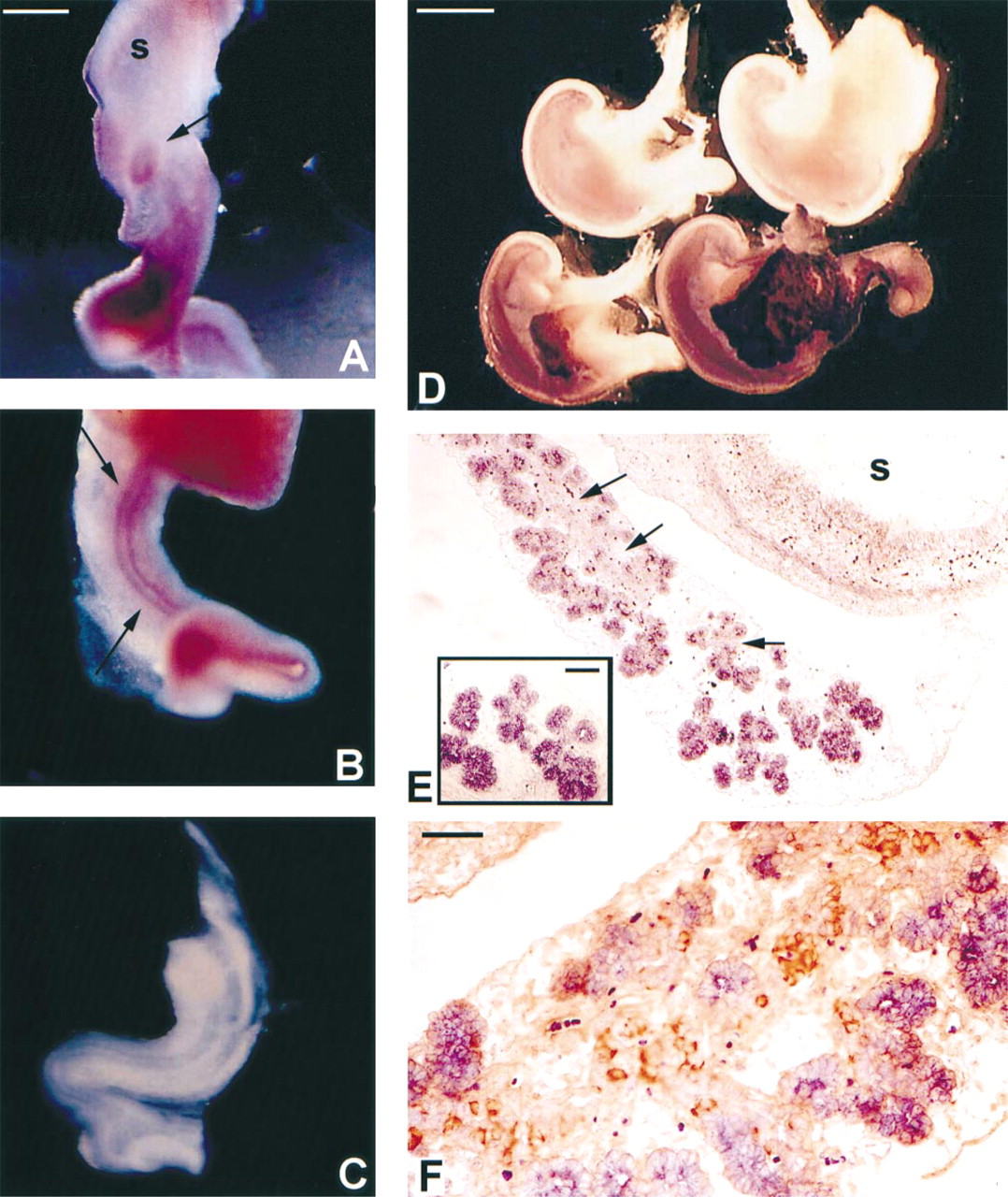
Expression of pask in developing pancreas. (
The recent work of Ushiro et al. (1998) describes cloning of the identical molecule which they term PASK, for the proline- and alanine-rich 5′ region. In their work they also describe the 2-kb and 4-kb transcripts but, interestingly, only the smaller transcript appears to be translated in vitro. Spatial expression analysis performed on adult tissue showed overlap in some tissues in which we detected early embryonic transcripts, such as the choroid plexus, kidney, heart, and adrenal medulla. Therefore, pask/pstk1, for which our early embryonic studies indicate that its expression is correlated with early tissue formation, continues to be expressed in these tissues and might therefore play a role in maintenance of adult tissue function. Interestingly, there were a significant number of specialized tissues, such as spleen, olfactory bulb, and stomach, that did not show significant expression in our analysis but were later shown to express this gene in the adult (Ushiro et al. 1998). This might indicate multiple roles of this kinase in either early tissue formation and/or maintenance of differentiated function in the adult state. Ushiro et al. (1998) suggest a role for this molecule in secretory epithelium function, which is consistent with our findings of strong expression in pancreatic epithelia and exocrine tissue, although they did not study pancreatic expression.
The approach of using degenerate PCR to detect the expression of specific gene families as a function of time in development, followed by spatial localization either by in situ hybridization or immunohistochemistry, is useful in studying the ontogeny of that tissue. For example, the expression of many of the kinases and receptors detected during this study had not been previously mapped to the pancreas. The expression of key regulatory molecules can be rapidly detected and mapped to particular cell types at key physiological transitions during development in tissue targets of interest, e.g., the gain of exocrine or endocrine function in the case of the pancreas through the use of these methods. This mapping also provides markers by which specific cell types can be identified and perhaps isolated and, in the process, information regarding possible regulation of either growth or differentiation can be obtained. In addition, as demonstrated here, this approach also provides the possibility of discovering new and interesting gene family members.
In conclusion, the approach that we have taken in terms of mapping the temporal and spatial expression of specific gene family members as a function of tissue formation and development is a useful one. This process combines precise dissection of stage-specific tissue with degenerate PCR, sequencing, and high-throughput in situ hybridization analysis. This approach can be applied to any target tissue or organ of choice. We are now exploring subtractive array hybridization technologies to increase the throughput of discovering genes within given families that are differentially expressed either as a function of time in development and/or spatial expression within a given tissue. The knowledge gained through these efforts will help to develop novel markers by which specific cell types can be identified in subsequent lineage studies, map the expression of important signal transduction genes as demonstrated herein and, finally, provide insights into mechanisms of growth and differentiation.
Footnotes
Acknowledgements
We would like to thank David Kagan for help with molecular biology, David Bumcrot for help with in situ hybridization, Nagesh Mahanthappa and Irina Karavanova for help with in situ analysis, and Mina Peshavaria for critical analysis of the manuscript.