
Abstract
The amino acid taurine has been implicated in several aspects of reproductive system physiology. However, its localization in these organs has not been previously analyzed. The aim of this study was to characterize its distribution in male rat reproductive organs by immunohistochemical methods. Taurine was localized in the smooth muscle cells of the tissues studied and in the skeletal fibers of the cremaster muscle. In the testis, taurine was found in Leydig cells, vascular endothelial cells, and other interstitial cells. No immunoreactivity was observed in the cells of the seminiferous tubules, either in germ cells at all spermatogenic stages or in Sertoli cells. However, peritubular myoid cells were immunostained. Most epithelial cells of the efferent ducts were immunolabeled, whereas the epithelial cells of the rete testis (extratesticular segments), epididymis (caput, corpus, and cauda regions), and ductus deferens were unstained. However, most epithelial cells from the intratesticular segments of the rete were immunopositive. Some cells identified as intraepithelial macrophages and lymphocytes, apical cells, and narrow cells were intensely immunolabeled. Regional differences in the distribution of these cell types along the ducts studied were also noted. The possible functional roles for taurine in these cells are discussed.
T
In the male reproductive system, taurine has been identified as the major free amino acid of sperm cells (van der Horst and Grooten 1966; Hernvann et al. 1986; Holmes et al. 1992a, b). Taurine may act as an antioxidant in preventing sperm lipid peroxidation (Alvarez and Storey 1983), as a capacitating agent (Meizel et al. 1980; Meizel 1985), and as a sperm motility factor (Fraser 1986; Boatman et al. 1990). It has been suggested that taurine inhibits plasma membrane Na+,K+-ATPase in membrane homogenates from epididymal hamster sperm cells (Mrsny and Meizel 1985). Taurine could also modify sperm phospholipid methyltransferase activity (LLanos and Ronco 1994). In spite of these important actions of taurine in the reproductive system, its localization in these tissues has not been previously studied. The immunohistochemical localization of taurine in the reproductive organs could contribute to the understanding of these and other functional roles with a different perspective beyond the scope of molecular biology and biochemical methods. The aim of this study was to determine the distribution of taurine in the testis and the convoluted ducts of the rat male reproductive system using immunohistochemical methods.
Materials and Methods
Albino Wistar rats (250 g) were used for the present study. Animals were anesthetized with a ketamine (0.08 ml/100 g), xylocaine (0.04 ml/100 g), and atropine (0.02 ml/animal) mixture and perfused through the ascending aorta with 100 ml phosphate buffer (0.12 M, pH 7.4), followed by the fixative solutions. The compositions of the fixatives used were 4% paraformaldehyde, 0.5%, 1%, or 1.5% glutaraldehyde, and 4% paraformaldehyde (control specimens). Tissue samples were removed from the rats and stored in the same fixative solutions used during perfusion for 6-24 hr, washed in buffer, dehydrated, and embedded in paraffin according to conventional methods (González-Santander et al. 1997).
Streptavidin-Peroxidase Method
Serial sections of the paraffin-embedded samples were obtained and the immunolabeling was performed in one section each 30 μm. Sections (5 μm thick) were mounted on silanized slides and allowed to dry overnight before immunohistochemical staining. After xylene deparaffinization, sections were hydrated and incubated for 20 min in 0.3% H2O2 followed by 0.03% H2O2 in methanol, washed in Tris-buffered saline (TBS), pH 7.5, and incubated for 10 min in a 0.1 M sodium borohydride solution in TBS, pH 7.5. Sections were then washed and incubated in 3% normal goat serum with 0.05% Triton X-100 in TBS, pH 7.5, at room temperature (RT) for 30 min, followed by overnight incubation at 4C with the rabbit anti-taurine antibody (Chemicon International; Temecula, CA) diluted from 1:500 to 1:1500 in the same buffer. Afterwards the sections were washed twice in TBS to remove unbound primary antibody and then incubated with the secondary antibody, biotinylated goat anti-rabbit IgG (Zymed Labs; San Francisco CA), for 1 hr at RT, and washed in TBS. Sections were incubated with the streptavidin-peroxidase complex (Zymed) for 10-30 min and washed in TBS, followed by Tris-HCl buffer, pH 7.6. The peroxidase activity was revealed using 3′-diaminobenzidine tetrahydrochloride (DAB) as chromogen (Sigma; St Louis, MO). The reaction product of DAB was intensified with nickel nitrate to obtain a dark black color of immunostained antigens [10 μl of nickel nitrate (8%) in 1 ml DAB-H2O2 solution]. Thereafter, the sections were dehydrated in ethanol and mounted in Eukitt (O. Kindler; Freiburg Germany) without being counterstained. To estimate the percentage of immunostained cells, 20 areas for each tissue region were chosen at random and at least 50 cells of each type were counted in each area. Only those cells with definitely recognizable nuclei were counted. Values obtained for each cell type were expressed as mean ± SD.
Control Experiments
The specificity of the immunohistochemical procedures was assessed by means of the negative controls performed: (a) omitting the primary antibody; (b) using nonimmune serum instead of the primary antibody; (c) incubating with an inappropriate secondary antibody after the incubation with the primary antibody at optimal titers; and (d) immunolabeling on 4% paraformaldehyde-fixed samples. The last control should not give a positive immunoreaction because the antitaurine antibody used here was prepared using taurine conjugated by glutaraldehyde to poly-L-lysine as immunogen, and therefore it recognizes only haptenic taurine on glutaraldehyde-fixed samples (Campistron et al. 1986). As positive controls, cerebellar sections from the same rats were immunostained simultaneously with sections from the reproductive organs. Moreover, the anti-taurine antibody used here has been previously utilized and extensively characterized to confirm its specificity by ELISA, showing that it virtually does not crossreact with other related molecules such as hypotaurine or GABA (Campistron et al. 1986).
Results
The distribution of taurine in the testis (Figures 1a-1g), rete testis (Figures 1h and efferent ducts (Figures 1j and 1k), epididymis (Figure 2), and ductus deferens (Figures 3a and 3b) of the rat was studied. With the different paraformaldehyde-glutaraldehyde solutions used, no differences in the pattern of taurine distribution were observed (Figures 1a-1g). No staining was found in the negative controls performed (not shown). Positive controls (Figure 3k) showed a taurine distribution in Purkinje cells of rat cerebellar sections similar to that reported in other studies (Campistron et al. 1986; Nagelhus et al. 1993). Moreover, an internal positive control for taurine immunostaining consisted of the smooth and skeletal muscles from the organs studied, in which taurine was detected as previously described in cat muscles (Quesada et al. 1992, 1993). The smooth muscle cells located beneath the basement membrane of the epithelium from the ducts studied were intensely immunostained (Figures 3a and 3b). The skeletal fibers of the cremaster muscle showed wide differences in taurine immunoreactivity among individual cells; some fibers were strongly immunoreactive, whereas others were weakly stained or unstained (Figure 3c).
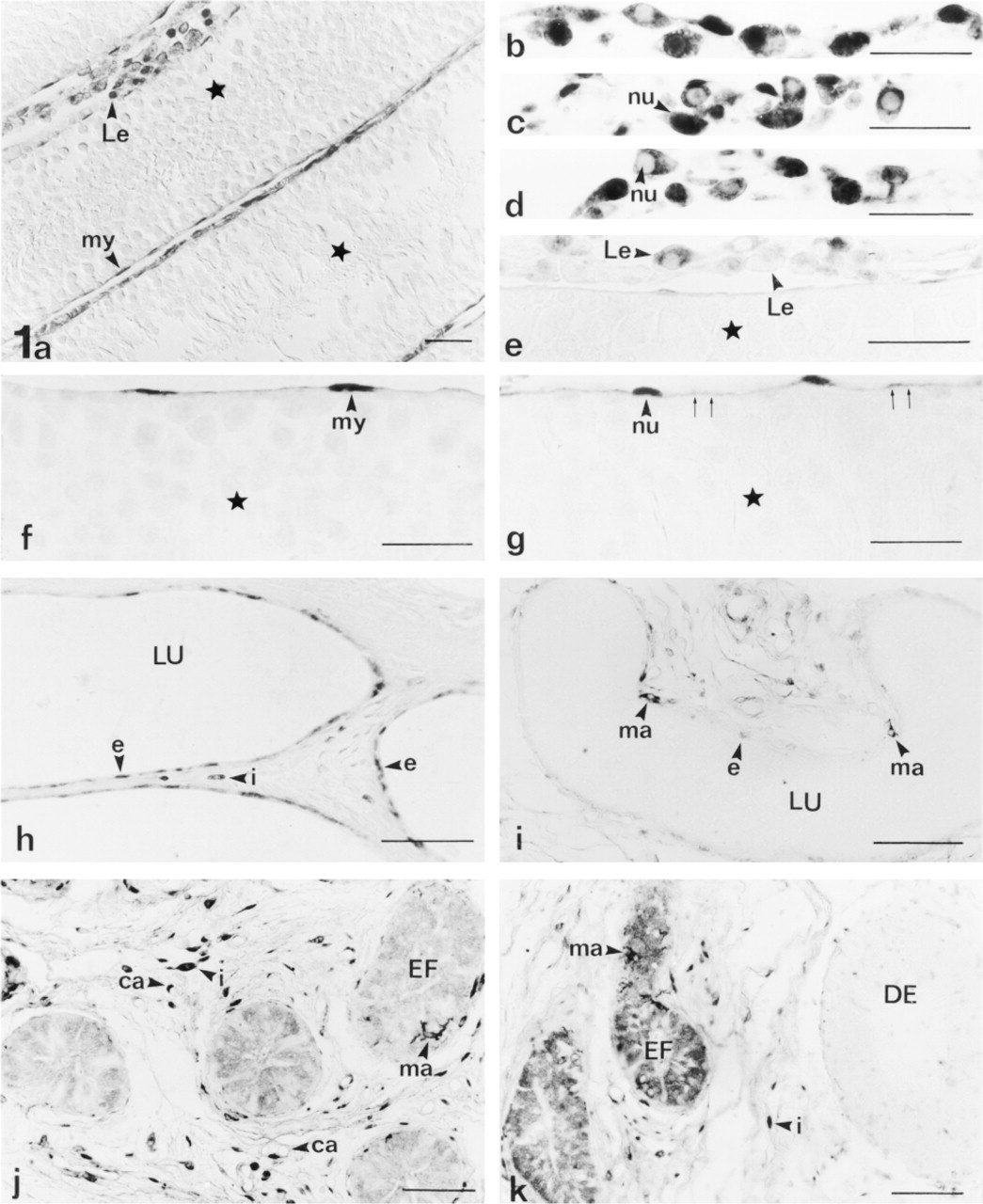
(a-g) Rat testis showing staining for taurine in the nucleus (nu) and cytoplasm (small arrows) of peritubular myoid cells (my) and Leydig cells (Le). All cells from the seminiferous tubules (stars) are immunonegative. Note the variable degree of staining in Leydig cells. (
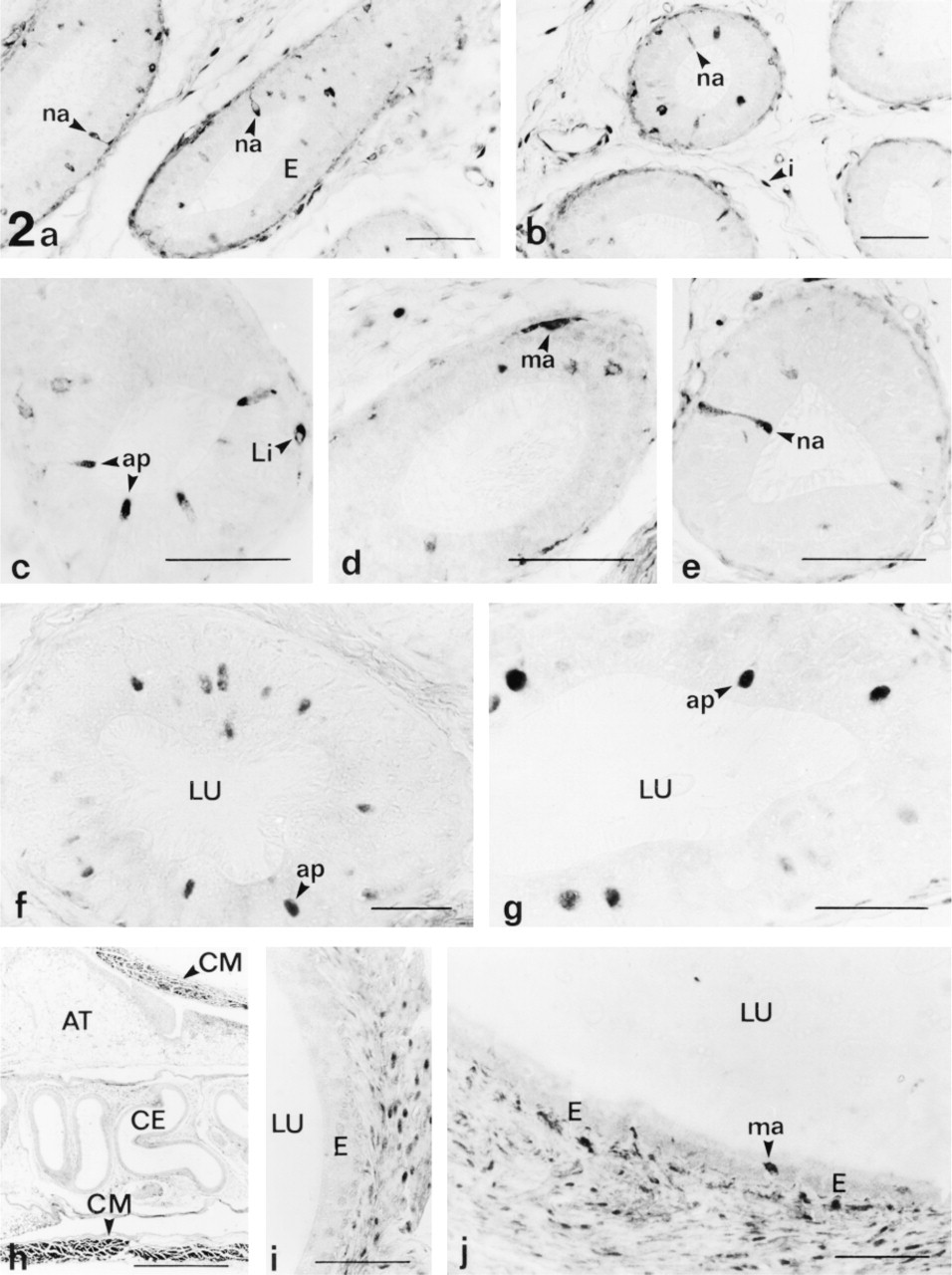
Taurine localization in the caput (
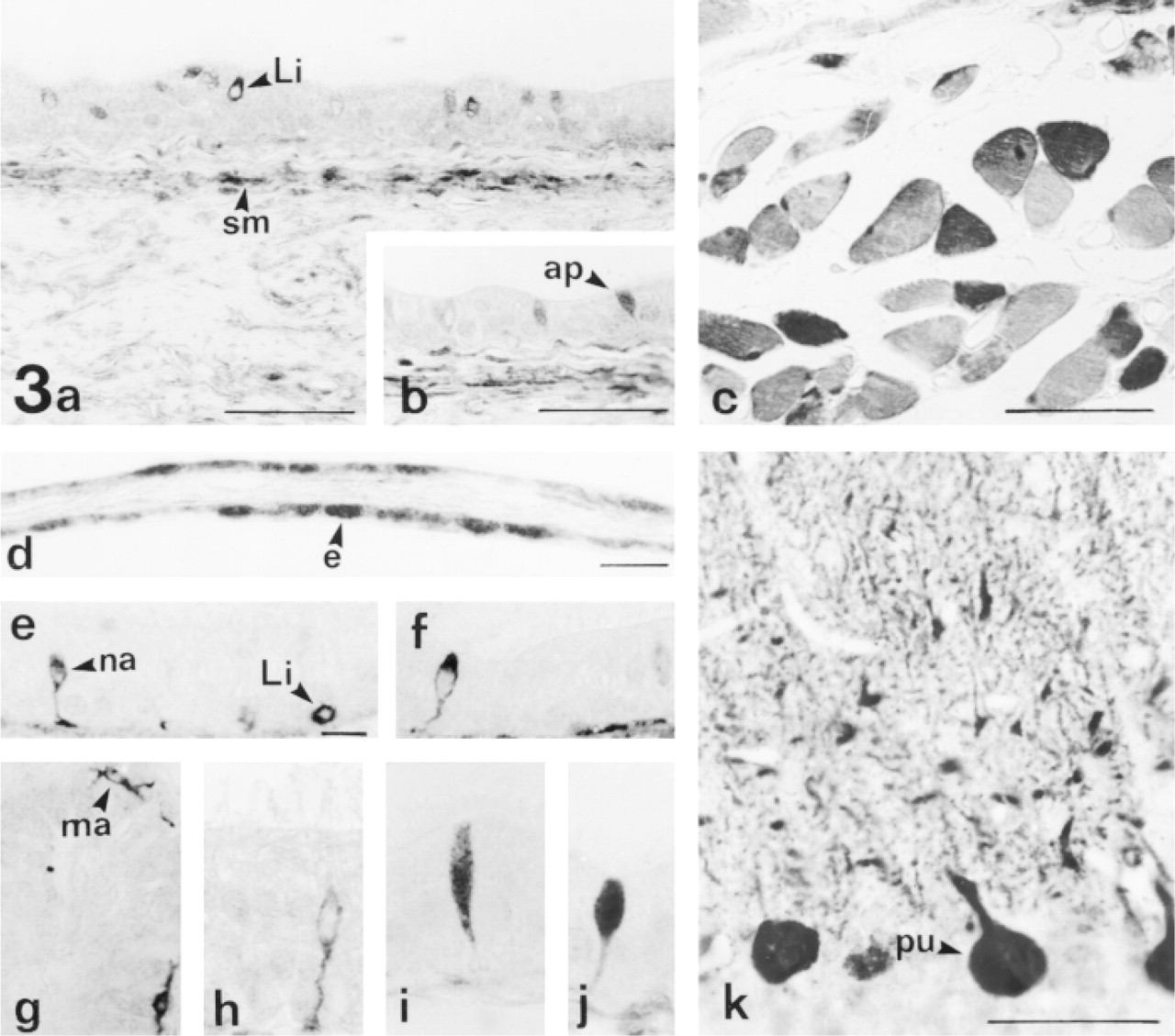
Rat ductus deferens showing taurine staining in smooth muscle cells (sm) and in some epithelial cells: apical cells (ap) and intraepithelial lymphocytes (Li). (
In the testis (Figures 1a-1g) taurine was detected in Leydig cells, vascular endothelial cells, and other interstitial cells. Most Leydig cells were stained (80.7 ± 4.1% of all Leydig cells). Leydig cells showed a variable degree of taurine immunostaining in their nuclei, being weak in some cells and as strong or stronger than in the cytoplasm in other cells. Moreover, taurine was homogeneously distributed in the cytoplasm of some Leydig cells, whereas in other cells it appeared to be located at discrete foci of the cytoplasm (Figures 1b-1e). In the seminiferous tubules, germ cells at all spermatogenic stages and Sertoli cells were unstained. However, peritubular myoid cells showed cytoplasmic immunolabeling along their thin cell processes that surround the seminiferous tubules, and a nuclear pool of taurine (Figures 1a, 1f, and 1g). In the tunica albuginea, some fibroblasts and/or myoid cells were stained.
In the rete testis, most epithelial cells of the initial segments (tubuli recti and tunical part) were immunopositive for taurine (Figures 1h and 3d). However, the epithelial cells from the extratesticular segment of the rete were immunonegative (Figure 1i). Moreover, taurine was also detected in some cells and blood vessels of the interstitial tissue located between the different segments of the rete testis (Figure 1h).
Most epithelial cells of the efferent ducts (Figures 1j and 1k) were immunostained (86.2 ± 9.1% of all epithelial cells). Frequently, taurine appeared homogeneously distributed in the nucleus and cytoplasm of these cells. However, a minor population of these epithelial cells showed intensely stained cytoplasm and weakly stained nuclei. Moreover, some cells scattered in the efferent ducts, which showed many cytoplasmic processes, were more intensely immunolabeled than adjacent epithelial cells. These cells were identified by their size, morphology, and distribution as intraepithelial macrophages (Figures 1j, 1k, and 3g). These macrophages were also intensely stained in other ducts, such as the rete testis (Figure 1i) and the epididymis (Figures 2d and 2j). In this cell type, taurine appeared mainly distributed in the perinuclear cytoplasm and in thin cytoplasmic processes, whereas the nucleus was usually weakly stained.
In the pseudostratified columnar epithelium of the epididymis, both main cell types, principal and basal cells, were immunonegative (Figure 2). Small round cells, which appeared isolated in the epididymal epithelium (probably intraepithelial lymphocytes), showed intense immunolabeling (Figures 2c and 3e). These lymphocyte-like cells were found at specific segments of the epididymis, being scattered all along the length of the caput and corpus, whereas they were not localized in the cauda epididymidis.
Taurine was found in some scattered epididymal cells, identified by the position of their nuclei in the upper half of the cytoplasm. This minor population of epididymal epithelial cells have been termed apical mitochondria-rich cells (Brown and Montesano 1980; Palacios et al. 1991), narrow cells (Sun and Flickinger 1980), holocrine cells (Martan and Risley 1963, 1964), and flask cells (Burkett et al. 1987). This cell type has been recently differentiated into two cell populations, apical and narrow cells, by their morphology, distribution, and immunostaining properties to several antibodies, although both show their nuclei in the upper half of the cytoplasm (Adamali and Hermo 1996). At the light microscopic level, apical cells are goblet-shaped cells with long or round nuclei, and narrow cells are slender elongated cells with flattened nuclei (Adamali and Hermo 1996; Fraile et al. 1996). Immunostained cells of both morphological types were observed. Narrow cells (Figures 2a, 2b, 2e, 3e, 3f, and 3h) were present only in the initial epididymal segments (caput epididymidis). In narrow cells, taurine was mainly found in their supranuclear region and alongside the nuclei, whereas their nuclei were usually weakly stained (Figures 3e, 3f, and 3h). They showed a thin cytoplasmic process extending towards the basement membrane that occasionally terminated in a peduncle. This infranuclear process (including the peduncle) was also immunostained (Figures 3e, 3f, and 3h). Apical cells were found in the caput (Figure 2c) and corpus epididymidis (Figures 2f, 2g, and 3i) and in the ductus deferens (Figures 3b and 3j). They were not found in the cauda epididymidis. In apical cells, taurine was usually found in the supranuclear cytoplasm and in the nucleus.
In the ductus deferens, most epithelial cells were unstained. However, taurine appeared to be distributed in some scattered cells that were identified as intraepithelial lymphocytes (Figure 3a) and apical cells (Figures 3b and 3j).
In the interstitial region among the different ducts studied, vascular endothelial cells and other interstitial cells (fibroblasts and/or periductal myoid cells) were also stained (Figures 1j, 1k, and 2). Moreover, a variable degree of taurine immunostaining in the endothelial cells of the capillaries was observed. Some capillaries were intensely immunoreactive, whereas others were not stained (Figure 1j).
Discussion
Mammalian spermatozoa are immature when they leave the testis. Their development continues during their passage through the epididymis, where they acquire fertilizing ability and a mature motility pattern. This maturation process appears to be inherent to the sperm cells as they age and is also influenced by the epididymal epithelium, which modifies the luminal environment by secretion and reabsorption of several substances (for review see Turner 1995). In this sense, it has been proposed that the epididymal epithelium secretes taurine/hypotaurine into the lumen (Hinton 1990; Holmes et al. 1992a). These amino acids would be later transported into sperm cells during their passage along the epididymis (Holmes et al. 1992a). These secretion/transport mechanisms have been suggested to be the main source for the high intracellular levels of these amino acids in sperm (Holmes et al. 1992a). This hypothesis is supported by the fact that the testicular fluid composition in taurine/hypotaurine is similar to circulating fluids (in μM quantities), whereas epididymal fluid shows a higher concentration (in the mM range) of these amino acids (Johnson et al. 1972; Hinton 1990). In accord with this notion, it has been shown that sperm cells are able to take up taurine by some β-amino acid transport system (Holmes et al. 1992a). The results of the present study appear to support this hypothesis because taurine was found in some epithelial cells at specific segments of the ducts studied. Taurine was found mainly in the efferent ducts, whereas most of the cells from other extratesticular segments (rete testis, epididymis, and ductus deferens) and the germ cells of the seminiferous tubules were immunonegative. However, in agreement with these results, the specific region that could secrete taurine into the lumen appears to be the efferent ducts rather than the epididymis. Moreover, an osmoregulatory role for taurine in the efferent ducts can be postulated, in agreement with the reabsorptive function of this organ (Ilio and Hess 1994; Hess et al. 1997).
Taurine/hypotaurine in sperm cells could originate not only via amino acid transport from the duct fluids but could also be synthesized from cysteine via the cysteine sulfinic acid decarboxylase (CSAD) pathway. Previous studies suggest a correlation between hypotaurine content and CSAD activity in the reproductive tissues of guinea pigs, being both higher in the epididymis and lower in the testis (Holmes et al. 1992a). Moreover, it appears that epididymal sperm could not synthesize these amino acids (Holmes et al. 1992a). All these data suggest that the biosynthetic route probably does not account for the origin of these amino acids in sperm but could be important in other cells of the reproductive organs.
The rat ductus epididymis is lined by an epithelium consisting of two main cell types (principal and basal cells) and other less common cell types, such as apical, narrow, and halo cells. Halo cells are actually considered intraepithelial lymphocytes and macrophages (Wang and Holstein 1983). According to our results, taurine is located in the intraepithelial lymphocyte-like cells and macrophages found in the efferent ducts, the epididymis, and the rete testis. Therefore, these taurine-containing cells probably correspond to the cell type termed halo cell. Moreover, these results are consistent with the known relation of taurine with the immune system. In this sense, cats (which have low activity levels of CSAD and are therefore dependent on a dietary source of taurine) treated with a low-taurine diet show changes in the proportion and function of leukocytes (for review see Sturman 1993). Furthermore, leukocytes show high concentrations of taurine, related to the modulation of the production of some mediators of the inflammatory/immune response (Marcinkiewicz et al. 1995; Park et al. 1995).
Apical and narrow cells are a minor population of epididymal epithelial cells. Although their function is unknown, it has been suggested that they could modify the luminal environment in which spermatozoa undergo maturation by cooperation with principal cells in reabsorption of testicular fluid (Sun and Flickinger 1980) and by acidification of the epididymal fluid (Kierszenbaum et al. 1981). The notion that these cells have a role in luminal acidification by secretion of protons, thus aiding in the quiescence of sperm motility, is based on the localization of carbonic anhydrase in these cell types (Cohen et al. 1976; Kaunisto et al. 1995; Adamali and Hermo 1996). Because taurine has several roles in transmembrane ion movements, we propose that the presence of taurine in these cell types could be associated with their function in the acidification of the luminal fluid through ion-exchange mechanisms. In this sense, it has been demonstrated that taurine uptake/release from several cell types occurs through an anion exchanger (band-3) protein (Martín del Río et al. 1996; Martín del Río and Solís 1998) which catalyzes transmembrane HCO- 3 movements. Furthermore, other mitochondria-rich cells such as stomach parietal cells, which present carbonic anhydrase and band-3 protein, also show high concentrations of taurine (unpublished data). Moreover, because apical, narrow, and halo cells contain abundant taurine, whereas other epithelial cells are immunonegative or less intensely immunolabeled, the use of taurine as a specific marker molecule for these cell types is suggested.
Leydig cells are the cellular source of androgen, i.e., testosterone, which is necessary for the regulation of spermatogenesis in mammals. Previous studies have demonstrated a relation between hypotaurine and testosterone. Hypotaurine almost disappears from the prostate and seminal vesicle of the guinea pig, rat, and mouse after castration, and its concentration is restored to almost normal values by administration of testosterone (Kochakian 1973, 1975, 1976). The immunolocalization of taurine in Leydig cells appears to support the existence of a poorly known relation between the physiology of testosterone/Leydig cells and taurine. Because the steroidogenic pathway involves the production of free radicals and lipid peroxidation (Peltola et al. 1996), the presence of taurine in Leydig cells could be related to its antioxidant capacity.
In neurons of the rat spinal cord, variable taurine immunostaining in their nuclei has been reported, being weak in some cells and as strong or stronger than in the cytoplasm in other cells (Lee et al. 1992). A similar labeling pattern is documented here in Leydig cells and in other cell types. These data suggest that whatever the functional role of nuclear taurine is (no hypothesis has yet been proposed), it should be highly dynamic, rather than static, and should involve variations in the nuclear taurine content under modifications of cell physiological conditions.
This study is the first to demonstrate the localization of taurine in the male reproductive organs. Such a specific distribution implies that taurine may play important roles in the male reproductive system and highlights the need for further investigation into the functions of taurine in the reproductive organs.