
Abstract
Keywords
Human enteroviruses (Evs) of the picornavirus family include poliovirus, Group A and B Coxsackievirus, echovirus, and enterovirus 68-71. The viral particles are 25-30 nm in diameter with icosahedral symmetry, and consist of a single-stranded positive-sense RNA of 7000-7500 bases and a viral capsid. The capsid contains four proteins, VP1 through VP4. Serotype-specific antigenic sites recognized by neutralizing antibodies are located predominantly in VP1. Such epitopes have also been mapped to VP2 and VP3. Major neutralization epitopes are often conformational, involving interaction of discontinued residues, and are heat-sensitive (Melnick 1996). However, enterovirus genus common epitopes localized to VP1 are not present on the surface of infectious virions. It has been shown that denaturation of enterovirus by heating exposes these group-reactive antigens while destroying the type-specific ones (Hummeler and Hamparian 1958; Melnick 1996).
Enteroviruses are one of the most common and important pathogens of humans. In the United States alone, the Evs are estimated to cause 10 million symptomatic infections annually (Strikas et al. 1986). Enteroviral infection is associated with more than 20 clinically recognized syndromes. This group of viruses has a particular role in myocardial diseases, accounting for half of myocarditis cases (Melnick 1996). An increasing body of evidence has linked enterovirus persistence to dilated cardiomyopathy (DCM) by using nucleic acid hybridization and reverse polymerase chain reaction (RT-PCR) (Bowles et al. 1986; Jin et al. 1990; Archard et al. 1998). It is evident that viral myocarditis may progress to end-stage DCM requiring cardiac transplantation, and enteroviral RNA persistence in the heart detected by molecular biology techniques is a strong predictor of poor prognosis of DCM (Why et al. 1994). Recent studies show that the molecular mechanisms by which the virus impairs heart function involve specific cleavage of cardiac distrophin protein (Badorff et al. 1999). However, the mechanisms of persistent viral infection in the heart are not clear. A simple, rapid, and sensitive in situ assay to detect viral protein expression will be valuable in definitive laboratory diagnosis and further investigation of mechanisms of viral persistence and pathogenesis of DCM. In this communication, we report in situ detection of enteroviral capsid protein VP1 in myocarditis and DCM by an improved immunohistochemical technique. A monoclonal antibody (MAb), 5-D8/1 (Yousef et al. 1987), specific for an epitope conserved on the capsid protein VP1 of the enterovirus genus, was employed for broad detection of different serotypes. This MAb has no reactivity with non-enteroviral strains either belonging or not belonging to the picornavirus family and no crossreactivity with human cardiac tissues. To enhance sensitivity, a recently developed EnVison detection system was used to detect antigen-antibody complex. Using this technique, we are able to detect and localize VP1 in myocardial samples not only from acute but also from chronic heart muscle disease and to demonstrate the presence of VP1 in other tissues of these patients. These results represent a new advance in laboratory diagnosis and understanding of enterovirus-associated myocardial diseases.
Materials and Methods
Human Tissue Samples
Autopsy or endomyocardial biopsy myocardial samples were obtained from 13 patients with histopathologically proven myocarditis (nine cases) or DCM (four cases). Cardiac control tissue was taken from 11 cases of accidental death or noncardiac diseases. Histologically, no abnormality was found in these control myocardial tissues. Tissue samples of other organs, i.e., liver, spleen, lung, kidney, and pancreas from some autopsy cases, were also collected. Myocardial tissues were not available from three fatal myocarditis cases, but their pancreas tissue samples were made available for etiological investigation and were included in this study. All specimens were fixed in 10% formalin and paraffin-embedded for routine histopathology. Some of them were fixed for over 1 year before embedding. Most of the blocks had been stored for 1-10 years before the present examination. These samples were collected from Shanghai Medical University (P. R. China) and consent was obtained from patients or their relatives. Clinically, these patients had a history of suspected enteroviral infection or died during outbreaks of enteroviral heart muscle disease. The histopathologic diagnosis of myocarditis or DCM is based on Dallas criteria (Aretz et al. 1987) and those described by Why et al. (1994).
Immunohistochemical Procedures
Enteroviral group-specific MAb 5-D8/1, peroxidase-conjugated rabbit anti-mouse or swine anti-rabbit Ig, normal mouse IgG2a, pronase, pepsin, serum-free protein, blocking reagents, antibody diluent, EnVision Plus detection, and labeled streptavidin-biotin (LSAB) systems were purchased from DAKO (Cambridge, UK). Anti-Coxsackievirus B3 MAb (IgG2a) and polyclonal antibodies were from Chemicon International (Harrow, UK) and Central Public Health Laboratory (Colindale, UK), respectively. Trypsin with buffer salt tablets was from Sigma Chemicals (Dorset, UK). Immunohistochemical procedures included exposure of antigens, blocking, incubation with primary antibody, incubation with conjugated secondary antibody or with detection systems (EnVison or LSAB), and appropriate wash between steps with Tris-buffered saline (TBS). All incubations were performed at room temperature (RT).
Briefly, tissue sections (5 ±) were dewaxed with xylene and rehydrated with graded ethanol. Endogenous peroxidase activity was blocked with 3% hydrogen peroxidase for 15 min. Antigen exposure was achieved by heat-mediated antigen retrieval. Sections were immersed in 10 mM citrate buffer (pH 6.0) within a plastic slide container, heated three times for 5 min in a microwave oven at full power, and left at RT for cooling (Shi et al. 1991). For comparison, tissue sections were permeabilized by traditional enzymatic digestion with trypsin, pronase, or pepsin according to suppliers' instructions. The tissue sections were blocked with the blocking reagents for 15 min and then incubated with primary antibodies (appropriately diluted to 1:100-1:500 in antibody diluent) for 30 min and washed with TBS. The secondary antibody in the EnVision detection system is goat anti-mouse Ig conjugated with dextran polymer, on which many peroxidase molecules were labeled. The sections were incubated with this reagent for 30 min, washed with TBS, and then reacted with diaminobenzidine substrate chromogen for 5-10 min. The color reaction was stopped by a wash in TBS and distilled water. Finally, mounted sections were examined under a Leitz Dialux 20 EB microscope and confirmed under a Olympus Provis AX70 microscope by two viewers. To compare with the EnVision system, detection of enteroviral VP1 was also performed with the three-stage and LSAB immunoperoxidase staining procedures according to suppliers' instructions. Control experiments includes omission or substitution of primary antibody with isotype- and concentration-matched mouse IgG2a or normal rabbit serum. Further control experiments included absorption of MAb 5-D8/1 with CVB3 before immunostaining.
Expression of Viral Capsid Proteins in Eukaryotic or Insect Cells
The capsid protein PV1 of Coxsackievirus B3 expressed in either eukaryotic or insect cells was used to confirm the specificity of MAb 5-D8/1 by immunofluorescence and western blot. Appropriate viral coding sequences were amplified by polymerase chain reaction (PCR) with high-fidelity pfu polymerase (Stratagene; Amsterdam, The Netherlands) from a cDNA clone of CVB3 (Zhang et al. 1993). Recombinant eukaryotic expression vector containing VP1 gene under the CMV promoter was constructed (pCMV/VP1) and expressed in cultured Vero cells by transfection. For expression in insect cells, recombinant baculovirus transfer vectors containing entire CVB3 coding sequences for structural and functional proteins were constructed. Recombinant baculoviruses containing CVB3 coding sequences were constructed by co-transfection of insect cells (sf9) with the recombinant transfer vectors and linearized baculovirus genomic DNA. CVB3 proteins were expressed under the control of a polyhedrin promoter. Recombinant baculovirus was propagated in sf9 cells after infection. CVB3-infected Vero or Hep-2 cells were used as positive controls.
Western Blotting
A cell lysate was prepared for SDS-PAGE from pCMV/VP1-transfected Vero cells, recombinant baculovirus-infected insect cells, and CVB3-infected Vero cells. VP1 proteins were seperated in a 12% SDS-PAGE, electrotransferred onto a nitrocellulose membrane, and subjected to immunostaining with MAb 5-D8/1 according to methods described previously (Ouyang et al. 1996). A chemiluminescence detection system, ECL (Amersham International; High Wycombe, UK), was used to visualize detected protein bands on X-ray films.
Immunofluorescence
Transfected or infected cells expressing CVB3 proteins were fixed in cold acetone and subjected to immunofluorescence for PV1 detection using MAb 5-D8/1 according to standard protocols (DAKO).
Results
Confirmation of Specificity of MAb 5-D8/1 Using In Vitro-expressed VP1 Protein
The specificity of MAb 5-D8/1 to a broad range of enteroviral serotypes, but not to other viruses and cellular antigens, has been documented (Yousef et al. 1987; Trabelsi et al. 1995). To test its immunoreactivity with in vitro-expressed rather than native viral capsid protein, we expressed VP1 in eukaryotic and insect cells. MAb 5-D8/1 reacted with the entire VP1 molecule expressed in the African green monkey kidney cell line (Vero) or in sf9 insect cells (Figure 1). The immunospecificity of MAb 5-D8/1 with these in vitro-expressed VP1s was confirmed by using immunofluorescence (not shown).
Detection and Localization of Enteroviral VP1 in Myocardium from Patients with Heart Muscle Diseases
Using MAb 5-D8/1 and the improved techniques involving combination of the microwave-based epitope unmasking and EnVision system, enteroviral capsid protein VP1 was detected in duplicate myocardial tissue sections from six of nine myocarditis and three of four DCM cases. The results were reproducible when further sections were tested 2-4 weeks later. Substitution of primary antibody with antibody diluent only or with isotype- and concentration-matched normal mouse IgG2a produced negative results on consecutive sections. Pre-absorption of the MAb 5-D8/1 (0.22 μg/ml) with CVB3 in twofold dilution from 1:2 to 1:32 (106-3 × 104 plaque forming units) for 30 min abolished the immunoreactivity. All control tissues from accidental death or noncardiac diseases were negative. Immunostaining signals were localized in cytoplasm of cardiac myocytes/myofibers. Where there were necrotic and infiltrating lesions in myocardial sections from myocarditis, VP1 was often localized near or around, but not in the center of, necrotic tissue (Figure 2). In general, there were two staining patterns of VP1: strong signals were distributed focally or confluently (in fulminent cases) involving a large area of sections in myocarditis, whereas VP1 signals were found in scattered myocytes in DCM cases (Figure 2).
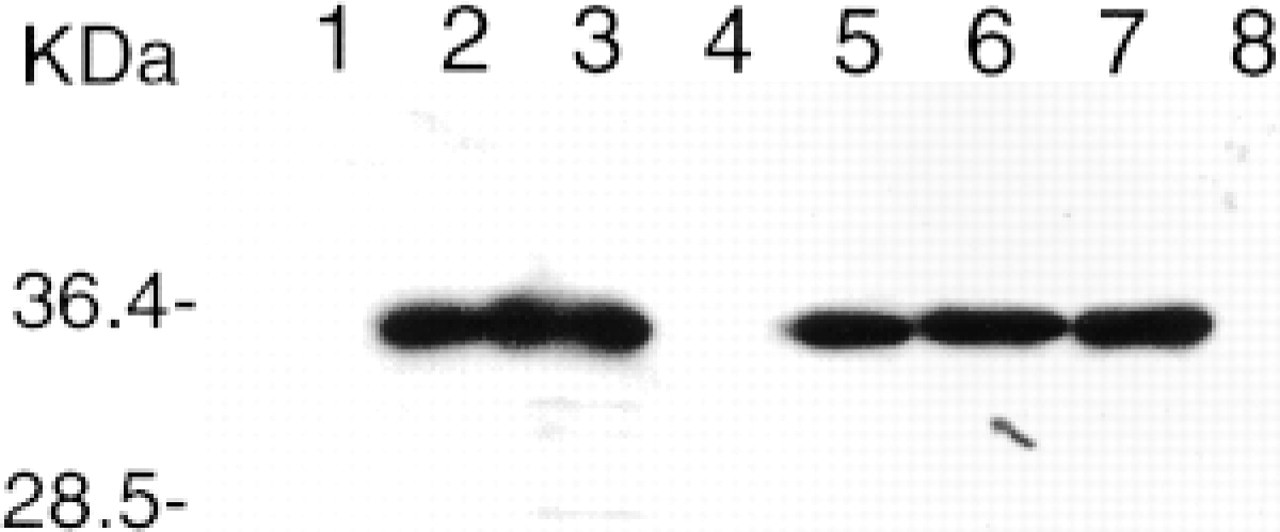
Western blot of in vitro-expressed VP1. The expression of VP1 was described in Materials and Methods. Total proteins of cell lysates were loaded for SDS-PAGE electrophoresis. MAb 5-D8/1 was used at a dilution of 1:500. After application of the ECL detection system, X-ray films were exposed for 10 or 30 sec and developed. Lane 1, mock-infected Vero cells as a negative control; Lanes 2 and 3, CVB3-infected Vero cells as a positive control; Lane 4, pCMVβ/0-transfected Vero cells as a negative control; Lane 5, pCMV/VP1-transfected Vero cells; Lanes 6 and 7, rBV/CVB3-infected sf9 cells; Lane 8, parental baculovirus-infected sf9 cells as a negative control. Molecular weight markers are in kD.
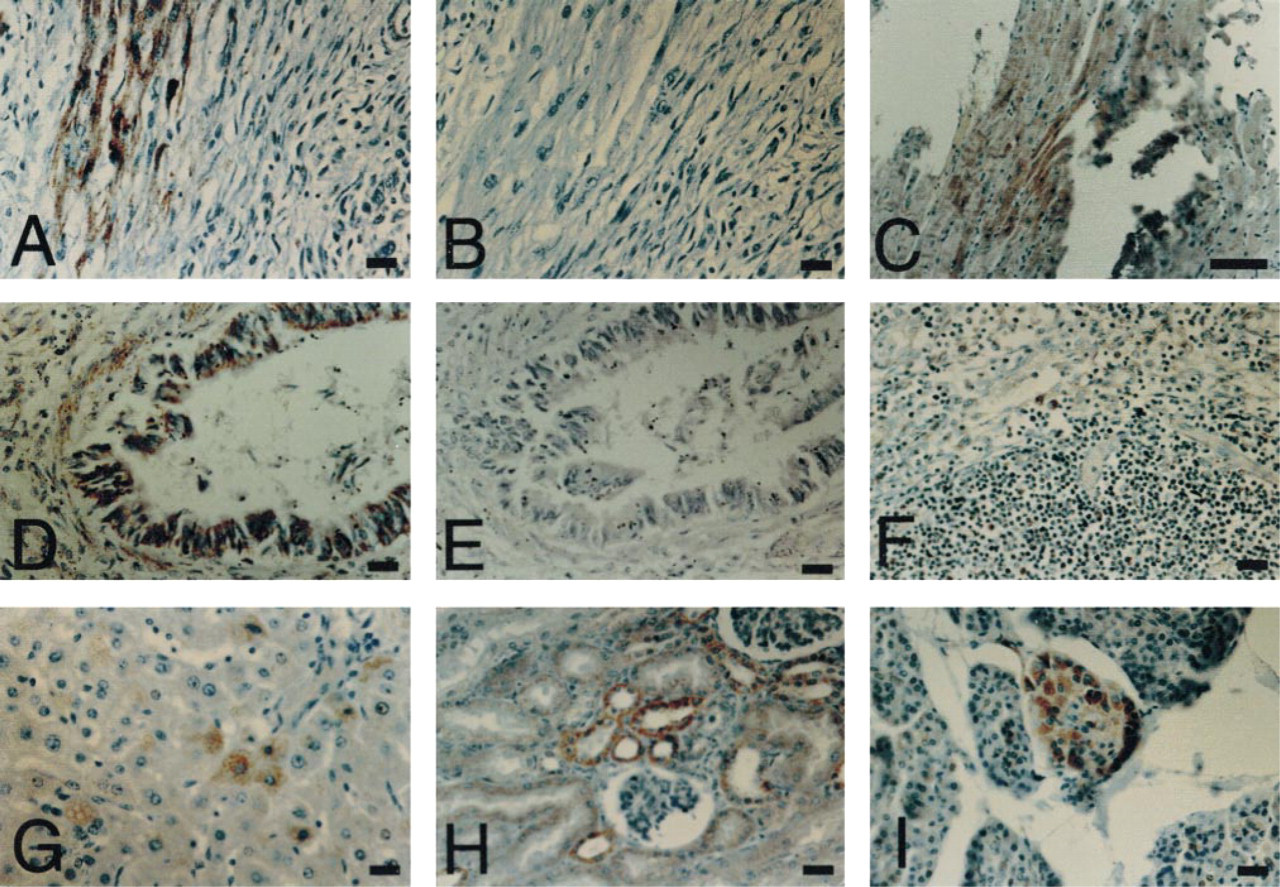
Photomicrographs of VP1 detection in patients with heart muscle diseases. MAb 5-D8/1 and concentration-matched IgG2a were used at a dilution of 1:500. Yellow-brown color indicates the detection of VP1 in tissue sections. (
Detection of VP1 in Other Tissues
Tissue samples of spleen, lung, kidney, pancreas, and liver were collected from a limited number of cases and investigated similarly. VP1 immunostaining was positive in one of two spleen, two of three lung, two of two kidney, three of five pancreas tissue samples from myocarditis. VP1 was also detected in one liver sample from a DCM case. A lung tissue sample from another DCM case was negative. The heart tissue samples from the above cases were all positive for VP1 except for three myocarditis cases, whose pancreas samples were positive but heart samples were not available for the present investigation. VP1 signals were localized in the cytoplasm of various cell types in these different tissues: lymphoid cells in spleen, endothelia in the lung, acinar cells in the pancreas, hepatocytes in the liver, and tubule cells in the kidney. VP1 detection was negative in the spleen, lung, pancreas, liver, and kidney from a control.
Method Comparison
Permeabilization of sections by conventional protease treatment was compared with that by heat treatment in a microwave oven. Consecutive sections were treated with 1 mg/ml trypsin, 0.5 mg/ml pronase, or 4 mg/ml pepsin at 37C for 15-20 min or with microwaving. VP1 signals were seen in sections treated with micro-waving as described above, but not in sections treated with any of these proteolytic enzymes. A different CVB3-specific MAb (IgG2a; Chemicon International, Temecula, CA) and a CVB3-specific rabbit neutralizing antiserum (Public Health Laboratory) were used to compare with the MAb 5-D8/1, and both failed to detect viral antigens in sections pretreated with proteolytic enzymes or microwaving. When different detection systems were compared, signals of VP1 immunostaining in the myocardium generated by the three-stage immunoperoxidase method or LSAB were comparable to that produced by the EnVision system, but a higher concentration of primary antibody was needed and more nonspecific background staining was generated by the former. The EnVision system is simpler, faster, more sensitive, and results in clearer background.
Discussion
Enteroviral capsid protein VP1 was detected reproducibly in heart and other tissue samples from patients with myocarditis or DCM. The high incidence of VP1 detection in this study does not represent a true incidence of enteroviral infection in the heart because the patient groups were highly selected: all had a history of enteroviral infection. Similar results have been reported in selected pediatric patients with fatal neonatal myocarditis (Foulis et al. 1990) or acute idiopathic DCM (Luppi et al. 1998). Further study involving a larger series of patients with myocarditis or DCM is being carried out and is expected to provide representative data on the incidence of VP1 detection in these diseases. Successful detection of VP1 in paraffin sections by using our optimized technique is also due to the specificity and nature of the MAb 5-D8/1. This antibody recognizes a non-neutralizing epitope mapped to residues 40-48 at the N-terminus of VP1 (Samuelson et al. 1995). This part of the protein does not form an exposed domain on the viral capsid (Muckelbauer et al. 1995). Sequence alignment shows that this region is highly conserved among a panel of serotypes of enterovirus. Experimentally, 5-D8/1 was immunoreactive with 36 enteroviral serotypes tested (Trabelsi et al. 1995). It also reacted with VP1 of CVB3 expressed in either eukaryotic or insect cells in Western blotting. This epitope was recognized by MAb 5-D8/1 in formalin-fixed, paraffin-embedded sections after microwave treatment but failed to react with the antibody after proteinase digestion. These results suggest that it is a linear epitope and that its exposure is heat-mediated. It appears that the other MAb and rabbit antiserum to CVB3 are not suitable reagents for detection of viral antigens in paraffin sections, which may explain why detection of enteroviral capsid protein antigens in fixed myocardial tissue was rarely achieved in the past. To increase the signal-to-noise ratio so as to enhance the sensitivity of VP1 detection, the EnVision detection system was employed (Sabattini et al. 1998). In this system, the second antibody was coupled with a dextran backbone to which many enzyme molecules can bind. This is particularly useful in investigation of persistent viral infection in chronic myocardial disease, e.g., DCM.
Pathological changes have been found in organs other than the heart in experimental animal models of viral myocarditis. Pancreatitis is common in mice after inoculation with Coxsackievirus (Zhang et al. 1994). CVB3 was also detected in spleen, lymph nodes, thymus, liver, central nervous system, and lung in mice with CVB3-induced myocarditis (Klingel et al. 1996). However, viral involvement in other organs of myocardial diseases is underestimated because of lack of direct evidence. Measurement of the levels of alanine amino transferase and aspartate amino transferase showed that the hepatic involvement was 72% in enteroviral perimyocarditis (13/18), 30% in DCM (3/10), and zero in acute myocardial infarction (0/10) (Kanda et al. 1995). Most recently, Arbustini et al. (2000) have reported detection of enteroviral genomic RNA by RT-PCR and viral crystalline arrays under an electron microscope in skeletal muscle of two patients who died of fatal myocarditis. In the present study we were able to demonstrate the in situ detection of VP1 in spleen, lung, kidney, pancreas, and liver tissues from some cases of myocarditis or DCM. Despite the fact that only a limited number of cases were investigated, these are the first observation of VP1 detection in fixed tissues from multiple organs other than the heart of patients with myocarditis or DCM. The results suggest that it is advisable to examine multiple organs or tissues in enteroviral myocardial diseases. This will help us to understand the pathogenesis of enteroviral infection, to improve the diagnosis, and will lead to more appropriate management of viral myocarditis or DCM.
In summary, the improved immunohistochemical method, involving MAb 5-D8/1 and EnVision detection system, pioneered in our laboratory is an enteroviral group-specific, rapid, easy, and sensitive assay. It can be used to detect VP1 in archival fixed tissues as old as 10 years. It will become an indispensable measure in routine laboratory diagnosis as well as in mechanistic study of diseases associated with enteroviral infection.
Footnotes
Acknowledgements
Acknowledgments
Supported by the Wellcome Trust (052954Z97), by the University of London Central Fund, and in part by the British Heart Foundation.