
Abstract
Tenascin-C (TN-C) and its isoforms are multidomain extracellular matrix (ECM) proteins that are believed to be involved in the regulation of stromal–epithelial interactions. Some of the interactions between TN-C and cells are mediated by integrins. In this study we analyzed the expression of TN-C and its large molecular weight splice isoform (TN-CL) and the putative TN-C-binding α9 and αvβ6 integrins during human wound repair. In 3-day-old oral mucosal wounds, immunoreactivity for α9 integrin localized abundantly at the migrating basal wound epithelial cells. TN-C and TN-CL were localized in the matrix between and underneath α9-expressing epithelial cells. In parallel with gradual downregulation of α9 integrin immunoreactivity in 7-day and older wounds, the expression of αvβ6 integrin was temporarily induced. Integrin αvβ6 co-localized in the same area as TN-C and TN-CL immunoreactivity at the cell–cell contacts of the basal and suprabasal cell layers of the wound epithelium. During granulation tissue formation and reorganization from 7 to 28 days after wounding, TN-C and TN-CL were abundantly localized in the granulation tissue. The findings show that TN-CL is expressed under the migrating epithelial front and in the granulation tissue during matrix deposition in wound repair. Preferential localization of α9 integrin in migrating epithelial cells and of αvβ6 integrin in epithelium after wound closure suggests different functions for these integrins in wound repair.
Keywords
W
Tenascin (TN-C) is a large hexameric glycoprotein that is abundantly expressed, particularly at epithelial–mesenchymal interaction sites during embryogenesis. TN-C shows a more restricted distribution in various adult tissues, including skin and oral mucosa, in which it localizes mostly to the papillary connective tissue immediately beneath the basement membrane zone (Crossin 1996; Lukinmaa et al. 1996). However, abundant expression of TN-C emerges in adult tissue during conditions that are associated with high rates of cell turnover and migration, including wound repair, blistering or hyperproliferative diseases; and tumorigenesis (Kruse et al. 1985; Chiquet-Ehrismann et al. 1986, 1995; Mackie et al. 1987; Borsi et al. 1992; Aukhil et al. 1996; Crossin 1996; Haapasalmi et al. 1996; Latijnhouwers et al. 1996).
Human TN-C is composed of structurally different domains, including epidermal growth factor-like repeats, units similar to fibronectin Type III homology repeat (TNfn), and a C-terminal sequence with homology to β- and γ-chains of fibrinogen (Siri et al. 1991). Up to nine domains located between TNfn5 and TNfn6 can be alternatively spliced in a developmentally regulated and cell-specific manner to generate structural variations among different TN-C molecules (Matsuoka et al. 1990; Prieto et al. 1990; Siri et al. 1991; Rettig et al. 1994; Mighell et al. 1997). Two major TN-C isoforms that are generated by including or omitting the type three repeats in the mRNA have different functional properties (Siri et al. 1991), and their expression is differentially regulated by cytokines in vitro (Tucker et al. 1993; Rettig et al. 1994; Zhao and Young 1995). The high molecular weight isoform is preferentially expressed at the onset of cell migration, proliferation, or tissue remodeling during neoplasia (Borsi et al. 1992), development (Matsuoka et al. 1990; Prieto et al. 1990), and blistering disease in skin (Schenk et al. 1995). Previous reports using antibodies that recognized all forms of TN-C have shown that, during early wound repair, TN-C expression is first upregulated at the connective tissue at wound margins and later in the granulation tissue (Mackie et al. 1988; Chuong and Chen 1991; Whitby et al. 1991; Luomanen and Virtanen 1993; Haapasalmi et al. 1996; Latijnhouwers et al. 1996, 1997). In addition, epithelial cells in the migrating front of the wound epithelium express TN-C mRNA (Aukhil et al. 1996). Biochemical analysis of TN-C from mouse skin wounds suggests that the amount of the higher molecular weight form is increased in wound granulation tissue compared with normal connective tissue (Chuong and Chen 1991).
Cell adhesion to TN-C is mediated by integrins in a cell type-specific manner (Chiquet–Ehrismann et al. 1995; Crossin 1996). Integrins are transmembrane heterodimeric molecules that are composed of noncovalently associated α- and β-subunits. To date, 18 α- and eight β-subunits have been described, which can form at least 24 different αβ heterodimers (Brake-busch et al. 1997; Katz and Yamada 1997; Camper et al. 1998). Among integrins present in epithelial cells, αvβ6 and α9β1 recognize the third TNfnIII (TNfn3). In TNfn3, however, αvβ6 integrin recognizes the common integrin recognition motif Arg-Gly-Asp (RGD), whereas α9β1 integrin binds to a sequence that includes the Ile-Asp-Gly (IDG) motif (Prieto et al. 1992; Yokosaki et al. 1994, 1998).
Previous studies have shown changes in the expression and distribution of epithelial integrins in wound repair compared with normal tissue (Juhasz et al. 1993; Larjava et al. 1993; Haapasalmi et al. 1996). Of the potentially TN-C-binding integrins, αvβ6 is not normally expressed in the epithelium but its expression is induced during wound repair (Breuss et al. 1995; Haapasalmi et al. 1996). Integrin α9-subunit is a recently characterized member of the β1 integrin family that is expressed by specialized cells, including stratified squamous epithelium in skin (Palmer et al. 1993; Yokosaki et al. 1994; Stepp et al. 1995; Wang et al. 1995; Stepp and Zhu 1997). The expression of integrin α9-subunit is developmentally regulated and coincides with the development of stratification of epithelium (Stepp et al. 1995; Wang et al. 1995), and its expression is also associated with cell proliferation in vivo and in vitro (Stepp and Zhu 1997; Desloges et al. 1998).
In this study we tested our hypothesis that the expression of α9β1 and αvβ6 is coordinately upregulated with the large TN-C isoform during wound repair. We provide evidence that α9 but not β6 integrin is expressed by migrating epithelial cells during early wound repair. This coincides with deposition of TN-C, including the large isoform, under the migrating epithelial cells. After epithelial confrontation, immunoreactivity for α9 integrin is reduced, whereas expression of αvβ6 integrin is induced and it localizes in the same area as TN-C. During later tissue differentiation and maturation, immunoreactivity for αvβ6 integrin and TN-C at the epithelial cell interfaces is coordinately reduced, whereas the immunoreactivity for TN-C is still strong in granulation tissue compared with normal connective tissue.
Materials and Methods
Histological Specimens
Tissue biopsy specimens of normal keratinized masticatory mucosa (gingiva, n = 10; alveolar mucosa, n = 3; palatal mucosa, n = 3) and non-keratinized buccal mucosa (n = 3) and ventral tongue (n = 1), were obtained from four healthy volunteers. Three-day, 7-, 14-, and 28-day-old wounds from keratinized palatal gingiva were obtained from the collection of wound sections from three volunteers previously used to localize integrins and basement membrane components during mucosal wound healing (Larjava et al. 1993). The tissue samples were obtained from the University of British Columbia, Vancouver, Canada, and University of Turku, Finland. The procedures were approved by the Ethical Committees for Human Experimentation of the corresponding organizations.
Antibodies
To localize different alternatively spliced TN-C isoforms, two monoclonal antibodies (MAbs) recognizing different TN-C epitopes were used (gifts from Dr. Luciano Zardi, Instituto Nazionale per la Ricerca sul Cancro, Genoa, Italy). MAb BC-4 recognizes an epitope within the EGF-like sequences that is common to all forms of TN-C. MAb BC-2 reacts with epitopes in the alternatively spliced FN-like repeats A1 and A4. In immunoblotting, MAb BC-2 recognizes the large TN-C isoform (280 kD) and MAb BC-4 the small and the large isoforms (190 kD and 280 kD, respectively). The characterization of the antibodies has been described previously (Siri et al. 1991; Balza et al. 1993). To localize integrin subunits, an affinity-purified anti-peptide antibody against human α9 integrin subunit (Palmer et al. 1993) and an MAb against αvβ6 integrin (E7P6; Weinacker et al. 1994) were used (generous gifts from Dr. Dean Sheppard, University of California, San Francisco).
Immunohistochemical Staining
Immediately after biopsy, fresh tissue blocks were mounted in Histoprep (Fisher Scientific; Fair Lawn, NJ) and snap-frozen in liquid nitrogen. Frozen sections (6 μm) were cut and fixed with −20C acetone for 5 min and stored at −70C until used. For immunoperoxidase staining for integrin α9-subunit, sections were incubated in 0.3% hydrogen peroxide in methanol for 30 min and washed in PBS/BSA. Sections were rinsed and incubated with normal blocking serum (Vectastain; Vector Laboratories, Burlingame, CA) for 60 min at room temperature (RT) and then incubated with the primary antibody in PBS containing 0.1% bovine serum albumin (BSA; Sigma Chemical, St Louis, MO) in a humid chamber at 4C for 16 hr. After washing with PBS/BSA, sections were incubated with biotinylated anti-rabbit antibody for 60 min and then reacted with ABC avidin–peroxidase reagent (Vectastain Elite kit; Vector Laboratories). Chromogen was developed using diaminobenzidine (DAB; Chemicon, Temecula, CA) and nickel–silver enhancement (Przepiorka and Myerson 1986). Reactions were monitored until suitable color development was achieved and sections were then washed with distilled water. The sections were mounted with Permount (Fisher Scientific; Pittsburgh, PA).
For immunofluorescence staining of αvβ6 integrin, sections were incubated for 60 min at RT with PBS/BSA and then with the primary antibody as above. After washing, sections were incubated with anti-mouse Ig–rhodamine (Boehringer Mannheim) in PBS/BSA for 60 min at RT. Sections were washed, briefly air-dried, and mounted with cyanoacrylate glue. Control sections were incubated with purified nonimmune rabbit or mouse IgG (10 μg/ml; Sigma) in PBS/BSA instead of the primary antibody (not shown).
Immunohistochemical staining for TN-C isoforms was performed using the alkaline phosphatase–monoclonal anti-alkaline phosphatase (APAAP) method. The primary antibodies BC-2 and BC-4 (undiluted hybridoma supernatant) were incubated for 30 min at RT. After washing with Tris buffer, sections were treated with rabbit anti-mouse immunoglobulin (diluted 1:70; Dako, Glostrup, Denmark) and then with the mouse APAAP complex (Dako). Both incubations were carried out for 30 min at RT. To increase the staining intensity, the incubation with the rabbit anti-mouse immunoglobulin and with the APAAP complex was repeated twice. Naphthol-AS-biphosphate (Sigma) and new fuchsin (Merck; Darmstadt, Germany) were used as substrate and developer, respectively. To inhibit endogenous tissue enzyme activity, the developing solution was supplemented with 0.25 mmol/liter levamisole (Sigma). After color development, the sections were counterstained briefly with hematoxylin and mounted in glycerol gelatin (Serva; Heidelburg, Germany). As negative control, the primary antibody was replaced by nonimmune serum (not shown).
Up to 10 sections of each sample were stained with each antibody. The staining was examined with a Zeiss Axioskop 20 light and fluorescence microscope and was photographed with an MC 80 Zeiss microscope camera. To evaluate the overall histology, including collagen fiber organization and degree of inflammation in the wound sections, a set of samples was stained with Mallory's phosphotungstic acidhematoxylin method (Puchtler et al. 1963) or with hematoxylin and eosin (not shown).
Results
Localization of α9 and αvβ6 Integrin and TN-C Splice Isoforms in Normal Keratinized and Non-keratinized Oral Mucosa
The expression of α9 integrin in keratinized epithelium of gingiva and alveolar and palatal mucosa was relatively weak. At epithelial rete ridges, α9 integrin was expressed by the basal epithelial cells and the immediately suprabasal cells, and was localized most abundantly against the basement membrane (Figure 1A). In the connective tissue, α9 integrin localized at blood vessels (Figure 1A). Immmunoreactivity for antibody BC-2, recognizing the large TN-C splice isoform, and for BC-4, recognizing all forms of TN-C, localized similarly at the epithelial basement membrane zone and over collagen fibers throughout the subepithelial connective tissue (Figure 1D and 1E). The strongest staining of TN-C was localized at the ECM of the connective tissue papilla area. In the deep connective tissue matrix, the staining intensity for TN-C decreased (Table 1). Some of the subepithelial vascular basement membrane areas were stained with anti-TN-C antibodies (Figure 1D and 1E).
In non-keratinized buccal mucosa (Figure 1B) and ventral tongue (Figure 1C), immunoreactivity for α9 integrin subunit was stronger compared with normal keratinized gingiva and showed most abundant staining at the cell membrane of the basal cells. In addition, α9 integrin had a pericellular distribution at the basal and suprabasal cell layers. In the connective tissue, α9 integrin localized at blood vessels (Figure 1B and 1C). Strongest immunoreactivity for both BC-2 (Figure 1E and 1F) and BC-4 (Figure 1H and 1I) antibodies was localized at the epithelial and most of the vascular basement membranes. Staining intensity of the blood vessels with the BC-4 antibody was slightly more intense than with the BC-2 antibody. At connective tissue papilla areas and directly under the basement membrane, TN-C was localized on collagen fibers but, unlike in keratinized mucosa, no TN-C immunoreactivity was extended into the deeper areas of the subepithelial connective tissue (Figures 1E, 1F, 1H, and 1I; Table 1). No expression of αvβ6 integrin was seen in keratinized and nonkeratinized normal oral mucosa (not shown).
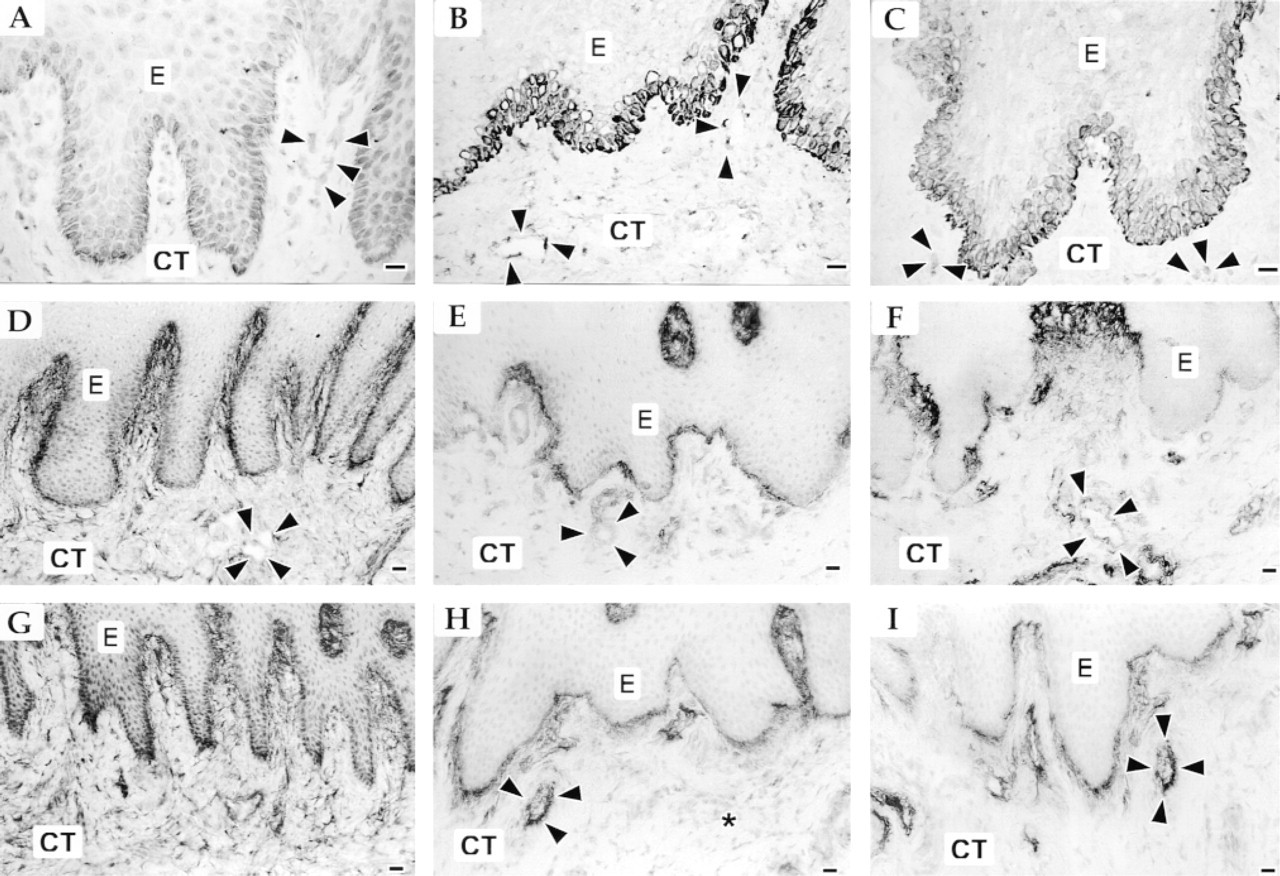
Immunolocalization of α9 integrin (
Localization of α9 and αvβ6 Integrins and of TN-C Splice Isoforms During Wound Healing of Keratinized Oral Mucosa
In 3-day-old wounds, epithelium had started to migrate through the wound bed provisional matrix. A moderate inflammatory cell infiltrate was localized in the wound bed and in the connective tissue under the epithelium next to the wound. In 7-day-old wounds, epithelium had completely covered the wound space and had started to differentiate. The inflammation in the granulation tissue was reduced compared with earlier time points, and collagen deposition in the granulation tissue had started. At 14 days after wounding, only scattered inflammatory cells were seen in the granulation tissue. New collagen fibers in the granulation tissue were relatively thin compared with normal tissue, and were mostly arranged horizontally across the wound area. By Day 28, the inflammation had completely disappeared, but the granulation tissue could still be distinguished from the more organized normal connective tissue and was composed of thick collagen fibers that were mostly organized horizontally across the granulation tissue. At this time, epithelium had completely regenerated and was morphologically similar to adjacent unwounded epithelium.
Localization and relative staining intensity of α9 and αvβ6 integrins and tenascin-C with an antibody that recognizes all tenascin-C isoforms (BC-4) or the large molecular weight isoform only (BC-2) in normal, nonkeratinized, and keratinized oral mucosa and in 3-, 7-, 14-, and 28-day-old woundsa

aRelative intensity of immunostaining and inflammation:
In the epithelium immediately next to the wound margin, and in keratinocytes migrating through the provisional wound matrix of the 3-day-old wounds, immunoreactivity for α9 integrin was strong in the basal cells (Figure 2A and 2B; Table 1). The most intense staining was localized at the basal cell membranes of the keratinocytes at the leading edge of the migrating epithelial front (Figure 2A and 2B, large black arrowheads). In the wound fibrin clot, α9 immunoreactivity localized to scattered inflammatory cells (Figure 2A and 2B, small arrows). In the unwounded connective tissue, immunoreactivity for α9 integrin localized abundantly at blood vessels (Figure 2A, open arrowheads). In 7-day-old wounds, in tissue sections from the central wound where migrating epithelial fronts had just joined to cover the wound surface, expression of α9 integrin at the basal membranes of the basal cells was still strong (Figure 2C and 2D, large black arrowheads). However, in serial sections further away from the central wound where epithelium had already started differentiation, immunoreactivity for α9 integrin was weaker (Figure 2E and 2F). Some forming blood vessels in the granulation tissue also showed positive staining for α9 integrin (Figure 2C and 2E, open arrowheads). As maturation of the epithelium continued for 14 days after wounding, epithelium did not show any immunoreactivity for α9 integrin (Table 1). In contrast, blood vessels in the granulation tissue showed strong staining for α9 integrin (Table 1). After 28 days, expression of α9 integrin was normalized in the epithelium. Most abundant staining localized against the basement membrane, similar to normal unwounded tissue (Figure 2G and 2H, large black arrowheads). Blood vessels at the subepithelial granulation tissue also showed positive staining for α9 integrin (Figure 2G and 2H, open arrowheads).
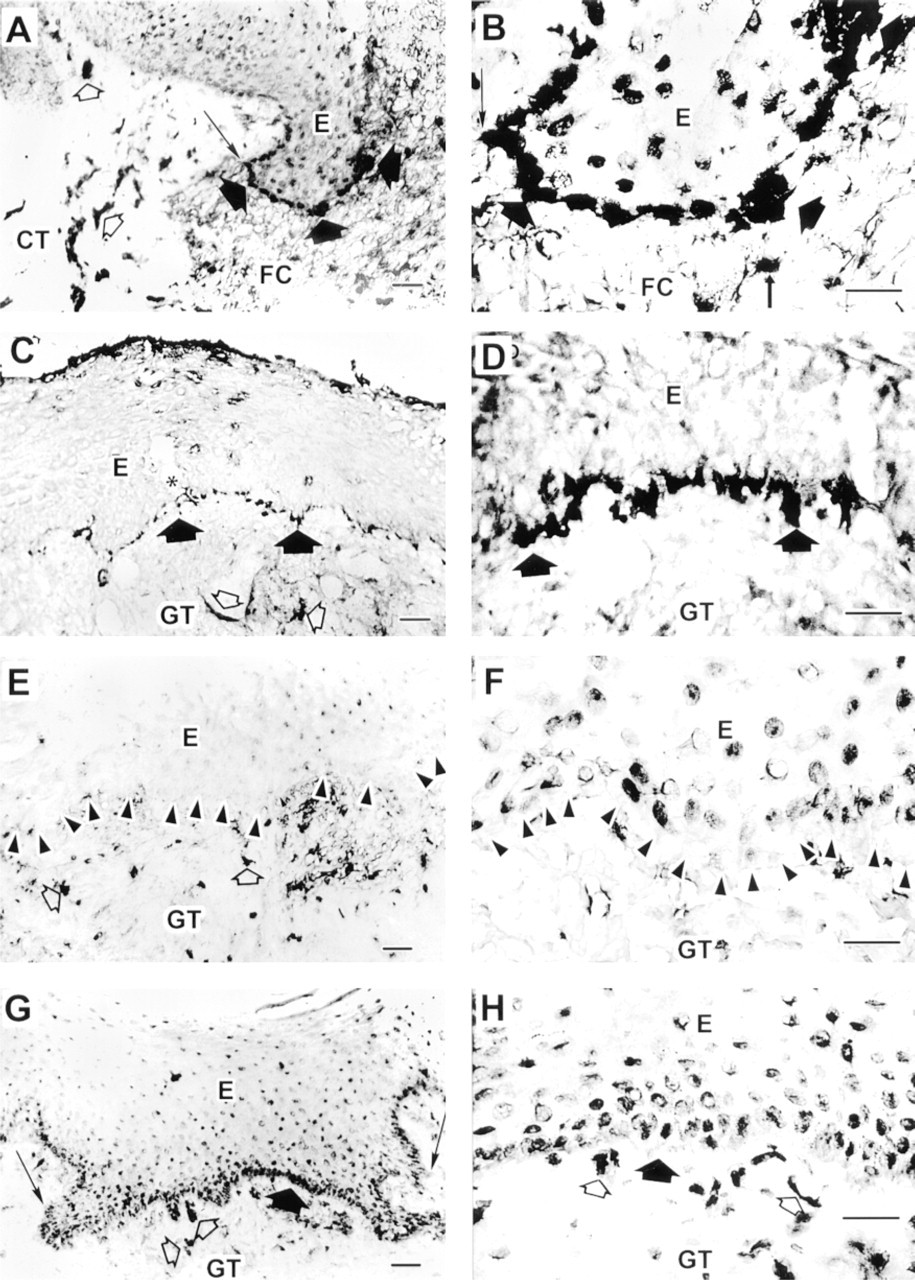
Localization of α9 integrin in 3- (
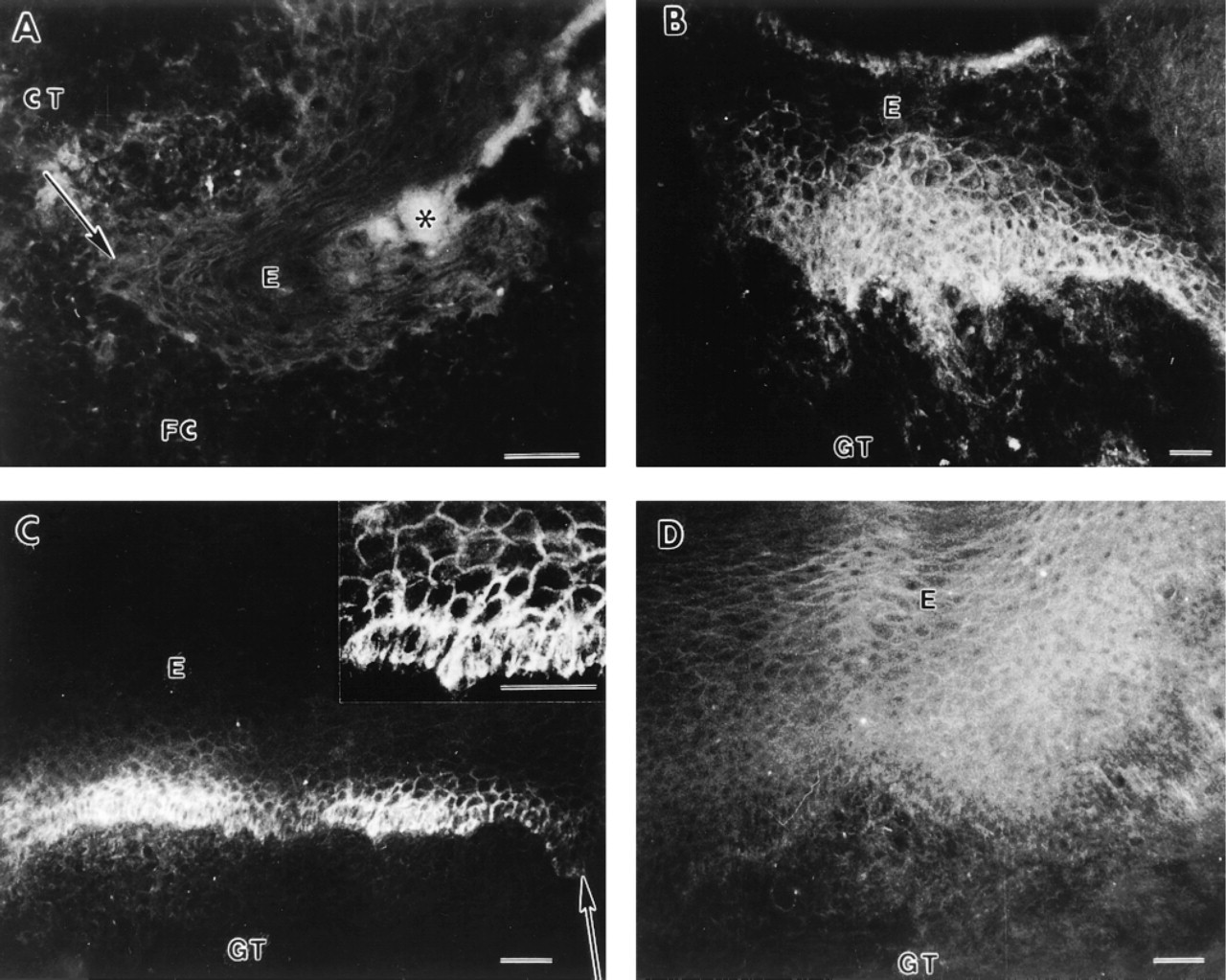
Localization of αvβ6 integrin in 3- (
No immunoreactivity for αvβ6 integrin was detected in the 3-day-old wounds (Figure 3A; Table 1). After 7 days, expression of αvβ6 integrin was induced at the cell membranes of the basal and several suprabasal cell layers of the wound epithelium (Figure 3B). At Day 14 after wounding, expression of αvβ6 integrin at the basal and immediately suprabasal cell layers was still strong (Figure 3C, inset), although its expression in the most suprabasal cell layers was reduced compared with the 7-day-old wounds (Figure 3C). After 28 days, the epithelium was totally negative for immunoreactivity for αvβ6 integrin (Figure 3D; Table 1).
After 3 days, immunoreactivity for both BC-2 (Figure 4A and 4B) and BC-4 (Figure 5A and 5B) anti-TN-C antibodies was similar and was localized around the cell membranes of the migrating basal epithelial cells, but no TN-C staining was seen in the provisional matrix of the wound bed (Figures 4A, 4B, 5A, and 5B; Table 1). At the cell–cell interfaces of the basal and some suprabasal cell layers in the epithelium next to the wound, immunoreactivity for both BC-2 (Figure 4A and 4B) and BC-4 (Figure 5A and 5B) antibodies also localized between the basal and immediately suprabasal epithelial cells. In the nonwounded area further away from the wound, TN-C immunoreactivity between the epithelial cells was gradually suppressed (Figure 4A and 5A). After 7 days, immunoreactivity for BC-2 (Figure 4C and 4D) and BC-4 (Figure 5C and 5D) antibodies was similar and was localized at the cell–cell interfaces at the basal and suprabasal cell layers of the wound epithelium. Furthermore, immunoreactivity for both BC-2 (Figure 4C) and BC-4 (Figure 5C) antibodies was relatively strong throughout the wound granulation tissue compared with adjacent normal connective tissue (Table 1). After 14 days of wounding, immunoreactivity for both BC-2 (Figure 4E and 4F) and BC-4 (Figure 5E and 5F) antibodies was similarly reduced between the suprabasal epithelial cell layers and was localized only around cell membranes of the basal cells. The granulation tissue showed similar strong immunoreactivity for both BC-2 (Figure 4E and 4F) and BC-4 (Figure 5E and 5F) antibodies. After 28 days, immunoreactivity for both BC-2 (Figure 4G and 4H) and BC-4 (Figure 5G and 5H) antibodies was no longer localized at the cell–cell interfaces at the wound epithelium, although their staining intensity was still relatively strong throughout the wound granulation tissue (Table 1).
Discussion
By using different human and animal wound healing models, previous studies have documented increased TN-C deposition and expression during wound repair (Mackie et al. 1988; Chuong and Chen 1991; Whitby et al. 1991; Juhasz et al. 1993; Luomanen and Virtanen 1993; Aukhil et al. 1996; Latijnhouwers et al. 1996, 1997). Furthermore, biochemical analysis of TN-C from mouse skin wounds has indicated that the amount of the higher molecular weight form is increased during wound repair compared with normal tissue (Chuong and Chen 1991). In rat dermal wounds, Mackie et al. (1988) localized TN-C under the migrating wound epithelium. However, by comparing the expression of the TN-C splice isoform mRNA containing or lacking the alternatively spliced sequences, Aukhil and co-workers (1996) did not detect any differences in the relative ratios of TN-C isoform mRNAs between wounded and unwounded control tissue in rat. On the other hand, by using various anti-TN-C antibodies previous studies from our laboratory and others have not detected TN-C deposition under the migratory epithelial cells in the early adult human wounds (Haapasalmi et al. 1996; Latijnhouwers et al. 1997). In the present study, by using MAbs against TN-C isoforms, we were able to show that TN-C is specifically deposited under migrating epithelial cells during epithelial migration in early wounds. The fact that we detected a similar staining pattern with both the BC-2 antibody, recognizing the large molecular weight TN-C splice isoform, and with the BC-4 antibody, recognizing all TN-C isoforms, suggests that the large isoform is indeed one of the molecules whose expression is already induced at the migratory epithelial front. However, we cannot exclude the possibility that the large TN-C isoform is co-expressed with the small TN-C isoform. One of the important properties of TN-C is its ability to modulate cell adhesion. Specifically, the large TN-C isoform interferes with or supports cell adhesion and migration on fibronectin, depending on the composition of the alternatively spliced domains in the molecule. The smaller TN-C isoform also inhibits cell adhesion but binds more strongly to FN compared with the large isoform (Chiquet–Ehrismann et al. 1991; Kaplony et al. 1991; Murphy–Ullrich et al. 1991; Fisher et al. 1997a, b; Phillips et al. 1998). It is possible that the relative abundance and molecular composition of the TN-C isoforms present in wounds may have modulatory effects on the epithelial cell migration on the provisional wound matrix that contains FN. However, functional studies will be needed to elucidate this point.
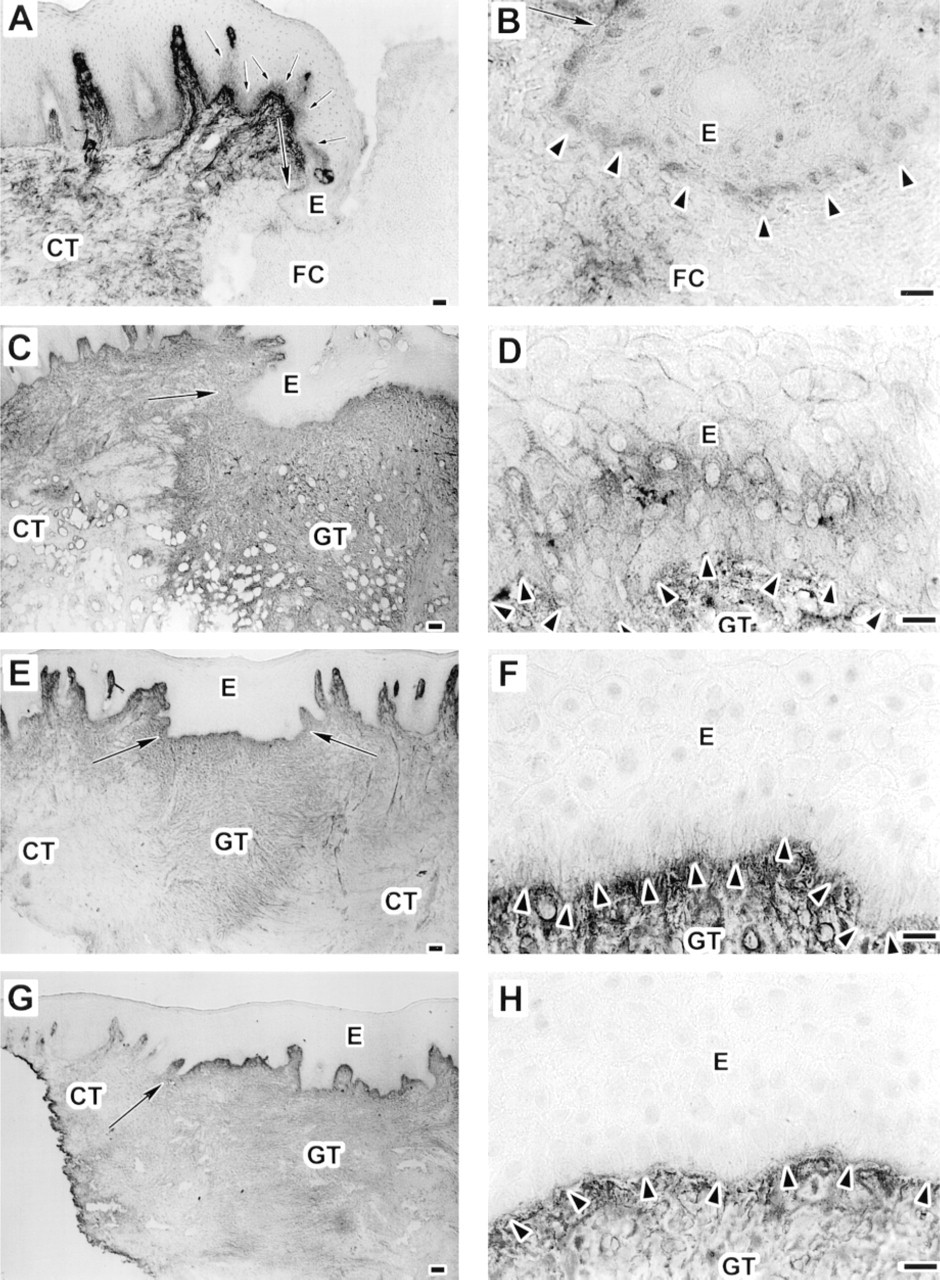
Immunolocalization of large tenascin-C isoform using MAb BC-2 in 3- (
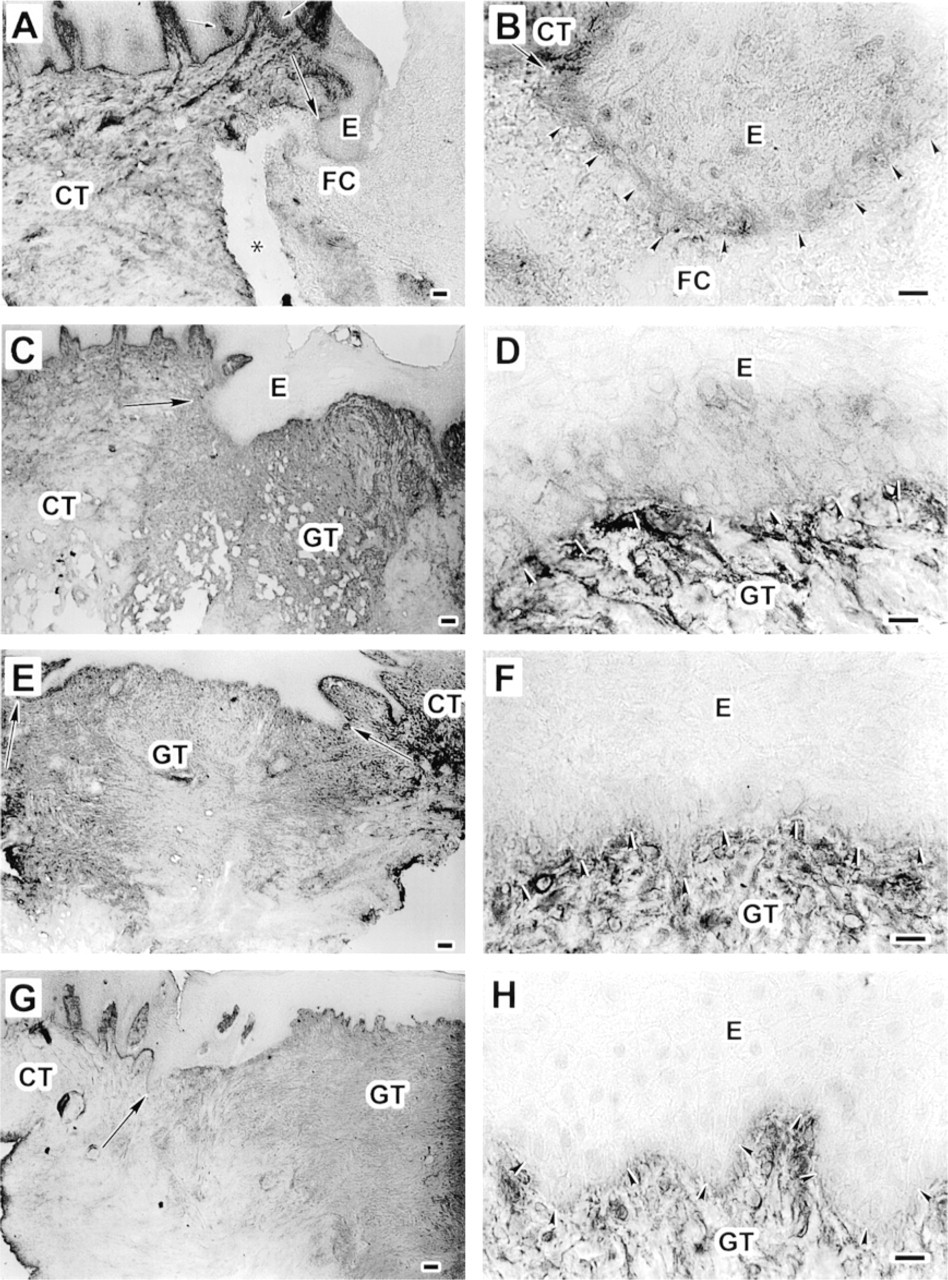
Immunolocalization of tenascin-C using monoclonal antibody BC-4 that recognizes all isoforms of tenascin-C in 3- (
Cell culture and in situ hybridization findings showing that migratory keratinocytes express TN-C mRNA in early wound repair suggest that, at this point, TN-C is produced by the migratory keratinocytes (Aukhil et al. 1996; Latijnhouwers et al. 1997). The findings in the present study showed that immunoreactivity for TN-C, including the large molecular weight isoform, was strongly localized in the granulation tissue when the epithelial sheet had completely covered the wound bed 7 days after wounding. At this time, granulation tissue is populated by proliferating fibroblasts (Clark 1993) and the expression of TN-C mRNA by keratinocytes is downregulated, whereas it is upregulated in fibroblasts (Mackie et al. 1988; Aukhil et al. 1996; Ramos et al. 1997). The differential spatiotemporal expression pattern of TN-C at the epithelial–connective tissue interface and in the granulation tissue suggests that the inductive signals for expression of TN-C by keratinocytes and fibroblasts are probably different. In fact, in vitro studies indicate that TN-C deposition by fibroblasts during early wound repair is probably induced by the migrating epithelial cells (Mackie et al. 1988). Cell culture findings suggest that, during the later phases of wound healing, TN-C expression by fibroblasts may be induced by cell proliferation. In fact, in fibroblasts, cell proliferation is specifically associated with induced expression of the larger TN-C isoform relative to the smaller one (Borsi et al. 1994). In addition, tensional force created between fibroblasts and extracellular matrix can upregulate TN-C expression (Chiquet–Ehrismann et al. 1994). Expression of α-smooth muscle actin is induced in fibroblasts that are actively involved in cell-mediated wound contraction (Darby et al. 1990). In our wound healing model, we have observed early expression of α-smooth muscle actin in fibroblasts at 7-day-old wounds, with a peak expression at 14 days after wounding (unpublished observation), which coincided with the strong TN-C expression in the granulation tissue.
During wound repair, immunoreactivity for the integrin α9-subunit was upregulated at the basal cell membrane of the migrating basal epithelial cells in 3- and 7-day-old wounds. Interestingly, in areas of 7-day-old wounds at which epithelial migration had stopped and epithelium had started to differentiate, and in 14-day-old wounds in which epithelial maturation was under way, expression of α9 integrin was relatively weak by the epithelial cells. This suggests that α9 integrin functions in cell migration in early wounds, whereas its expression must be downregulated during epithelial differentiation. In corneal wound repair, α9 integrin also localizes to the migrating corneal epithelial cells (Stepp and Zhu 1997). Interestingly, during murine embryonic development, α9 integrin expression coincides with epithelial stratification (Wang et al. 1995). During wound repair, epithelial stratification starts at the migratory cell front abundantly expressing α9 integrin, suggesting that α9 integrin has a similar function during embryonic development and wound repair.
The expression of α9 integrin in migrating epithelial cells coincides with expression of TN-C under the migrating epithelial front and co-localizes with integrin β1-subunit during early wound repair (Juhasz et al. 1993; Larjava et al. 1993; Haapasalmi et al. 1996). Therefore, α9β1 integrin may be one of the cell surface receptors that mediates cell migration on TN-C-containing provisional matrix during wound re-epithelialization. On the other hand, in addition to regulating cell migration, TN-C can also induce cell proliferation. The mitogenic activity appears to be associated with a region in the fibronectin Type III domain that is recognized by α9β1 integrin (End et al. 1992). In fact, heterologous expression ofα9β1 integrin in cells plated on this TN-C fragment induces cell proliferation (Yokosaki et al. 1996). Therefore, it is also possible that interaction between α9 integrin and TN-C may be one of the mechanisms that regulate epithelial cell proliferation during wound repair.
Downregulation of α9 integrin in the wound epithelium coincided with induced expression of αvβ6 integrin from Day 7 to Day 14 after wounding. The expression of αvβ6 integrin temporarily and spatially overlapped with the expression of TN-C at the cell–cell interfaces at the wound epithelium. During granulation tissue reorganization and remodeling between Days 7 and 28, the expression of αvβ6 integrin and TN-C in the wound epithelium was gradually downregulated. Expression of integrins at the cell–cell contacts is a common finding, although no ligands for integrins are normally present. Co-localization of TN-C and αvβ6 integrin at the cell–cell interfaces at the wound epithelium suggest that, at least during wound repair, integrins may also bind their ligands at the epithelial cell interfaces. The finding that heterologous expression of α9β1 integrin in cells plated on TN-C induces, whereas β6 integrin downregulates, cell proliferation (Yokosaki et al. 1996) suggests that the switch in the expression of α9 and αvβ6 integrins in the wound epithelium may be one of the mechanisms that regulate cell responses during epithelial differentiation.
Previous studies after wound repair up to Day 7 after wounding have demonstrated induced αvβ6 integrin expression by wound epithelial cells (Breuss et al. 1995; Haapasalmi et al. 1996). Accordingly, in the present study, peak expression of αvβ6 integrin was detected in 7-day-old wounds. However, after 14 days, expression of αvβ6 integrin was still upregulated in the epithelium covering the granulation tissue. At 28 days after wounding, when the organization of the wound granulation tissue was more complete, the wound epithelium became completely negative for αvβ6 immunoreativity. Findings from studies using mice with targeted disruption or constitutive overexpression of β6 integrin gene indicate that epithelial αvβ6 integrin expression may downregulate inflammation in the underlying connective tissue (Huang et al. 1996, 1998). In addition, the ability of αvβ6 integrin to activate latent TGF-β (Munger et al. 1999) suggests that it may play a role in the activation of this growth factor to stimulate matrix production by fibroblasts in the wound granulation tissue. In fact, on the basis of our histochemical staining (not shown), collagen deposition in the granulation tissue coincided with expression of αvβ6 integrin in the epithelium.
Our findings in this study showed expression of α9 integrin by inflammatory cells in the provisional matrix of the blood clot after 3 days of wounding. Recent findings have shown that α9β1 integrin selectively expressed on neutrophils mediates their adhesion on activated endothelial cells through binding to VCAM-1, which mediates chemotaxis of neutrophils on endothelial cells (Taooka et al. 1999). Therefore, our findings suggest a role for α9 integrin in the extravasation of neutrophils in the acute inflammatory phase of wound repair.
The integrin α9-subunit was relatively highly expressed in basal and suprabasal cells of normal nonkeratinized epithelium of ventral tongue and buccal mucosa, whereas it showed a weaker expression in keratinized epithelium of gingiva, in which it localized mostly to the basal cells. The regional variations in keratinization and differentiation of oral mucosa affect also the expression pattern of other integrins in the oral epithelium. In normal keratinized oral mucosa, expression of integrins is mainly confined to the basal epithelial cells that express α2β1, α3β1, α6β4, and αvβ5 integrins (Jones et al. 1993, 1997; Larjava et al. 1993; Thorup et al. 1997), whereas in normal non-keratinized oral mucosal epithelium these integrins are also expressed by suprabasal cells (Jones et al. 1993; Thorup et al. 1997). In primates, oral epithelial cell proliferation is highest in nonkeratinized and is slowest in the keratinized epithelium (Rowat and Squier 1986), suggesting that increased suprabasal expression of α9 integrin in human nonkeratinized mucosa may be associated with enhanced proliferation of the epithelial cells. In fact, in conditions such as wound healing or psoriasis, rapid cell proliferation is associated with increased integrin expression in suprabasal cell populations (Hertle et al. 1992; Jones et al. 1993, 1995; Moles and Watt 1997; Rikimaru et al. 1997). Suprabasal localization of α9 in these processes is identical to that of β1 integrin (Larjava et al. 1993) and suggests that suprabasal expression of α9β1 integrin may play a role in the regulation of cell proliferation and differentiation during tissue maintenance.
In the basal epithelial cells of normal keratinized and nonkeratinized oral mucosa, α9 integrin showed the most abundant staining against the basement membrane. TN-C immunoreactivity for both BC-2 and BC-4 antibodies was also localized at the basement membrane zone in all areas of oral mucosa, indicating that the large TN-C isoform is also normally expressed in oral mucosa. Electron microscopic localization of TN-C in rat skin indicates that TN-C is actually a component of the basement membrane (Mackie et al. 1988) and may therefore serve as a ligand for α9 integrin. Interestingly, TN-C showed differential localization in the connective tissue of keratinized gingiva and in nonkeratinized buccal and ventral tongue mucosa. In gingiva, TN-C was expressed throughout the connective tissue matrix but in only few vascular basement membranes. In contrast, TN-C was mainly localized at the epithelial and vascular basement membranes in the nonkeratinized oral mucosa but relatively weak staining was detected in the connective tissue. It is possible that the differential TN-C expression in various regions of oral mucosa may reflect functional differences in these tissues.
Previous findings have shown that α9 integrin is temporarily expressed in vascular smooth muscle cells during murine development (Wang et al. 1995). In contrast, α9 integrin is not usually expressed by endothelial cells during development or in adult tissue (Palmer et al. 1993; Wang et al. 1995). We localized α9 integrin at the blood vessels of normal oral mucosal connective tissue and in wound granulation tissue. Although we did not specifically show if α9 integrin expression localized to vascular smooth muscle cells or endothelial cells, the finding that α9 integrin is expressed at blood vessels in adult human oral mucosa and in wound granulation tissue is new, and it suggests that vascular expression of α9 integrin may be regulated in a tissue-specific manner.
Taken together, our findings suggest that interaction between α9 integrin and TN-C isoforms, including the large molecular weight isoform, may play a role in the maintenance and differentiation of normal keratinized and nonkeratinized oral mucosa. Furthermore, the finding that TN-C isoforms and their putative receptors, α9 and αvβ6 integrins, are spatially and temporally coordinately expressed during wound repair, suggest that interactions of these integrins with TN-C may be among mechanisms that regulate epithelial cell proliferation, migration, and phenotype during wound repair.
Footnotes
Acknowledgements
Acknowledgments
Supported by grants from the Medical Research Council of Canada.
We would like to thank Cristian Sperantia, Andre Wong and Bruce McCaughey for expert technical assistance.