
Abstract
Although the intercellular adhesion molecule-1 (ICAM-1) is recognized for its pivotal role in inflammation and immune responses, its role in developmental systems, such as the cyclic growth (anagen) and regression (catagen) of the hair follicle, remains to be explored. Here we demonstrate that ICAM-1 expression in murine skin is even more widespread and more developmentally regulated than was previously believed. In addition to endothelial cells, selected epidermal and follicular keratinocyte subpopulations, as well as interfollicular fibroblasts, express ICAM-1. Murine hair follicles express ICAM-1 only late during morphogenesis. Thereafter, morphologically identical follicles markedly differ in their ICAM-1 expression patterns, which become strikingly hair cycle-dependent in both intra- and extrafollicular skin compartments. Minimal ICAM-1 and leukocyte function-associated (LFA-1) protein and mRNA expression is observed during early anagen and maximal expression during late anagen and catagen. Keratinocytes of the distal outer root sheath, fibroblasts of the perifollicular connective tissue sheath, and perifollicular blood vessels exhibit maximal ICAM-1 immunoreactivity during catagen, which corresponds to changes of LFA-1 expression on perifollicular macrophages. Finally, ICAM-1-deficient mice display significant catagen acceleration compared to wild-type controls. Therefore, ICAM-1 upregulation is not limited to pathological situations but is also important for skin and hair follicle remodeling. Collectively, this suggests a new and apparently nonimmunological function for ICAM-1-related signaling in cutaneous biology.
Keywords
The hair follicle (HF) is the only organ that for the entire lifetime of the mammalian organism shows cyclic switches between periods of massive epithelial cell proliferation and terminal differentiation (anagen), followed by rapid organ involution (catagen) and resting (telogen) (cf. Stenn et al. 1996, 1998; Paus et al. 1998; Paus and Cotsarelis 1999). Murine skin displays a synchronized HF cycle, i.e., all HFs are mostly in the same stage of the hair cycle, whereas human skin displays a mosaic pattern of HF cycling, i.e., HFs in different stages of the hair cycle may reside close to each other in the same skin region. Because both spontaneous and induced HF cycling in murine skin appears in waves, with all HF in almost the same stage of the cycle, the synchronized hair cycle in mice is an excellent model in which to study physiological remodeling of the skin that is associated with distinct stages of the hair cycle (Paus 1996; Paus et al. 1998).
Typical examples for hair cycle-dependent skin remodeling are alterations of the skin and the HF immune system during synchronized HF cycling in mice, such as (a) changes in the number of intraepithelial T-cells and Langerhans cells, (b) changes in the follicular expression patterns of MHC Class I molecules, and (c) changes in contact sensitization responses (Hofmann et al. 1996; Paus et al. 1994a,c, 1998; Paus 1997; Tokura et al. 1997). Furthermore, remodeling of the regressing HFs during catagen is accompanied by phagocytosis of excess basement membrane collagen by macrophages (Parakkal 1969). An active involvement of perifollicular macrophages and mast cells in the catagen-associated skin remodeling has been postulated (Westgate et al. 1991; Paus and Czarnetzki 1992; Paus 1996; Maurer et al. 1997a). Furthermore, potent immunosuppressants can either induce (dexamethasone) or suppress (cyclosporine A, FK506) murine catagen development (Paus et al. 1994b, 1996; Maurer et al. 1997b). Unfortunately, practically nothing is known about the molecular mechanisms that may link HF regression and immunology.
It is reasonable to explore the possible role of cell adhesion molecules in this context. Among the cell adhesion molecules that may be relevant to hair biology (e.g., ICAM-1, NCAM, E- and P-cadherin, and β1-integrins) (Hirai et al 1989; Chuong et al. 1991; Chuong 1993; Kaplan and Holbrook 1994; Müller-Röver and Paus 1998), ICAM-1 is of special interest from both a developmental and an immunological perspective. ICAM-1 acts as a major cell adhesion molecule in a large array of inflammatory and immune responses. Its ligands include lymphocyte function-associated antigen-1 (LFA-1, CD11a/CD18), macrophage differentiating antigen-1 (MAC-1, CD11b/CD18), CD43, fibrinogen, hyaluronan, and rhinoviruses (cf. van de Stolpe and van der Saag 1996). ICAM-1 mediates the adhesion of T-cells to antigen-presenting cells or to target cells, is important for T-cell/T-cell and T-cell/B-cell interactions, and has a co-stimulatory effect on T-cell activation. ICAM-1 is also crucial for the adhesion of monocytes, lymphocytes, and neutrophils to activated endothelium (cf. Springer 1994; van de Stolpe and van der Saag 1996). During inflammatory responses, ICAM-1 expression is a marker for macrophage activation (Goebeler et al. 1990; Leenen et al. 1994). Finally, defects in ICAM-1 have been implicated in the pathogenesis of obesity (Dong et al. 1997).
During human fetal HF morphogenesis, ICAM-1 is transiently expressed on the outer cells of the hair germ (Kaplan and Holbrook 1994). The functional significance of this expression pattern is unknown. However, ICAM-1-deficient mice and K14/ICAM-1-overexpressing mice reportedly do not exhibit gross abnormalities of HF development (Sligh et al. 1993; Williams and Kupper 1994; Xu et al. 1994; King et al. 1995; Davis et al. 1996). Abnormalities of postnatal HF cycling in these mice have not been investigated.
The ICAM-1 ligand LFA-1 belongs to the β2-integrin family and is expressed on most types of white blood cells (cf. Springer 1990; van Kooyk and Figdor 1997). LFA-1 plays a major role in leukocyte-leukocyte and leukocyte-endothelial cell interactions. It is involved in mediating the adhesion of cytotoxic T-cells to their target cells, mixed lymphocyte reactions, and T-cell-dependent antibody response (cf. van Kooyk and Figdor 1997).
To examine the functional role of ICAM-1 expression in HF growth and regression, we have studied the immunohistological expression patterns of ICAM-1 and one of its key ligands, LFA-1, both during neonatal HF development in mice (Paus et al. 1997; Vielkind et al. 1995) and during the depilation-induced murine hair cycle in adolescent C57BL/6 mice (Paus et al. 1990). This was correlated with a semiquantitative analysis of ICAM-1 and LFA-1 gene expression (RT-PCR) in murine dorsal skin during the hair cycle. Finally, we have studied catagen development in ICAM-1-deficient mice.
Materials and Methods
Animals
Syngenic C57BL/6 mice (female, 6–9-week-old and 1–21-day-old neonatal mice; Charles River, Sulzfeld, Germany) as well as homozygous ICAM-1-deficient mice (female, 18-day-old Icamtm1Bay mutant strain; The Jackson Laboratory, Bar Harbor, ME) and age-matched wild-type controls (female 18-day-old, C57BL/6J; Jackson Laboratory) were used. These mice were housed in group cages, with 12-hr light periods, at the Virchow Hospital, Animal Facilities, Berlin, and were fed water and mouse chow ad libitum. Neonatal mice were examined because most pelage HFs in mice develop during the peri- and neonatal period (Vielkind et al. 1995), so that all stages of HF morphogenesis can be conveniently studied in neonatal mice (Paus et al. 1997).
Skin Specimens and Hair Cycle Induction
Dorsal skin was obtained from C57BL/6 mice in various stages of neonatal HF development and adolescent HF cycling (Paus et al. 1997). Depilation-induced anagen (Paus et al. 1990) was compared with spontaneous anagen VI. Using a special skin harvesting and processing technique for obtaining longitudinal cryosections through mouse HFs (Paus et al. 1994a,c), skin samples covered with embedding medium (GSV; SLEE Technik, Main, Germany) were snap-frozen in liquid nitrogen, and stored at −80C until cryosectioning (6 μm). Sections were placed on poly-L-lysine-covered glass slides, air-dried for 1 hr, and fixed in acetone at −20C for 10 min to be stored at −20C until immunohistology was performed.
At least six skin sections each were examined from five mice per hair cycle stage of the depilation-induced hair cycle and from days 1–21 after birth. Therefore, at least 100 different HFs obtained from five mice per stage of HF morphogenesis or cycling were analyzed. At least 10 skin sections and 100 HFs per mouse (= 700 HFs per group) each were examined from seven ICAM-1-deficient and seven corresponding wild-type mice.
Immunohistology
After extensive pilot assays had been performed to determine the most sensitive method for the detection of hair cycle-dependent ICAM-1 expression, the following protocol was found to yield optimal results. Sections were preabsorbed for 15 min at room temperature (RT) with 5% bovine and 5% mouse normal serum in Tris buffer (pH 7.6), followed by 15 min each with avidin and biotin blocking solution (Vecta-Stain kit; Vector Laboratories, Burlingame, CA) at RT. Every step was followed by washing for 15 min at RT (buffer was changed every 5 min). A specific biotinylated monoclonal hamster anti-mouse ICAM-1 antibody (clone 3E2, Scheynius et al. 1993; Pharmingen, San Diego, CA) was used at a dilution of 1:1600 or 1:100 (neonatal skin) in Tris buffer containing 1% bovine normal serum and 4% mouse normal serum for 2 hr at 4C. Cryosections of mouse skin sections incubated with nonimmune hamster IgG at the same concentration as the primary antibody were used as negative controls. Positive controls included (a) cryosections of murine spleen and thymus, (b) inflamed telogen skin after 3 days of topical dimethylsulfoxide treatment (DMSO; 100%), which causes generalized upregulation of ICAM-1 expression via induction of an acute irritant contact dermatitis, and (c) untreated telogen skin with its well-characterized constitutive, orthotopic ICAM-1 expression on cutaneous blood vessels (Goebeler et al. 1990, 1994; Scheynius et al. 1993; Williams and Kupper 1994). The observed antigen distribution was compared to published expression patterns for use as internal positive controls.
The following specific rat anti-mouse antibodies were used at the dilution indicated: anti-LFA-antibody (clone 2D7, Springer et al. 1982; 1:100), anti-MAC-1-antibody (clone M1/70, Springer et al. 1979; 1:8000), anti-γδ-T-cell receptor antibody (clone GL3, Goodman and LeFrancois 1989; 1:75) purchased from Pharmingen; anti-NLDC-145 antibody (clone T-2013, Breel et al. 1987; 1:100) purchased from BMA (Augst, Switzerland); pan-reticular fibroblast marker (clone ER-TR 7, Van Vliet et al. 1986; 1:200) purchased from Camon (Wiesbaden, Germany). The antibodies were diluted in Tris buffer containing 1% normal bovine serum and 4% normal mouse serum and were incubated for 1 hr at RT. After washing for 30 min, sections were then incubated with ABC complex (Vecta-Stain kit) for 45 min at RT and washed again for 30 min at RT (buffer was changed every 5 min). The alkaline phosphatase color reaction was developed as described (Handjiski et al. 1994). For all antibodies, a standard avidin-biotin complex (ABC) method was used as described (Paus et al. 1994a,c; Eichmüller et al. 1996). Anti-MHC Class-II antibodies (clone ER-TR 3, Van Vliet et al. 1984; 1:100) and anti-pan-macrophage-antibodies (clone ER-MP 23, Leenen et al. 1994; 1:150) were purchased from BMA. These antibodies were visualized by a standard alkaline phosphatase-anti-alkaline phosphatase (APAAP) technique as described (Paus et al 1994a,c; Eichmüller et al. 1996). Nonimmune IgG of the host species (at the same concentration as the primary antibody) was used as negative control.
Analysis of HF Regression in ICAM-1-deficient Mice and Wild-type Controls
Age- and sex-matched 18-day-old ICAM-1-deficient and wild-type littermates were used for the analysis of postnatal HF regression (catagen) (Paus et al. 1997). Skin was harvested from defined dorsal truncal regions and 100 HFs per mouse were analyzed by quantitative histomorphometry, as described (Paus et al. 1994b; Maurer et al. 1997a,b).
Data Photodocumentation and Analysis
All immunoreactivity (IR) patterns were analyzed qualitatively and semiquantitatively in at least 100 HFs per stage of HF development or cycling derived from five different mice and were recorded in a computerized schematic representation of all stages of murine HF development and cycling (Paus et al. 1997). For photodocumentation, a Zeiss microscope and an image analysis system (ISIS; Metasystems, Belmont, MA) were used. “Strong” ICAM-1 IR was defined as at least half of the staining intensity of skin blood vessels in untreated telogen skin. Two selected skin compartments [interfollicular dermis, peri-infundibular outer root sheath (piORS)] were also assessed by quantitative immunohistomorphometry for their ICAM-1 expression. The number of ICAM-1+ interfollicular dermal fibroblasts was counted within a standardized microscopic field, which is defined here as a 31-μm-wide region parallel to each follicle and directly below the basement membrane of the epidermis, using an ocular grid (X400 magnification). The quantitative histomorphometric data of the depilation-induced hair cycle (dermal and follicular ICAM-1 expression) and the mean catagen score of ICAM-1-deficient and wild-type control mice were tested by the Mann-Whitney test for nonpaired samples.
RT-PCR
Semiquantitative RT-PCR analysis of ICAM-1, LFA-1, and of constitutively expressed β-actin was performed using the previously described RT-PCR techniques (Pethö-Schramm et al. 1996). Briefly, total RNA was isolated from full-thickness dorsal truncal skin samples of 6–8-week old C57BL/6 mice, using a single-step guanidine thiocyanate-phenol-chloroform method with RNAzol B (Biotech Laboratories; Houston, TX). cDNA was synthesized by reverse transcription of 3 μg total RNA, using a cDNA synthesis kit (Invitrogen; San Diego, CA).
The following sets of oligonucleotide primers were used:
β-actin: 5′-TGG AAT CCT GTG GCA TCC ATG AAA C-3′ and 5′-TAA AAC GCA GCT CAG TAA CAG TCC G-3′
LFA-1: 5′-ACT ATG TAG TGT TGA CCT GGA-3′ and 5′-CTG AGC CCA CCA GGC TTC-3′
(Accession number M60778; Kaufmann et al. 1991); no crossreaction with Mac-1)
ICAM-1: 5′-AGC TAG CGG ACC AGA TCC-3′ and 5′-ATA CAG CAC GTG TTC C-3′
(Accession number X52264; Ballantyne et al. 1989)
Amplification was performed using taq polymerase (GIBCO; Grand Island, NY) over 36 cycles, using an automated thermal cycler (Perkin-Elmer Cetus; Norwalk, CT). Each cycle consisted of the following steps: denaturation at 94C (1 min), annealing at 55C (30 sec), and extension at 72C (1 min). PCR products were analyzed by agarose gel electrophoresis and enzymatic digestion using standard methods (Sambrook et al. 1989). Staining was densitometrically assessed with a video scanner, using Scan Pack 2.0 (Biometra; Göttingen, FRG).
Results
Constitutive ICAM-1 Expression Is a Late Event During Neonatal Mouse Skin and HF Morphogenesis
Using the methodology delineated above, no ICAM-1 IR could be detected in the developing HFs of neonatal mouse skin before Day 19 post partum. Unexpectedly, even the first stronger ICAM-1 expression on neonatal skin vasculature was not detected before Day 7 post partum (Figures 1A and 1B), i.e., at a time when follicle development had far advanced and had reached Stage 5–7 of development (Vielkind et al. 1995; Philpott and Paus 1998). In contrast to adult skin (see below), ICAM-1 IR of endothelial cells in neonatal mouse skin was only weak. Even when a 16-fold higher concentration of ICAM-1 antibody (1:100) was applied than had been used successfully for ICAM-1 detection in adolescent skin (1:1600), neonatal skin sections revealed only faint ICAM-1 expression on endothelial cells of the plexus profundus on Day 1 post partum, which slightly increased on Days 4–7 post partum (Figure 1C).
During this period of neonatal skin and HF development, no ICAM-1 IR was seen in any epithelial or mesenchymal follicle compartment, in the interfollicular epidermal, or in dermal cells (Figures 1A and 1B). The first strong follicle-associated ICAM-1 IR appeared by Day 21 post partum on fibroblasts of the proximal connective tissue sheath of the HF (similar to adolescent catagen development; see corresponding Figures 2E-2G), i.e., at a time when the HF had entered into its first catagen stage, thus initiating HF cycling.
ICAM-1 Expression Is Hair Cycle-dependent and Is Confined to Defined Skin Regions
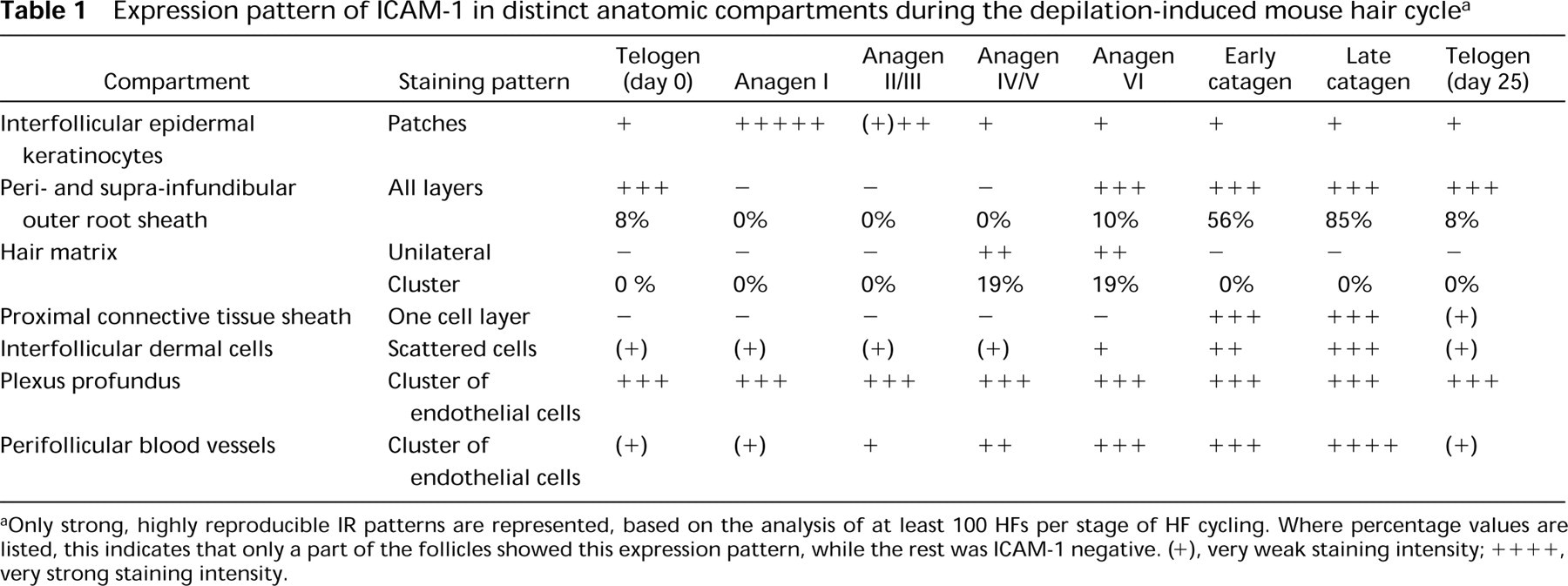
When adolescent mouse skin was examined, it became apparent that, contrary to reports in the literature (Goebeler et al. 1990, 1994; Lenz et al. 1993; Scheynius et al. 1993; Williams and Kupper 1994), constitutive ICAM-1 IR was also present on distinct keratinocyte and fibroblast populations, as summarized in Figure 3. ICAM-1 IR in adolescent mouse skin was strikingly restricted and hair cycle-dependent, and displayed substantial differences between defined anatomic follicle compartments. In addition, individual HFs differed in their ICAM-1 expression pattern, and the percentage of HFs exhibiting intra- and/or perifollicular ICAM-1 IR fluctuated substantially during the murine hair cycle. Analysis of full-thickness cryostat sections from murine back skin incubated with a biotinylated monoclonal antibody against ICAM-1 (1:1600; avidin-biotin complex technique) revealed a dynamic hair cycle-dependent pattern of strong ICAM-1 IR in six defined skin regions during induced HF cycling in adolescent C57BL/6 mice (Figure 3; Table 1):
Clusters of keratinocytes in the interfollicular epidermis (Figure 2A).
Peri-infundibular outer root sheath (piORS) keratinocytes (Figures 2B and 2C).
Keratinocytes of the anagen hair matrix, most pronounced in a unilateral cluster of hair matrix cells (Figure 2D).
Fibroblasts of the proximal connective tissue sheath (Figures 2E-2G).
Interfollicular dermal fibroblasts (Figures 2H and 2I).
Endothelial cells of the plexus profundus and of perifollicular blood vessels (Figure 2J).
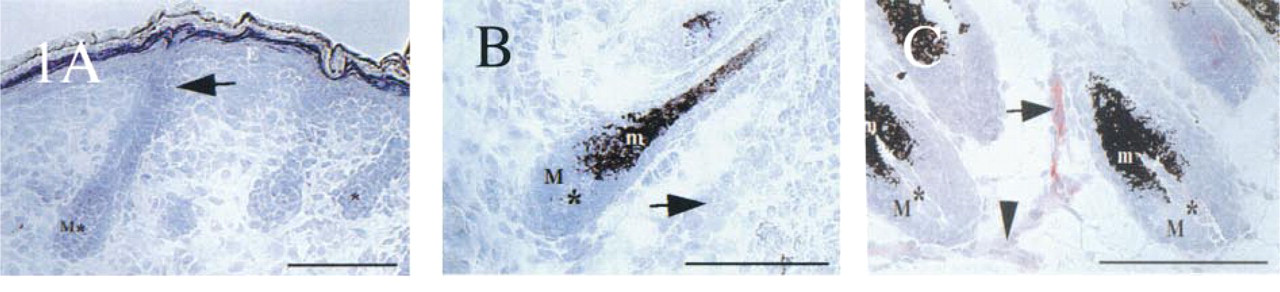
Expression patterns of ICAM-1 during neonatal HF morphogenesis in C57BL/6 mice. (
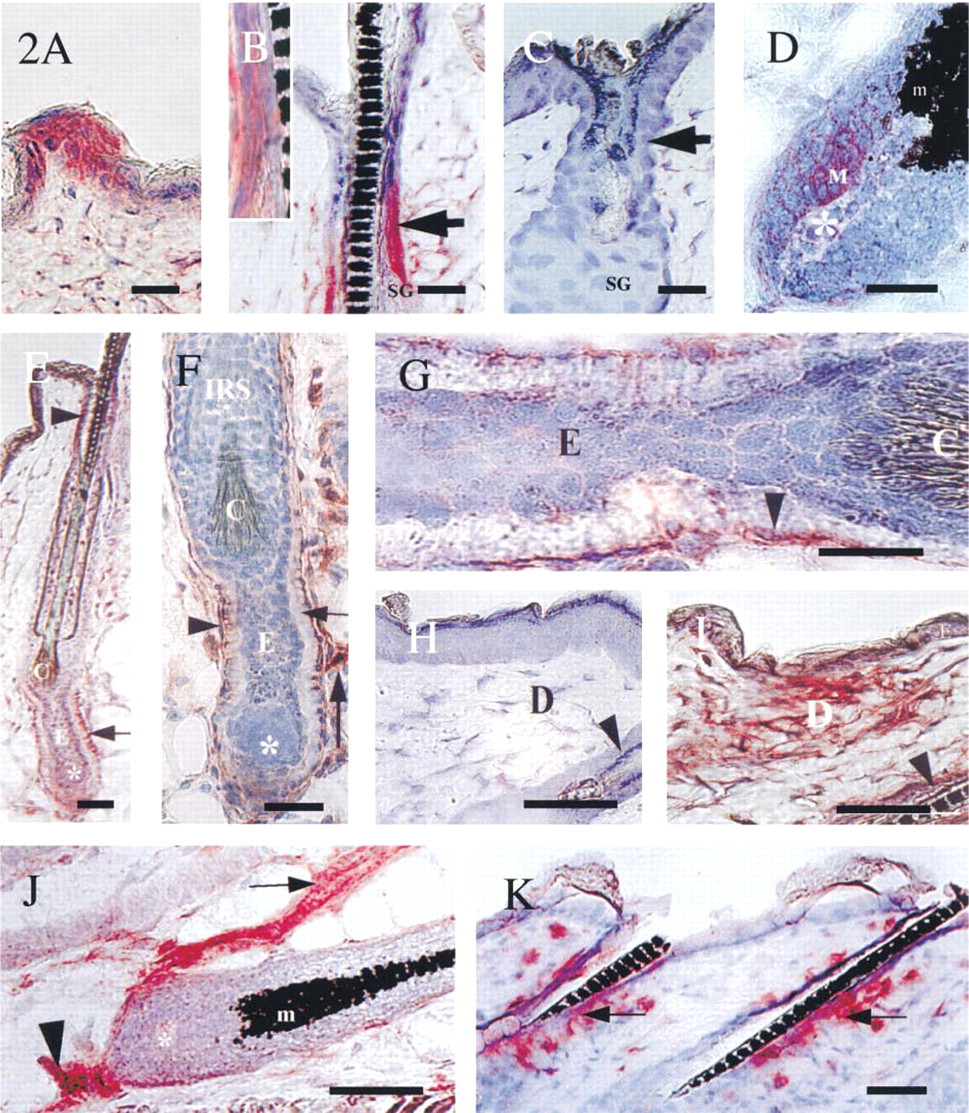
Expression patterns of ICAM-1 during the depilation-induced adolescent hair cycle in C57BL/6 mice. (
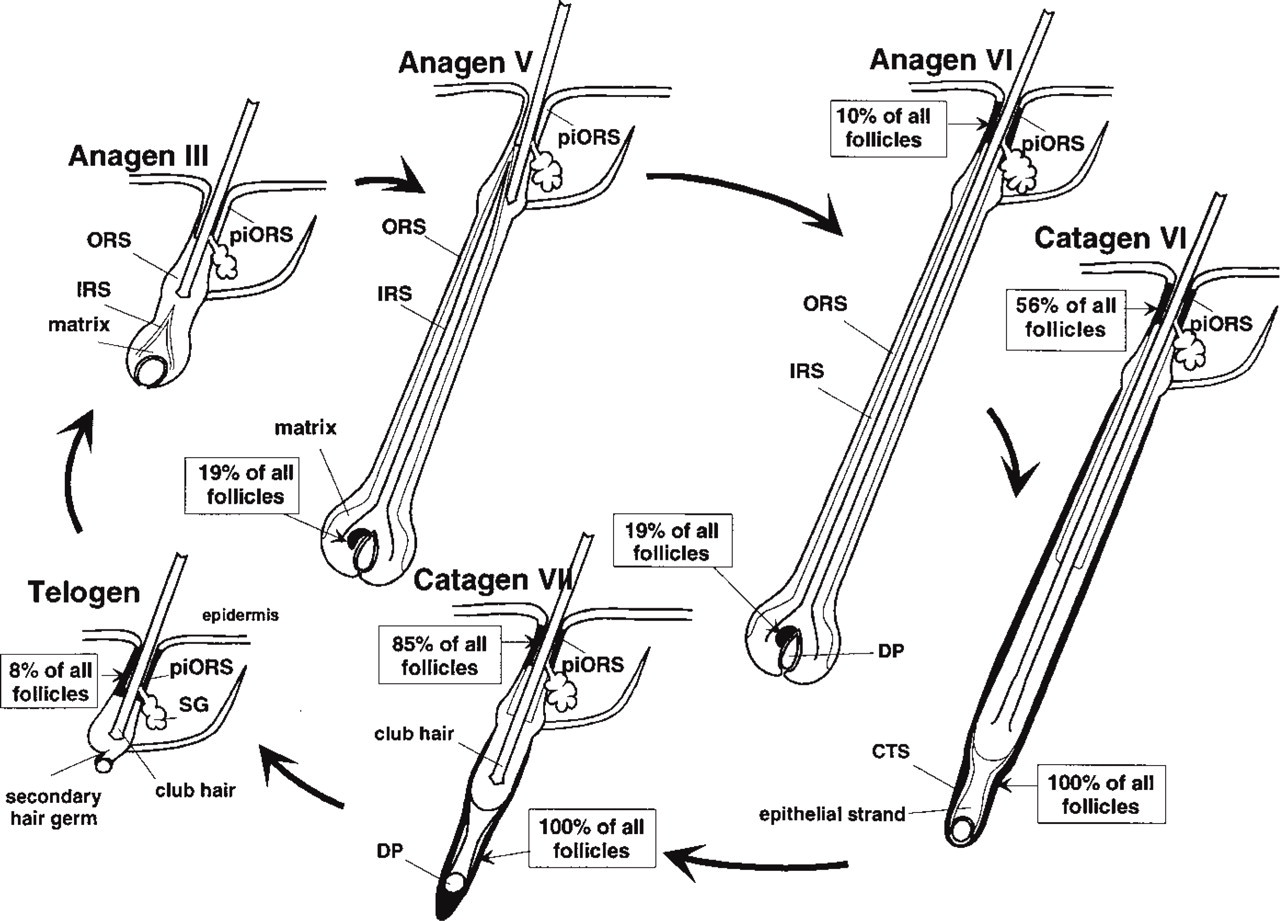
Schematic representation of ICAM-1 immunoreactivity during the depilation-induced mouse hair cycle (only strong ICAM-1 IR is represented). “Peri-infundibular outer root sheath” (piORS) refers to outer root sheath keratinocytes localized around the insertion of the infundibulum of the sebaceous gland. Where percentage values are listed, this indicates that only a part of the follicles showed this expression pattern, whereas the rest were ICAM-1 negative. DP, dermal papilla; CTS, connective tissue sheath; IRS, inner root sheath; ORS, outer root sheath; SG, sebaceous gland.
In these mouse skin regions, the following observations on ICAM-1 IR were made. First, Several sections harvested at Day 1 post depilation (p.d.) [i.e., skin sections with all HFs in the earliest stage of synchronized, induced anagen development (anagen I)] showed a general epidermal increase in ICAM-1 IR compared to telogen (see Table 1), most likely as a response to the slight skin wounding associated with anagen induction by depilation (Argyris 1967; Paus et al. 1990). This phenomenon was virtually absent on all sections harvested at Day 3 p.d, i.e., even early during anagen development (anagen II-III). Scattered, patch-like clusters of ICAM-1+ interfollicular epidermal keratinocytes (Figure 2A) were noted throughout the entire hair cycle. These ICAM-1+ epidermal cell patches displayed weaker staining intensity during early and middle anagen compared to an intermediate staining intensity during telogen, late anagen, and catagen (Table 1).
The peri-infundibular region of the distal outer root sheath (piORS) showed a well-circumscribed ICAM-1 IR (Figure 2B), which changed substantially during the hair cycle (Table 1; Figure 3). Telogen skin only rarely exhibited follicles with a strongly ICAM-1+ piORS (defined here as at least half of the staining intensity of untreated telogen skin blood vessels), i.e., in 8% of all telogen follicles. The average percentage of strongly ICAM-1+ follicles decreased to 0% after anagen induction by depilation (p<0.05). Skin sections of Day 1 (anagen I) and Day 3 p.d. (anagen II-III) showed only very faint ICAM-1 IR of the piORS, and no follicles at all were detected that displayed a strongly ICAM-1+ piORS. At Days 5 and 8 p.d. (anagen III-V), ICAM-1 IR of the piORS was completely absent (Figure 2C). Follicles with ICAM-1+ piORS appeared to be fairly regularly distributed throughout the back skin and occurred most frequently in groups of two or three follicles with a strongly ICAM-1+ piORS. Unexpectedly, even topical application of 100% DMSO for 3 days, which causes a marked irritant dermatitis, induced only a faint upregulation of ICAM-1 IR in the piORS of anagen and telogen follicles. This was in striking contrast to the expected dramatic increase in dermal fibroblast ICAM-1 IR after DMSO treatment (not shown).
Expression pattern of ICAM-1 in distinct anatomic compartments during the depilation-induced mouse hair cycle a
Only strong, highly reproducible IR patterns are represented, based on the analysis of at least 100 HFs per stage of HF cycling. Where percentage values are listed, this indicates that only a part of the follicles showed this expression pattern, while the rest was ICAM-1 negative. (+), very weak staining intensity; ++++, very strong staining intensity.
Compared to early and middle anagen (Days 1–8 p.d.), the percentage of follicles with a strongly ICAM-1+ piORS increased significantly during progressing later anagen and spontaneous HF entry into catagen (Figure 4A). In catagen VII-VIII, 85% of all follicles showed an ICAM-1+ piORS (Figures 2B, 3, and 4A), followed by the maximum at Day 20 p.d., when skin is composed of late catagen and the beginning of telogen development.
In line with our previous reports (Paus et al. 1994a,c, 1998; Rückert et al. 1998), sequential sections revealed co-localization of ICAM-1+ keratinocytes in the piORS with γδT-cells, nonclassical MHC Class I (MHC Ib) molecules (Qa-2a), and NLDC 145+ Langerhans cells in this region of the distal HF epithelium (not shown; for comparison see Paus et al. 1998; Rückert et al. 1998). Furthermore, the perifollicular mesenchyme of at least every fifth follicle displayed a dense infiltrate of LFA-1+ cells of macrophage phenotype at the level of the piORS (Figure 2K) throughout all hair cycle stages (cf. Eichmüller et al. 1998). Further differentiation by double staining (Eichmüller et al. 1996) revealed that the majority of these LFA-1+ cells were also MHC Class II+ and ER-MP 23+, but MAC-1− (not shown; see Eichmüller et al. 1998).
Strikingly asymmetric ICAM-1 expression was detected on hair matrix keratinocytes in 19% of all induced anagen IV-VI follicles, most pronouncedly in a unilateral cluster of ICAM-1+ hair matrix keratinocytes (Figure 2D, “M”).
Starting from catagen III, several of the most proximally located fibroblasts of the proximal connective tissue sheath became strongly ICAM-1+. During the subsequent stages of catagen development, from the dermal papilla to the insertion of the sebaceous gland, a continous layer of ICAM-1+ mesenchymal cells became detectable surrounding the HF basement membrane (Figures 2E-2G). The highest staining intensity was noted during catagen VI on cells of the proximal connective tissue sheath surrounding the epithelial strand of the regressing hair bulb (Figures 2E-2G). One layer of fibroblasts directly adjacent to the glassy membrane of the epithelial strand of the regressing follicle exhibited ICAM-1 IR (Figures 2F and 2G). On serial sections, these cells were strongly immunoreactive with the pan-reticular fibroblast marker ER-TR 7 but not with antibodies against E- or P-cadherin as markers of follicular keratinocytes (cf. Müller-Röver and Paus 1998) or antibodies against MHC class II or ER-MP-23 as macrophage markers (cf. Eichmüller et al. 1998; Paus et al. 1998) (not shown).
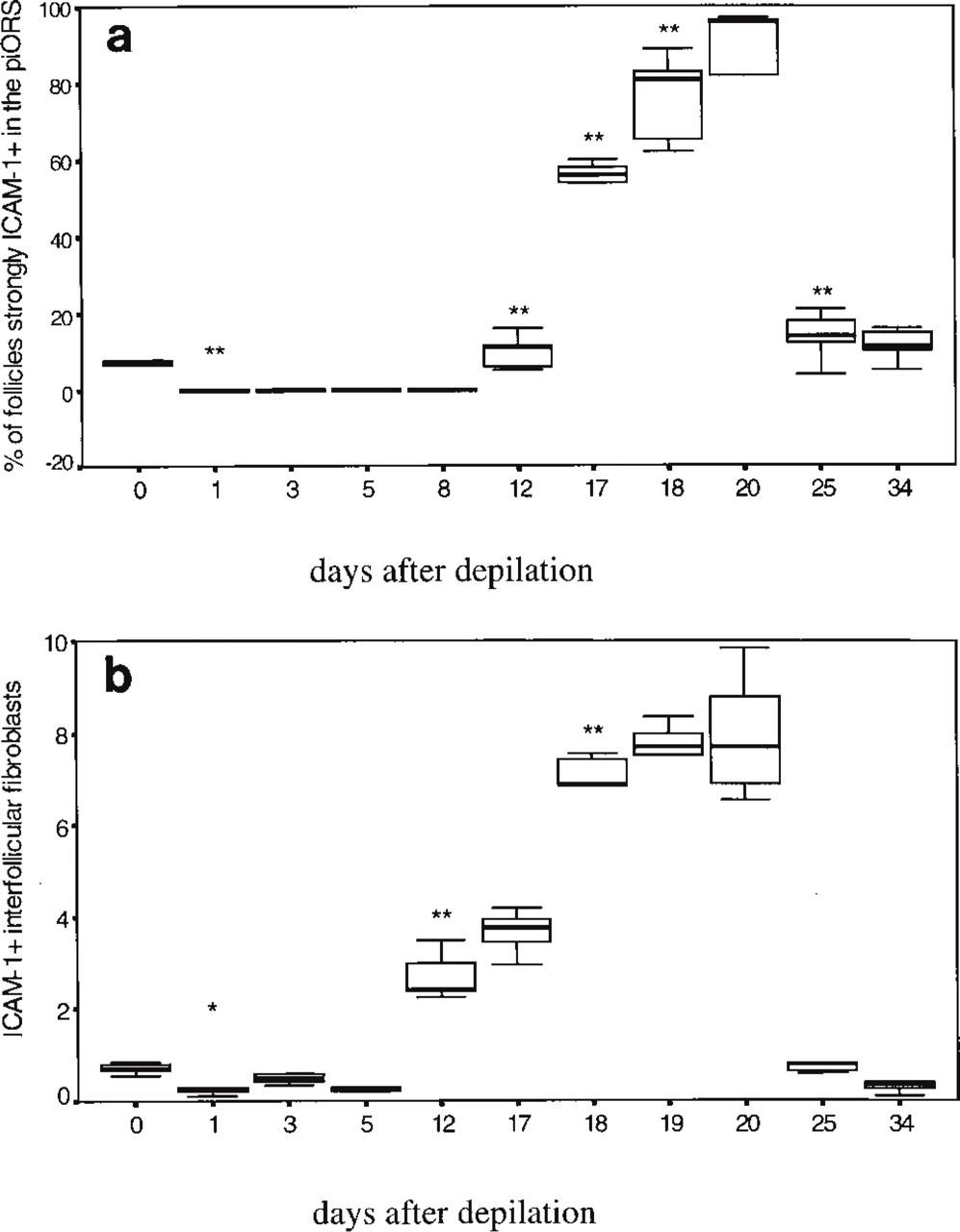
Percentage of follicles strongly ICAM-1+ in the periinfundibular region of the outer root sheath (piORS) at different days p.d. (defined as “at least half of the staining intensity of telogen skin blood vessels”). Five mice per hair cycle time point and 20 follicles per mouse were counted at X400 magnification. Box-and-whisker plots show median (large black bar), interquartile range (25%-ile to 75%-ile, box) and range (in between small bars), ∗∗, highly significant change compared to preceding hair cycle time point (p<0.01). (
Interfollicular dermal fibroblasts showed a highly dynamic hair cycle-dependent pattern of ICAM-1 IR (Figures 2H, 2I, and 4B). In the interfollicular dermis, a small but significant decline of strongly ICAM-1+ fibroblasts was noted shortly after anagen induction (Day 1 p.d.), followed by a strong, significant increase (p<0.01) during anagen VI and catagen development (Days 12–20 p.d.; cf. Figures 2I and 4B) and another sharp decline to again reach telogen values (Day 25 p.d.; telogen; Figure 4B). This substantial upregulation of constitutive ICAM-1 IR on interfollicular dermal cells during the anagen-catagen transformation was very similar to that seen in inflamed telogen skin after induction of an irritant contact dermatitis by DMSO treatment and was noted around all catagen follicles, irrespective of whether they had developed after depilation-induced or spontaneous anagen.
On endothelial cells of the plexus profundus, strong constitutive ICAM-1 IR was noted in all hair cycle stages (Figure 2J; Table 1). A gradually increasing number of perifollicular blood vessels displayed strong ICAM-1 IR during middle and late anagen (Figure 2J; Table 1). This was reminiscent of the upregulation of ICAM-1 IR observed on perifollicular blood vessels during the later stages of HF morphogenesis (Figure 1C), yet much stronger.
Control sections of 10-week-old mice that had developed anagen V and VI spontaneously displayed the same ICAM-1 IR patterns as seen during depilation-induced anagen V and VI. Again, the strongest cutaneous anti-ICAM-1 staining intensity was found on the endothelial cells of the plexus profundus and on perifollicular blood vessels, while the strongest follicular ICAM-1 IR was detected on piORS keratinocytes. In addition, the ICAM-1 IR patterns in the hair matrix and interfollicular epidermis, as well as those on interfollicular dermal cells and dermal papilla fibroblasts, were the same during spontanous anagen as described and documented above for depilation-induced anagen. This excludes the possibility that the observed changes in ICAM-1 IR merely reflect wound healing-related artifacts.
LFA-1 Expression Is Hair Cycle-dependent and Is Apparently Upregulated on HF-associated Immune Cells
Throughout the depilation-induced hair cycle, LFA-1+ cells were detected in the epidermis, in and around the piORS, and scattered in the interfollicular dermis. The highest cutaneous density of LFA-1+ macrophages was found in the upper dermis around HFs with strongly ICAM-1+ piORS (Figure 2K).
The anti-LFA-1 staining intensity and the number of LFA-1+ cells exhibited substantial hair cycle-dependent fluctuations. Immediately after anagen induction by depilation, the LFA-1 IR, as well as the number of dermal LFA-1+ macrophage-like cells and the number of intraepithelial LFA-1+ Langerhans cells, declined substantially compared to telogen (Day 0). During middle anagen (Day 8 p.d.), a strong increase in LFA-1 IR and a numerical increase of dermal LFA-1+ macrophage-like cells were noted.
During late anagen and the anagen-catagen transition (Days 12–18 p.d.), LFA-1 IR was seen on macrophage-like cells that accumulated around and inside the piORS (Figure 2K). These intraepithelial LFA-1+ cells in the piORS co-localized with markers for intrafollicular γδT-cells and Langerhans cells (not shown; cf. Eichmüller et al. 1998; Paus et al. 1994a,c, 1998). Compared to anagen and catagen, a lower anti-LFA-1 staining intensity and a decline in the number of LFA-1+ macrophage-like cells around the piORS was noted during telogen (Days 0 and 25 p.d.). Virtually no LFA-1 IR was seen in the piORS in telogen.
ICAM-1 and LFA-1 mRNA Expression
ICAM-1 and LFA-1 mRNA steady-state levels in full-thickness back skin homogenates of adolescent C57BL/6 mice were assessed by semiquantitative RT-PCR (Figure 5). This revealed a significant decline of ICAM-1 transcript levels early after depilation-induced anagen development (Days 1–3 p.d.) compared to telogen (p<0.01) and a significant upregulation during middle anagen (Day 8 p.d) (p<0.05). During the subsequent anagen-catagen-telogen transition, ICAM-1 transcript levels again declined significantly (p<0.01). These hair cycle-associated fluctuations in ICAM-1 transcript levels corresponded well to the hair cycle-related changes noted in ICAM-1 antigen expression (Figure 4; Table 1). Interestingly, almost the same fluctuations of LFA-1 mRNA steady-state levels were found as for ICAM-1. A significant decline of the LFA-1 transcript levels was seen immediately after anagen induction (p<0.01), a significant increase on Day 8 p.d. (p<0.05), and again a significant decline during the anagen VI-catagen-telogen transition (p<0.01) (Figure 5D). Thus, the steady-state levels for both ICAM-1 and LFA-1 mRNA peaked in mouse skin with almost all HFs in anagen VI or catagen.
ICAM-1-deficient Mice Show Catagen Acceleration
To further probe the concept that ICAM-1 expression is functionally important for HF regression, the first spontaneous postnatal catagen development was compared by quantitative histomorphometry between ICAM-1 knockout mice and age-matched wild-type littermates. Icamtm1Bay knockout mice displayed a statistically significantly accelerated HF regression compared to wild-type controls (Figure 6). At Day 18 post partum, a significantly higher percentage of HFs in wild-type mice were still in early catagen (p<0.02) compared to HFs in ICAM-1 knockout mice, and vice versa, a significantly higher percentage of HFs in ICAM-1 knockout mice were already in late catagen (p<0.05) compared to HFs in wild-type littermates.
Discussion
In this study we demonstrate that mouse skin and HFs exhibit a more widespread and much more dynamic constitutive ICAM-1 expression pattern (Figures 1–3) than was previously recognized (Goebeler et al. 1990, 1994; Scheynius et al. 1993; Williams and Kupper 1994). In particular, novel physiological and hair cycle-dependent patterns of ICAM-1 expression are identified in defined anatomic skin compartments (Table 1): on selected epidermal keratinocytes (Figure 2A), in the peri-infundibular outer root sheath (piORS) (Figures 2B and 2C), in the hair matrix (Figure 2D), and in the proximal connective tissue sheath (Figures 2E-2G) as well as on perifollicular endothelial cells (Figure 2J).
Four observations suggest a functionally important role for ICAM-1-related signaling in the control of HF regression. Increasingly strong ICAM-1 expression during skin and HF morphogenesis can be found only when the HFs enter into late neonatal anagen (Figures 1A-1C) and their first catagen stage. ICAM-1 expression in adolescent mouse skin peaks during the middle and late stages of synchronized HF regression (catagen), at both the protein (Figures 2B, 2E-2G, 2I, 3, and 4; Table 1) and the mRNA level (Figure 5). Finally, ICAM-1-deficient mice display accelerated catagen development compared to wild-type controls (Figure 6). This makes ICAM-1 the first adhesion molecule to be implicated in catagen control. Previously, only secreted cytokines, growth factors and hormones (cf. Stenn et al. 1996; Paus et al. 1996, 1997; Schilli et al. 1997; Cotsarelis and Paus 1998) as well as macrophages (Parakkal 1969; Westgate et al. 1991; Paus and Czarnetzki 1992) and mast cells (Maurer et al. 1997a; Paus et al. 1994d) have been considered as elements of catagen control.
Despite identical morphological appearance, neighboring mouse pelage follicles of identical cycle stage differ in their ICAM-1 expression pattern. Furthermore, each hair cycle stage is characterized by a changing percentage of selected, rather regularly distributed follicle populations that display a defined ICAM-1 expression pattern (Figures 1–3; Table 1). The ICAM-1 expression of cells is widely accepted to be under the control of their local cytokine milieu, with practically all proinflammatory cytokines upregulating ICAM-1 expression (Springer 1990; Kunkel et al. 1996; van de Stolpe and van der Saag 1996; Bos 1997). The very heterogeneous yet tightly regulated follicular ICAM-1 expression patterns revealed in the current study suggest that the peri- and intrafollicular cytokine milieu of HFs of the same type and cycle stage is much less uniform than has been appreciated thus far. The challenge now is to characterize, first, how morphologically indistinguishable follicles with distinct ICAM-1 expression patterns differ in their local cytokine milieu and, second, to define the relevance of any such differences for the control of synchronized HF cycling and for other HF functions.
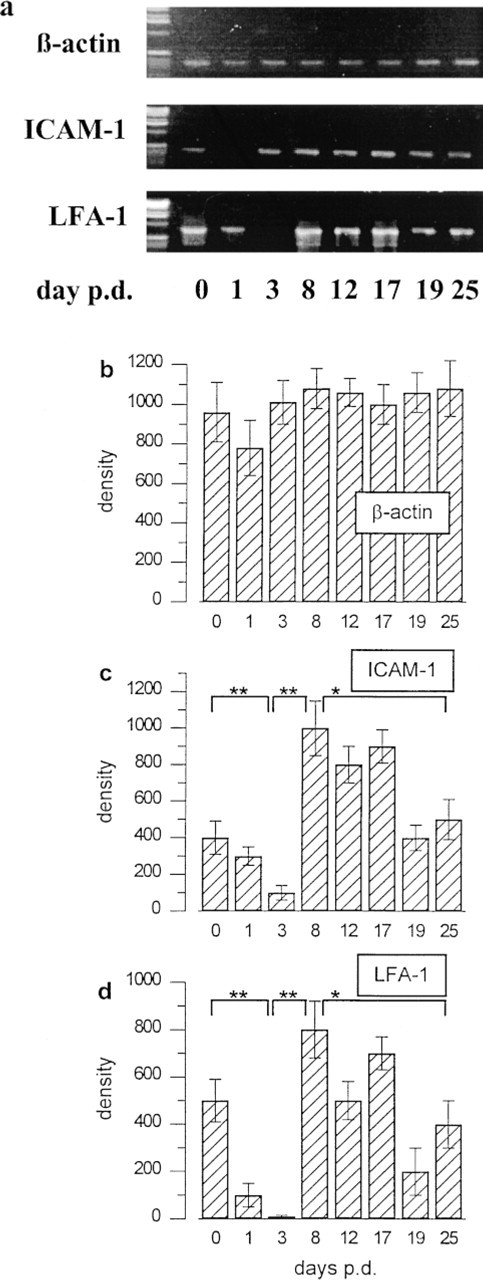
mRNA steady-state levels for ICAM-1 and LFA-1 during the depilation-induced mouse hair cycle. (
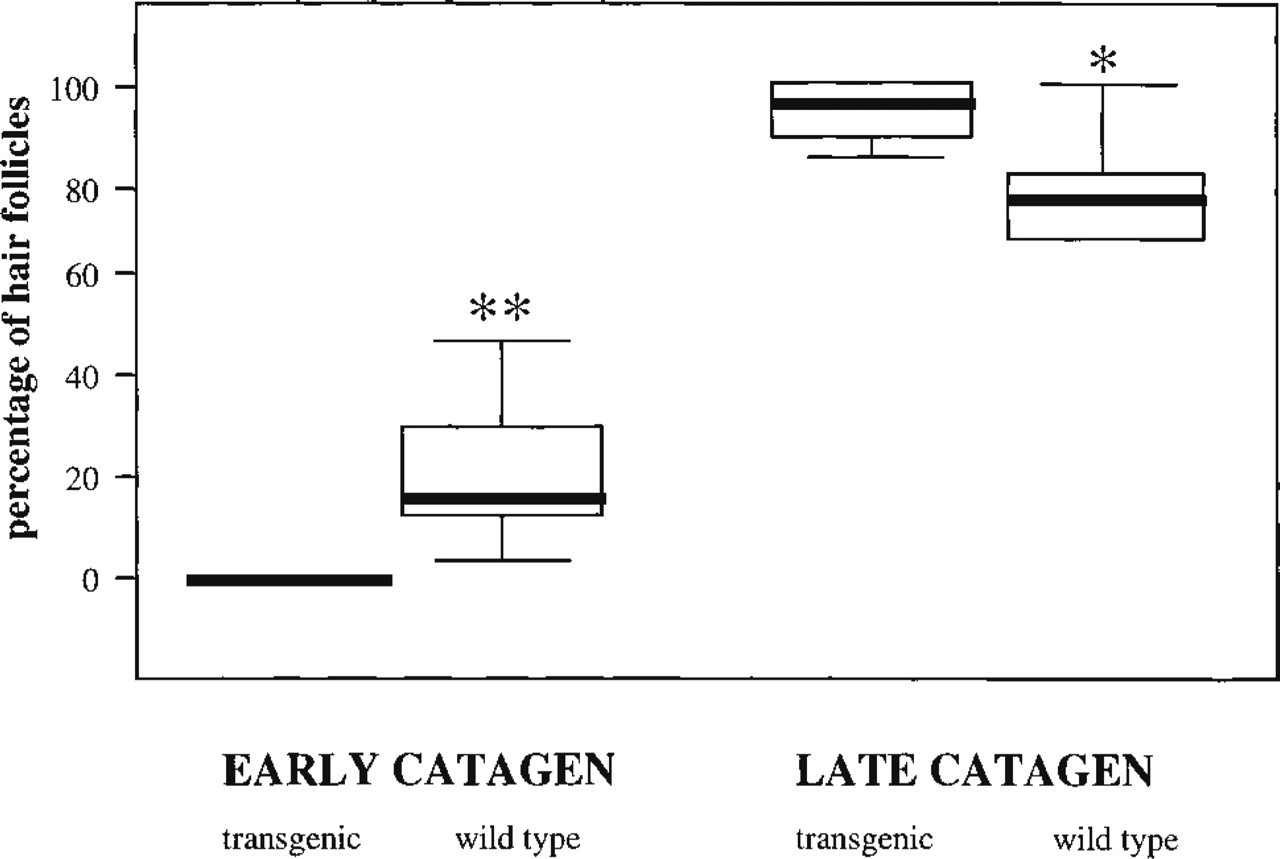
A significant acceleration of spontaneous catagen development at Day 18 post partum was found in ICAM-1-deficient mice. The percentage of hair follicles in early and late stages of catagen was assessed histomorphometrically for 700 HFs per group (100 HFs per mouse). ∗/∗∗Significant difference from transgenic mice, ∗p,0.05; ∗∗p<0.02; values are means ± SEM.
ICAM-1 expression on endothelial cells (Figure 2J) and piORS keratinocytes (Figure 2B) may primarily serve immunological functions. Vascular ICAM-1 expression is essential for leukocyte recruitment into inflammatory sites (cf. Springer 1994; van de Stolpe and van der Saag 1996). Therefore, changes in the constitutive vascular ICAM-1 expression may be one tool for mediating the as yet ill-explained but striking hair cycle-associated changes in the skin immune system and its responses (Hofmann et al. 1996; Paus 1997; Tokura et al. 1997; Cotsarelis and Paus 1998; Paus et al. 1998). For example, advanced anagen development may induce an intracutaneous cytokine milieu that upregulates endothelial ICAM-1 expression, leading to increased physiological extravasation and accumulation of immune cells in anagen and/or catagen skin. ICAM-1 expression of the piORS during late anagen VI, catagen, and telogen (Figures 2B, 2E, and 3) may serve to accumulate LFA-1+ macrophage-like cells around this level of the HF (Figure 2K), which is the major port of entry for microbial invasion of the skin and a preferred route of entry for noxious chemical agents (Paus 1997). Because ICAM-1 expression by keratinocytes is believed to be an essential element in initiating cutaneous inflammatory responses (Barker et al. 1991; Nickoloff and Griffiths 1991; Bos 1997), environmental stimuli, such as dense colonization of the follicle canal by various microorganisms, may continuously induce the piORS epithelium to elaborate cytokines that upregulate ICAM-1 (e.g., TNFα or IFN-γ). This, in turn, may attract dendritic epidermal T-cells and Langerhans cells to accumulate in this region (Paus et al. 1994c; Cotsarelis and Paus 1998) to generate an immunological microenvironment that is conducive to effective infection defense. The transient downregulation of ICAM-1 in the piORS observed during early anagen may reflect the generally immunosuppressive skin environment previously documented for synchronized early anagen (Hofmann et al. 1996; Paus 1997; Tokura et al. 1997).
Originally, ICAM-1 was mainly viewed as a simple “glue” for β2-integrin binding. This view has been changed since the recognition that ICAM-1 also serves intracellular signaling functions. For example, antibody cross-linking of ICAM-1 can induce activation of the transcription factor AP-1 as well as IL-1 β transcription (Koyama et al. 1996), can upregulate MHC II expression, can stimulate tyrosine phosphorylation of the Src family kinase p53/p56, and can activate Raf-1 and mitogen-activated protein kinase (Holland and Owens 1997). Adhesion molecules such as ICAM-1 therefore exert profound “topobiological” signaling functions (Edelman 1988, 1992; Müller-Röver and Paus 1998). In the case of ICAM-1, such morphoregulatory and classical immunoregulatory functions may converge during catagen. Note that HF regression is characterized by precisely choreographed, apoptosis-driven involution of the HF epithelium (Straile et al. 1961; Lindner et al. 1997) and by extensive matrix remodeling in which perifollicular macrophages and mast cells have been implicated (Parakkal 1969; Westgate et al. 1991; Paus and Czarnetzki 1992; Paus 1996; Stenn et al. 1996; Maurer et al. 1997a).
The precise cause of accelerated catagen development in ICAM-1 -/- mice remains subject to further investigations. Although ICAM-1 deficiency may modulate the extravasation of blood immune cells into the skin, we found no substantial alteration in the number of MAC-1+, LFA-1+, and MHC-II+ immune cells in key anatomic compartments (especially around the regressing epithelial strand) of ICAM-1 -/- mice compared to wild-type controls (not shown). This suggests that catagen acceleration in ICAM-1 -/- mice might rather be due to a systemic alteration of the skin and hair follicle immune system in these mutants, e.g., ICAM-1 deficiency of perifollicular immune cells or fibroblasts may modulate their cytokine release, leading to a catagen-promoting cytokine milieu.
In summary, this study demonstrates that the transient upregulation of cell adhesion molecule expression, such as ICAM-1 and LFA-1, is not limited to pathological conditions (e.g., during inflammation and infection) and that it may also reflect an as yet underinvestigated involvement of ICAM-1 in physiological tissue remodeling processes such as HF involution. Further studies on the role of ICAM-1-related signaling in catagen control may provide one of the missing links between developmental and immunological aspects of hair growth control.
Footnotes
Acknowledgements
Supported in part by grants from the European Union under the Industrial & Materials Technologies Programme (Brite-Euram III: BE97-4301) (to IAM, RP) and Wella AG, Darmstadt (to RP), by a grant from Boehringer Ingelheim Fonds to SMR, and by a grant from the National Cancer Institute (CA 34196) (to JPS).