
Abstract
Zinc ions play an important role in testis development and spermatogenesis. Thus, nutritional zinc deficiency leads to aberrant testicular development, reduced spermatogenesis, and male sterility. The precise actions of zinc in mediating these functions and the mechanisms by which zinc is itself regulated in the testis, however, have not been adequately elucidated. We have assessed the distribution of the zinc-regulating proteins ZnT-1 and metallothionein I/II (MT I/II) in the mouse seminiferous tubule. Colabeling for ZnT-1 and MT I/II demonstrated unique patterns of distribution for these proteins, with ZnT-1 present in Sertoli cells in addition to luminal spermatozoa and MT I/II restricted to spermatocytes. These findings were confirmed by dual-label immunofluorescence for ZnT-1 and the Sertoli cell marker, vimentin, and by immunoelectron microscopy. The differential expression patterns of ZnT-1 and MTs support the hypothesis that ZnT-1 and MTs play different roles in the regulation of intracellular zinc in this organ. The specific expression of ZnT-1 in the Sertoli cells, moreover, is consistent with their role in maintaining a nurturing, closely regulated environment for spermatogenesis.
Z
The functional importance of testicular zinc is demonstrated by the severe consequences of zinc deficiency on testicular development and function. Zinc deficiency is known to result in hypogonadism, inhibition of spermatogenesis, and defects in the morphology of spermatozoa (Hidiroglou and Knipfel 1984). In addition, severe zinc deficiency leads to necrosis of germ-cell precursors that may lead to tubular atrophy and abnormal differentiation of spermatids (Merker and Gunther 1997). Zinc is also known to provide protection against the toxic effects of cadmium (Gunnarsson et al. 2004) and lead (Batra et al. 2004), although the precise mechanism by which this is accomplished is unknown.
In the rodent testis, histochemical staining for chelatable zinc in testicular slices has shown that chelatable zinc is found in the cytoplasm and smooth endoplasmic reticulum of spermatagonia and of spermatocytes at all stages of development, as well as occasionally in Leydig cells in the interstitial tissue (Sorensen et al. 1998). The same basic distribution was described earlier by Vera-Gil et al. (1991) using 65Zn and autoradiography. The presence of chelatable Zn2+ ions in spermatocytes has been suggested to play an essential role at the onset of meiosis (Sorensen et al. 1998). The requirement for zinc in the proper development of testicular germ cells must be considered together with the known toxicity of zinc to many cell types. A precise and reliable system for the regulation of intracellular free zinc (Zni) is clearly required. Regulation of Zni is likely mediated by a combination of factors, including zinc transporter proteins that either extrude (McMahon and Cousins 1998) or otherwise mediate influx or accumulation of cellular zinc (Gaither and Eide 2000, 2001), and metallothioneins (MTs) that essentially buffer intracellular zinc (Mehra and Bremner 1987; De et al. 1991). Although the expression and action of testicular MTs have been documented, their function has primarily been related to protection against cadmium toxicity (e.g., Liu et al. 2001; Ren et al. 2003). In terms of other proteins known to regulate zinc, little is known, e.g., regarding the distribution and function of zinc transporter in the testis. ZnT-1 belongs to a family of zinc transporters, i.e., ZnT-1 through 9 (see Liuzzi and Cousins 2004 for review). These proteins are characterized by multiple membrane-spanning regions and a histidine-rich intracellular loop (McMahon and Cousins 1998). Gene and protein expression of the first of these to be described, ZnT-1, are upregulated by acute increases in dietary zinc (McMahon and Cousins 1998). Expression of ZnT-1, moreover, has been reported to confer resistance against zinc toxicity (Palmiter and Findley 1995). In preliminary experiments, we observed that ZnT-1 protein is present in the testis of neonatal mice (unpublished observations). We report here on the testicular expression and spatial distribution of ZnT-1 and MTs in the mouse seminiferous tubule.
Materials and Methods
Histology and Immunohistochemistry
All procedures involving animals were approved by the University Animal Care and Use Committee. Under ketamine/xylozine anesthesia, 10-week-old CD-1 mice were perfused through the left cardiac ventricle with 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS). The testes were removed, left in fresh fixative solution at 4C overnight, and paraffin embedded. Sections (8 μm) were cut, mounted onto slides, then deparaffinized and trypsinized in preparation for immunohistochemistry or immunofluorescence. For the former, the sections were first immersed for 10 min in a solution containing 30% H2O2 and 10% methanol to block endogenous peroxidase. This was followed by extensive rinses in PBS, and 30-min incubation in a blocking solution containing 4% normal goat serum, 1% bovine serum albumin, and 0.5% Triton-X-100 in PBS. For immunofluorescence, the endogenous peroxidase-blocking step was excluded.
The polyclonal antibody prepared against ZnT-1 was produced as described elsewhere (Sekler et al. 2002). It was used diluted in 0.1 M PBS with 0.01 M Triton-X-100 to 1/500. The monoclonal mouse anti-goat MT I/II (E9; DAKO, Denmark) antiserum was used at 1:200; anti-H5 (vimentin; Developmental Studies Hybridoma Bank) at 1:100. Visualization of the antibody complex was achieved using the avidin-biotin-HRP method (Vector Labs; Burlingame, CA). For immunofluorescence, MT I/II and vimentin were visualized by means of a Cy3-conjugated, donkey anti-rabbit IgG (Jackson Labs; West Grove, PA) diluted 1:100 in PBS; ZnT-1 by Cy2-conjugated goat anti-rabbit IgG (Jackson Labs) diluted 1:50 in PBS. Immunostained slides were examined in a laser confocal microscope using appropriate excitation and barrier filters. Images were captured digitally and processed in a Macintosh computer using NIH Image 1.61 (Wayne Rasband, NINDS, NIH).
To ensure that ZnT-1 immunoreactivity was selective, sections were processed for immunohistochemistry and immunofluorescence with the ZnT-1 antiserum as described above, after it was preincubated for 16 hr with the preimmune serum. Other sections were incubated in normal rabbit serum without the primary ZnT-1 antibody and subsequently processed normally with Cy2-conjugated secondary antiserum for immunofluorescence and biotin-conjugated IgG and avidin-biotin-DAB as described above for immunohistochemistry.
Immunoelectron Microscopy
Immunohistochemistry for ZnT-1 and MT1/II (see above) was carried out on 50-μm-thick sections cut on a vibrating microtome. The immunolabeled tissue was postfixed with 1% OSO4 for 30 min and stained en bloc with 1% uranyl acetate for 60 min and dehydrated in ethanols to propylene oxide before embedding flat in epon-araldite between glass slides. Ultrathin (∼80 nm) sections were cut on a Reichert-Jung ultramicrotome and mounted on copper grids for analysis at 60 kV in a JEOL 100SX transmission electron microscope. Negatives were developed and scanned into Adobe Photoshop for contrast/brightness enhancement and resizing.
Western Blots
Olfactory bulbs, neocortex, cerebellum, striatum, thalamus, hypothalamus, and hippocampus were microdissected and maintained throughout the procedure on ice in a buffer containing 250 mM sucrose, 50 mM NaCl, and 10 mM Hepes buffer, together with a protease inhibitor mixture (Complete, Boehringer Mannheim). Tissue was then sonnicated, frozen, and stored at −70C until used. Protein concentration was determined using a modified Lowry protocol (Markwell et al. 1978). The SDS-page gels and Western blots were performed as described previously (Sekler et al. 1995). Rabbit anti-ZnT-1 was used at a dilution of 1:500. The immunostain was detected using a peroxidase-conjugated, donkey anti-rabbit antibody (Amersham; Piscataway, NJ) using the chemiluminescense reagent (Pierce Ltd.; Rockford, IL) (Sekler et al. 1995). Analyses from different brain regions were performed four to five times. As a control, the antiserum was preabsorbed with 0.4 mg of the immunizing peptide as described previously (Maunsbach et al. 1997).
Results
ZnT-1 Is Highly Expressed in Testis Preparations
Initial efforts to characterize ZnT-1 distribution in mouse testis employed immunoblots of total testicular mouse extracts using the antibody generated against the C-terminal domain of ZnT-1 (Sekler et al. 2002). As shown in Figure 1A, the antibody stained a single, 60-kD peptide corresponding to the expected molecular weight of ZnT-1. As shown in Figure 1B, preabsorption of the antiserum completely abolished the staining of the 60-kD band, indicating that the antibody binds selectively to testicular ZnT-1.
Chelatable Zinc and Zinc-homeostatic Proteins in the Seminiferous Tubule
In the rat, developing germ cells accumulate or reduce intracellular zinc concentrations during different stages of spermatogenesis (Sorensen et al. 1998). We hypothesized that these Zni changes are mediated in part by ZnT-1 and MT I/II. To test this hypothesis in the CD-1 mouse, we characterized the distribution of ZnT-1 and MT I/II in paraffin-embedded sections.
Figure 2 shows the distribution of Cy2- and Cy3-labeled ZnT-1 and MT I/II proteins, respectively, in the mouse testis. Intense ZnT-1 immunofluorescence was observed at all levels of the tubule, in Sertoli cell cytoplasm (particularly, but not exclusively along the inner plasma membrane) adjacent to cell profiles at the basal portion of the tubule including spermatocytes and round spermatids. Cells in the interstitial tissue also exhibited ZnT-1 immunoreactivity, as did luminal spermatozoa. Immunofluorescence in the interstitial space, however, was also observed in sections incubated with primary antiserum initially exposed to preimmune serum (Figures 2D and 2E) and even without primary antiserum (data not shown), indicating that this labeling was nonspecific. Immunofluorescence for MT I/II was most intense in spermatocytes (Figure 2B) at all stages of meiosis. It was absent in spermatids and spermatozoa.
An antibody against the intermediate protein, vimentin, which in the testis specifically labels Sertoli cells (Franke et al. 1979), was employed to confirm that ZnT-1 is indeed labeling Sertoli cells. As shown in Figures 2F and 2G, there was a striking overlap between the distributions of ZnT-1 immunoreactivity and the Sertoli cell marker. This result was also obtained (data not shown) using an antiserum against another cytoskeletal marker of Sertoli cells, NF200 (Davidoff et al. 1999).
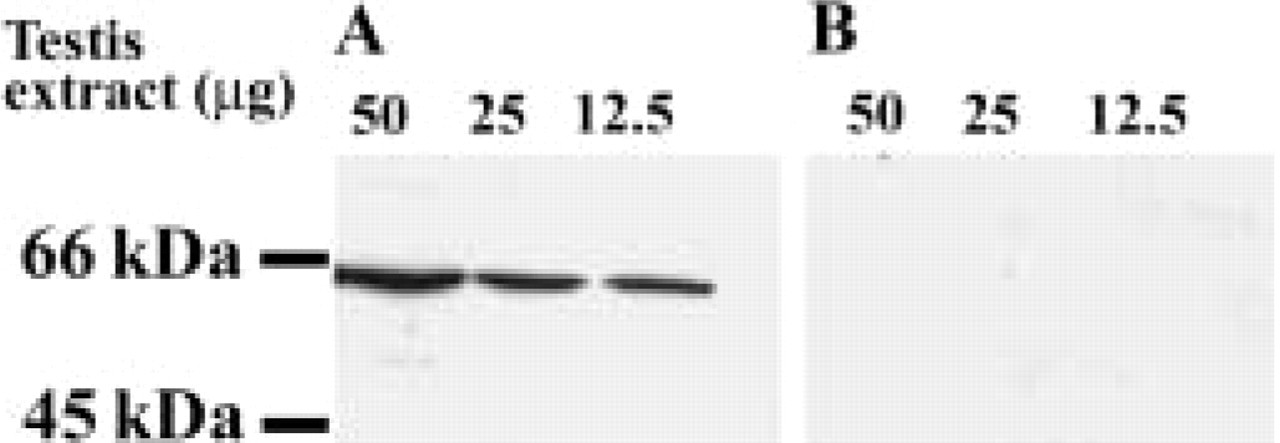
ZnT-1 expression in testis homogenates. (
Figure 3 shows ultrastructural localization of ZnT-1 in the seminiferous tubule. Immunoreactive ZnT-1 was observed in Sertoli cell perikarya in the plasma membrane throughout the tubule wall. Spermatagonia at the adluminal side of the tubule were invariably unlabeled also, as were spermatocytes and the smaller round spermatids, which exhibited the distinctive beginning of an acrosome on one side of the nucleus (Figure 4B). Elongated or rectangular-shaped spermatids containing varying amounts of cytoplasm, in contrast, exhibited dark reaction product along their plasma membrane. In the lumen of the tubules, the tails and part of the body of many of the spermatozoa exhibited a thin deposit of DAB precipitate (Figures 4E and 4F).
In sections immunolabeled for MT I/II, Sertoli cell cytoplasm between developing spermatogenic cells was clear and free of DAB reaction product (Figure 4). Spermatocytes were frequently MT-immunoreactive, however, with the DAB distributed homogeneously in their cytoplasm (Figures 4B and 4C).
Taken together, our results demonstrate that ZnT-1 and MT I/II are expressed by separate cell types in the seminiferous tubule of the CD-01 mouse: ZnT-1 in Sertoli cells and late spermatids and spermatozoa, MT I/II in spermatocytes.
Discussion
Zinc deficiency leads to atrophy of germ cells in the seminiferous tubules and to the failure of spermatogenesis (Hidiroglou and Knipfel 1984; Vallee and Falchuk 1993; Sorensen et al. 1998). Interestingly, it is the later stages of spermatogenesis, i.e., in spermatids, where zinc accumulates at relatively high concentrations. Because of its toxicity to many cell types (Swanson and Sharp 1992; Choi and Koh 1998; Kim et al. 2000), however, strict regulation of Zni is essential and is probably achieved through a delicate functional balance of zinc-regulating elements (see Liuzzi and Cousins 2004 for review). One of these, ZnT-1, is shown here, for the first time, to be expressed by specific cell types in the mouse testis. Interestingly, this protein was localized to Sertoli cells, as well as late-stage spermatids and spermatazoa. Leydig cells in the interstitial tissue were also strongly positive for ZnT-1. In contrast, MT I/II protein was expressed exclusively in germ cells, notably spermatocytes. This is in substantial agreement with De et al. (1991) in CD-1 mice and Tohyama et al. (1994) in rats, who reported that MT I and II gene transcripts were expressed by spermatocytes and were absent in Sertoli and interstitial cells. In contrast, Danielson et al. (1982) reported that Sertoli and interstitial cells expressed the gene encoding MT I/II.
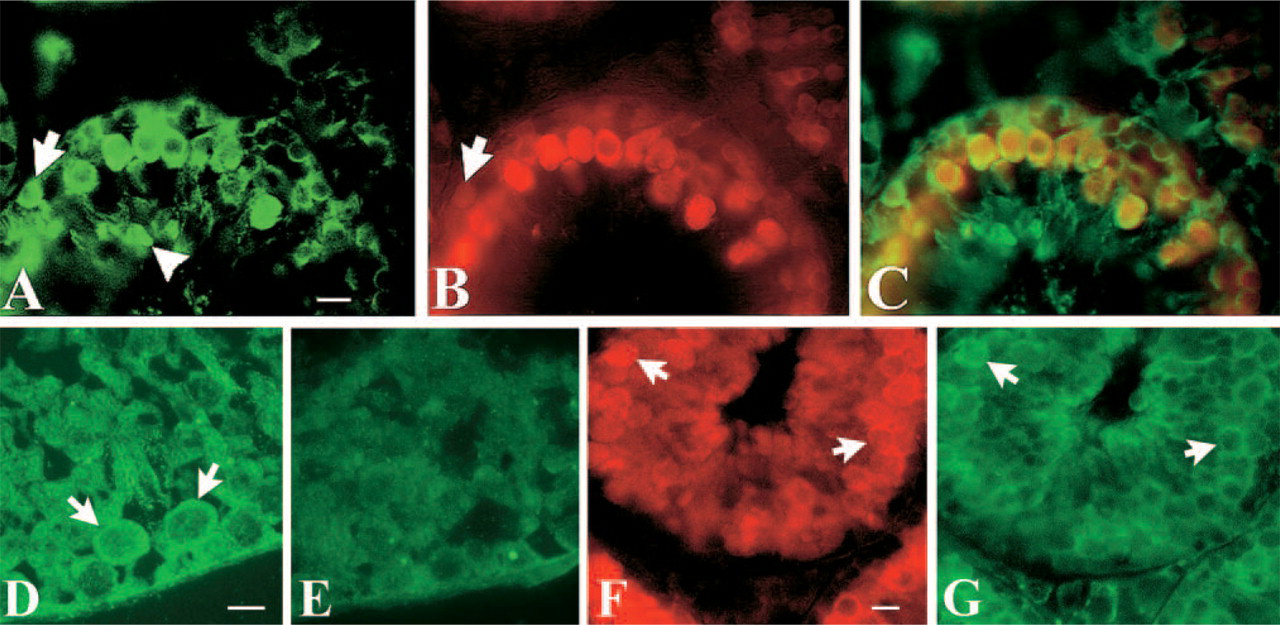
Confocal imaging of Cy2-conjugated IgG-labeled ZnT-1 and Cy3-conjugated IgG-labeled MT I/II. (
Although zinc plays a key role in testicular physiology, no zinc transporter protein has previously been demonstrated in this organ. Gene transcripts for ZnT-3 and ZnT-4 have been identified in testis, e.g., by Northern blots (Palmiter and Findley 1995), but expression of the corresponding proteins has not been directly demonstrated. This is consistent with other reports showing a lack of correlation between ZnT transcript and protein levels (Wenzel et al. 1997; McMahon and Cousins 1998) and is also borne out functionally. Mutant mice with a targeted deletion of the ZnT-3 gene, for example, showed dramatically lower chelatable zinc content in the brain but exhibited no change in testicular zinc levels (Cole et al. 1999). For this reason, we have focused on the testicular expression of the ZnT-1 protein, which is ubiquitous, and the only zinc regulator known to be in the plasma membrane (McMahon and Cousins 1998). In fact, the present study is the first to directly examine the subcellular distribution of this protein—in any tissue. Our findings indicate that ZnT-1, although indeed present in the plasma membrane, is also present in the cytoplasm of the Sertoli cells. Some of the labeled protein is probably there as a result of our use of detergents such as Triton-X-100 (see Figure 3D), but a cytoplasmic localization has been described for ZnT-1 by us (Nitzan et al. 2002; Sekler et al. 2002) and by others as well (e.g., Kelleher and Lönnerdal 2003; Liuzzi and Cousins 2004). Indeed, much of this non-plasma-membrane-bound ZnT-1 appears to be associated with membranes, including the nuclear membrane, various vesicles, and the Golgi apparatus within Sertoli cells. It will be interesting to determine whether there is a dynamic process through which ZnT-1 can move between these two cellular compartments.
In the testis, protecting developing germ cells from toxic concentrations of zinc (and other heavy metals) is an obvious priority. This is true as well for neurons in the brain (Choi and Koh 1998) and in pancreatic beta cells (Kim et al. 2000). As the key mediator of the environment for germ cell development, Sertoli cells are likely to be the most critical element in zinc regulation. Nevertheless, MT I/II, as well as other zinc-binding proteins, undoubtedly participate in regulating intracellular zinc either via the Sertoli cells or directly, in the germ cells. The relatively high zinc content of spermatids (Sorensen et al. 1998), for example, could indicate the involvement of a zinc importer, e.g., ZIP (Gaither and Eide 2001), that catalyzes the influx of zinc into cells.
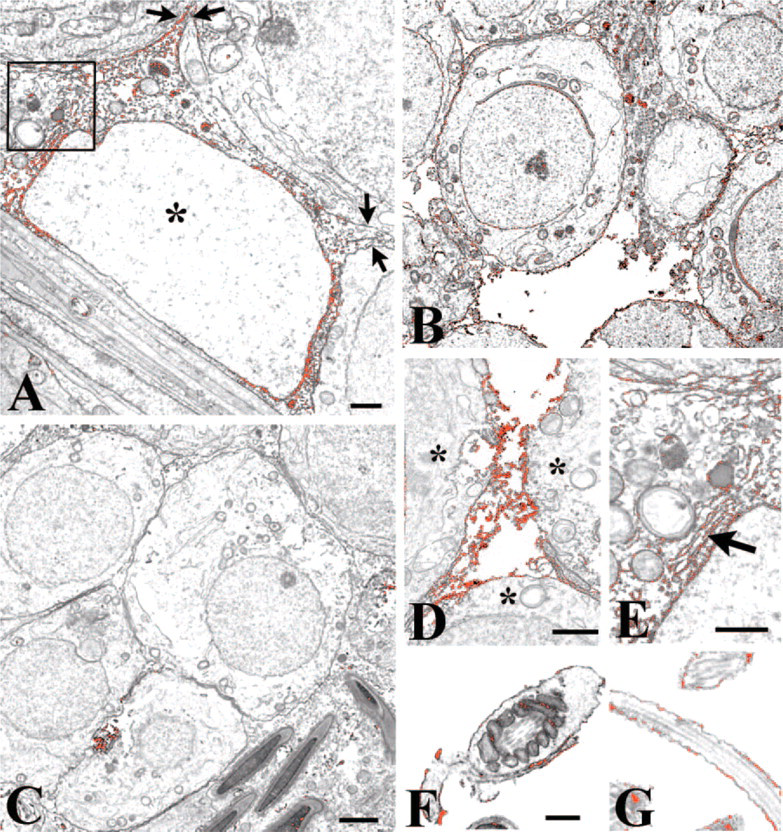
Immunoelectron microscopy for ZnT-1. DAB precipitate has been pseudocolored red to facilitate identification of labeling. Testicular cells were analyzed following avidin-biotin immunolabeling for ZnT-1, embedding in Epon-Araldite resin, and ultrathin sectioning onto copper grids. Labeling was restricted to Sertoli cell cytoplasm and luminal spermatozoa. The red flocculant material in each frame represents the ZnT-1-IgG-biotin-avidin-HRP complex. (
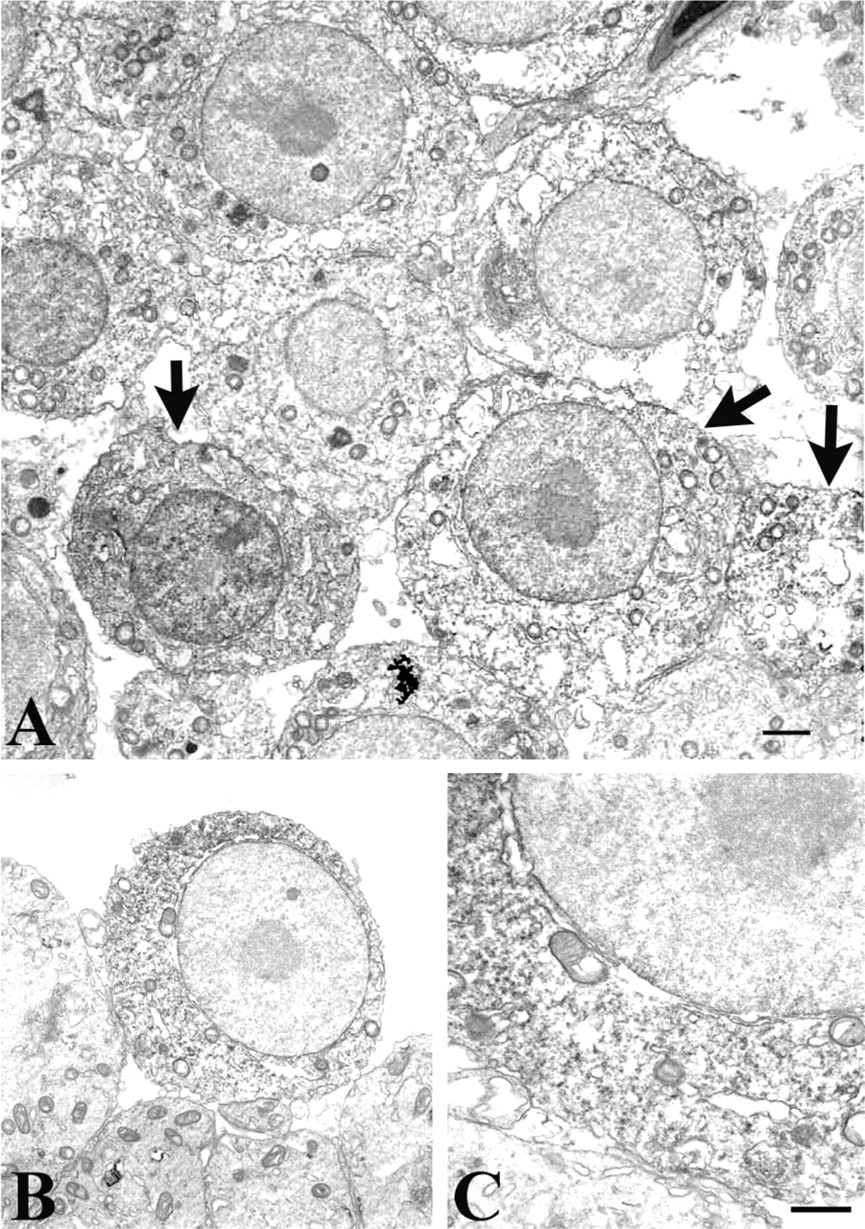
Immunoelectron microscopy for MT I/II. Primary and secondary spermatocytes were the principal cells labeled with the monoclonal antibody to MT I/II. (
Previous studies have examined the relationship of testicular MT to resistance to the toxic environmental contaminant, cadmium (Cd). MTs have been thought to have a detoxifying effect on heavy metals by sequestering them using their numerous cysteine residues (Nordberg and Nordberg 2000). Exposure to Cd has increased dramatically in modern times with the advent of commercial zinc mining, smelting, and cigarette smoking, among other causes. Cadmium accumulates in many tissues, particularly liver, kidney, and testis (Liu et al. 2001). In the testis, Sertoli cells appear to be particularly sensitive to Cd cytotoxicity, which disrupts the inter-Sertoli tight junctions, affecting spermatogenesis. Cadmium is also carcinogenic and can cause hemorrhaging in the capillaries of the testis, inducing degeneration and necrosis of the seminiferous tubules (Nolan and Shaikh 1986). The role of metal-binding proteins such as MTs in the species- and cell-specific nature of Cd toxicity in the testis has been extensively studied in recent years. It appears that in some species and in selected strains within those species, MT I and II are constitutively expressed in Sertoli cells (Ren et al. 2003), while in others they are not (De et al. 1991). A protective effect of MT with regard to Cd has not yet been definitively demonstrated (see Dalton et al. 1996). In contrast, pretreatment with zinc has long been known to counter the destructive actions of the heavy metal on testis (Waalkes and Perantoni 1988). It may be significant, therefore, that ZnT-1, possibly activated by the zinc-finger protein, metal response element-binding transcription factor (MTF) (Daniels et al. 2002), is rapidly and robustly induced by both zinc and cadmium (Langmade et al. 2000). We have recently observed, moreover, that ZnT-1 acts to partially block the L-type calcium channel (Segal et al. 2004). Because this channel, which has been identified and characterized in testis (Goodwin et al. 1997), is also a primary entry point for Zn2+, this effect could represent an important mechanism for regulating intracellular levels of this ion.
It is not hard to conceive of a role for ZnT-1-mediated regulation of Zn/Ca in germ-cell-nurturing Sertoli cells in the context of spermatogenesis. Indeed, Barone et al. (2004) have reported that voltage-gated calcium channels mediate experimentally induced apoptosis of germ cells in the rat seminiferous tubule. Additional experiments are required to test the hypothesis that ZnT-1, acting at L-type calcium channels, is modulating this process, and/or that this protein imparts a degree of resistance to zinc or Cd toxicity.
As mentioned above, transcripts (but not the proteins they encode) for other zinc transporters, e.g., ZnT-3, have been reported previously in germ cells in the testis (Palmiter and Findley 1995). ZnT-3 in the central nervous system encodes a protein required for packaging of ionic zinc into glutamine-containing synaptic vesicles (Palmiter and Findley 1995). What role, if any, ZnT-3 might have in the testis is not clear. But the significant differences in their gene sequences, transcript, and protein distributions and the demonstrated ability of zinc to induce ZnT-1 but not ZnT-3 (Takiguchi et al. 2001) suggest that the two zinc transporters are likely to play quite distinct roles in any tissue where they coexist.
Footnotes
Acknowledgements
This work was supported by the BGNegev Foundation (WFS, IS, MH).
We thank Liza Weinfeld for her technical assistance.