
Abstract
For detailed study of complex structures such as corpuscular mechanoreceptors, confocal microscopy can be used with multiple immunolabeling that identifies specifically different subcomponents. In addition, anatomic interpretation is enhanced by three-dimensional reconstruction. Confocal laser micrographs, reconstructed from serial images 1 μm thick of human skin Meissner corpuscles simultaneously immunostained for neurofilaments (NF 70-200) and protein S-100 (PS-100), clearly reveal the complex 3D relationship between Schwann-related lamellar cells immunoreactive for PS-100 and the nerve fibers marked by NF 70-200. The nerve fiber, after branching into the corpuscle, divides into several ramifications, presenting discoidal expansions and flattened fringed sections. The mean nerve diameter was 4 μm ± 1 (2-5 μm) and the mean size of the discoidal expansions was 15 μm ± 1 (7-30 μm). Corpuscle size varied from 30-140 ± 1 μm in length and from 20-60 ± 1 μm in diameter. This study confirms the presence of neural discoidal areas in Meissner's corpuscles, which are probably involved to some extent with the transduction process. Despite the accuracy of immunolabeling and imaging, an extracorpuscular neural network was never observed in the vicinity of corpuscles, thus giving doubt as to their existence.
Keywords
S
At present, immunolabeling techniques are the most reliable means for identifying neural structures. Of the many neuropeptides and proteins that have been reported in sensory endings, protein S-100 (PS-100) has proved to be a reliable marker of Schwann cells, and nerve fibers are labeled specifically by antibodies against neurofilaments (NF 70, NF 70-200) (Iwanaga et al. 1982; Stefansson et al. 1982; Dalsgaard et al. 1984; Hachisuka et al. 1984; Björklund et al. 1986; Haro et al. 1991; Karanth et al. 1991; Ramieri et al. 1992; Kennedy and Wendelschafer-Crabb 1993).
For complex structures such as corpuscular mechanoreceptors, simultaneous specific labeling by different markers allows better identification of the different substructures, thus improving anatomic interpretation (Dubövy et al. 1993; Vega et al. 1996). At present, acquisition of serial optical sections by confocal laser scanning microscopy (CSLM) combined with three-dimensional (3D) reconstruction techniques provides high-contrast images that depict microscopic anatomy more realistically than was previously possible (Mayhew 1992; Castano et al. 1993, 1995; Rumio et al. 1995b). In addition, with appropriate software, image analysis is improved by the possibility of virtual rotation of the object in space, allowing observation of the reconstruction in toto from any desired viewpoint (Cheng and Summers 1990; Carlsson 1991; Parazza et al. 1993). This study used double immunolabeling and CSLM to investigate Meissner corpuscles in human fingers.
Materials and Methods
Tissue Preparation
Human digital pulps of two second digits were obtained from non-replantable traumatic amputations on the dominant hand of two manual workers (43 and 51 years old) without medical antecedents who gave their informed consent in accordance with the Helsinki Declaration of 1975. Samples were immediately excised and fixed with 4% formaldehyde in pH 7.4 PBS 0.01 M for 24 hr at 4C. After rinsing in distilled water, pulp tissue was cut into small fragments perpendicular to the skin surface (1 times; 6 times; 3 mm) and immediately embedded in agar or paraffin. Thick serial sections (70 μm) were made from agar-embedded samples with a vibratome (Vibraslice 752 M; Campden Instruments, Oxford, UK) and treated for immunolabeling. Thin (7-μm) sections for conventional light microscopy were cut from paraffin-embedded samples with a microtome (Reichert-Jung 2050 Microm; Vienna, Austria).
Immunofluorescence and Immunochemistry
Agar-embedded samples were labeled using a floating technique. To enhance marker penetration, samples were pretreated by microwaving (Micromat 120 AEG; frequency 2.45 Hz, 850 W) in a 0.01 M pH 6 sodium citrate buffer (three sessions of 5 min each). After incubation in 0.01 M PBS with 2% skimmed milk powder at 4C for 1 hr and rinsing, primary antibodies were applied for 48 hr at 4C in a humid atmosphere with gentle agitation. Immunolabeling was performed with murine monoclonal antibodies (clone 2F11) to NF 70-200 (Chemical Credential; Costa Mesa, CA) at a dilution of 1:100 and with rabbit polyclonal antibodies to PS-100 (Immunotech 1071; Marseille, France) at a dilution of 1:48. After rinsing in 0.01 M PBS, pH 8.6 (three times for 20 min) and incubation in 1% bovine serum albumen (BSA) for 30 min, a second rinse in 0.01 M PBS, pH 8.6 (three times for 20 min) was done. Secondary fluorescence-conjugated antibodies were sequentially applied at 4C in a humid and dark atmosphere for 3 hr with 0.01 M PBS, pH 8.6 (1 hr) and 1% BSA (30 min) rinses between the two sequences as follows: first 1:100 rhodamine-conjugated (TRITC) goat anti-rabbit antibodies (Jackson; West Grove, PA) were used to recognize NF 70-200 and then 1:75 fluorescein-conjugated (FITC) rabbit anti-murine antibodies (Dako; Glostrup, Denmark) were applied to identify PS-100. After rinsing in 0.01 M PBS, pH 8.6 (1 hr), samples were mounted in an antifading medium (0.05 M Tris-HCl, 8%, pH 7.6, glycerol (90%), 1.4 diaziabyclo-octan (2%) (Sigma, St Louis, MO; D 2522) between a slide and a coverslip sealed with nailpolish. Control sections were prepared by substituting normal serum for primary antibodies or by eliminating the first incubation.
For light microscopy, thin sections were treated with the same immunochemical process as described above for PS-100 antibodies (Immunotech 1071). Specific staining was revealed by incubation with biotinylated donkey anti-rabbit antibodies (Jackson) followed by a peroxidase-conjugated avidin-biotin complex (1: 400, PBS 7.4, BSA 1%, 1 hr) (StreptABComplex/HRP; Dako) and diaminobenzidine tetrachloride baths. Counterstaining of cell nuclei was performed with hematoxylin.
Confocal Microscopy
Confocal optical sections were collected with a confocal scanning laser microscope LSM 410 Invert (Carl Zeiss; Oberkochen, Germany) using a X40 objective lens (Plan-Neofluoar, 1.3 NA, oil immersion). Simultaneous imaging of FITC and TRITC fluorescence was performed with the following settings. Both labels were excited with the 488-nm line of an argon laser while the excitation beam was separated from emitted fluorescence by a 510-nm dichroic mirror. The two fluorescences were segregated with a 560-nm beam separator and respectively selected with a 520-560-nm bandpass filter (FITC spectrum) and a 590-nm longpass filter (TRITC spectrum). The signal-to-noise ratio was improved by averaging eight ordered frames for each optical section. During the scanning session, correction of attenuation of emitted and reflected light due to sample penetration was accomplished by gain compensation of the photomultiplier (voltage magnification). Neither fluorescence bleached appreciably even during long exposures (up to 15 min/50 optical sections).
Tridimensional Reconstruction
The stacks of confocal images were edited, processed, and reconstructed with the Edit3D software developed in our laboratory (Parazza et al. 1993). This program runs on various computer platforms and operating systems: SGI workstations (IRIX 5.x,6.x), PowerMacintosh microcomputers (MacOS and LinuxPPC), and Intel PCs under Linux.
Processing
After collecting the confocal series, it was necessary to process these data to correct for image degradation inherent to confocal microscopy. Two main image artifacts were taken into account: (a) poor signal-vs-noise ratio (SNR) and (b) loss of fluorescence intensity as we penetrated deeper inside the thick tissue sections.
A 3D median filter was used to reduce the noise. This filter has the property of removing “salt and pepper” noise while preserving edge information in images. It uses a 3 times; 3 times; 3 window centered on the voxel (volume element) to be filtered, sorts the intensities of the voxels within the window, and returns the median value. This median intensity value is then substituted to the original value of the current voxel.
Although the biological samples are usually transparent they tend to absorb and scatter a small amount of light. In tissue sections in which thickness is greater than 5 μm, the absorption phenomenon is no longer negligible. On the one hand, the excitation light is gradually attenuated as a function of depth on the way in. On the other hand, the emitted light is also attenuated on the way out. This results in a real decrease in the signal intensity as a function of Z. The curve of attenuation of fluorescence intensity as a function of depth can be modeled by a log-logistic function (Rigaut and Vassy 1991). The correction consisted of multiplying the intensities of the voxels by a coefficient whose value increased with depth according to the following formula:
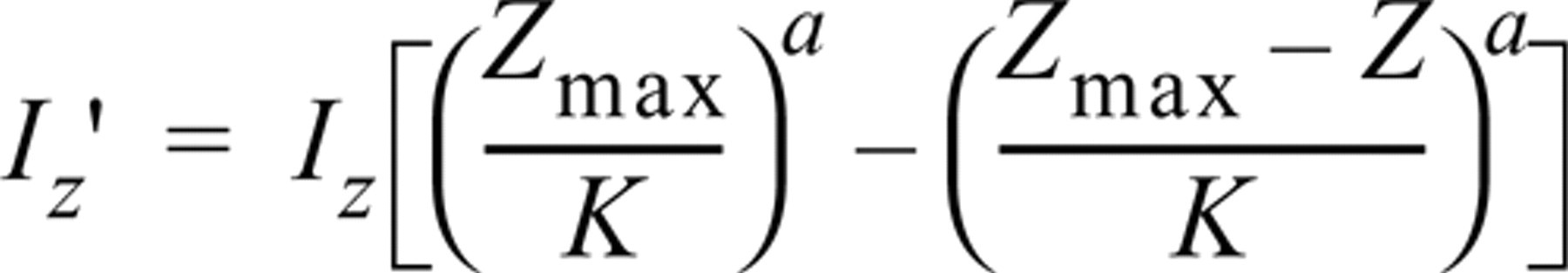
with Z the current depth, Zmax the deepest section of the series, Iz the voxel intensity at current depth, and I ′ z the new voxel intensity. K and a are empirically defined constants.
Reconstruction
Different visual attributes of the 3D reconstruction can be selected to best visualize specific details. According to the needed attributes (shape or content of the object), a surface or a transparency-rendering approach can be used. Surface rendering is obtained by simulating bundles of parallel rays of light traversing the pixels (elementary points) of the viewing screen of the computer. When one of these rays hits a surface voxel of an object (e.g., nerve fibers), the intensity of the corresponding screen pixel was modulated as a function of the cosine of the angle between the incident ray and the normal to the object surface at the hit spot. The voxels of the object were first obtained by applying a simple intensity threshold rule: All voxels whose intensity is greater than the given value are considered to belong to the object. The other voxels are labeled as background.
In contrast to the surface model in which only the surface voxels are considered, the volume model is intended to show all the information contained in the 3D cube. This display mode might be called “see-through” or transparent viewing. The method for representing transparent volumes is called “ray casting.” It consists of dividing the viewing screen into small pixels. Then we consider that each pixel of the viewing screen receives a ray of light that originated from infinity and that traversed the 3D data volume. The intensity of a given ray is the result of the information encountered along its path. In other words, the light intensities of all the traversed voxels are accumulated and contribute to the brightness and color of the pixel displayed on the screen.
Results
As deduced previously with conventional histological techniques, 3D reconstruction and double immunolabeling show the characteristic organization of Meissner corpuscles with a complicated interweaving of the helical nerve fiber winding within the stacks of lamellar cells, forming a spring-like framework (Figures 1A-1C). Corpuscles are usually found alone or grouped in a dermal papilla, with their long axis vertically oriented (Figure 1A). Lamellar cells are highly immunoreactive for PS-100 and are well differentiated from the nerve component labeled by antibodies against NF 70-200 (Figures 1C-1D and 2A-2B). The afferent nerve fiber can be followed along its extra- and intracorpuscular course. A single extracorpuscular nerve fiber can branch into several sensory endings (Figures 2A and 2B). Each corpuscle is penetrated through its base by dividing nerve ramifications which run in an helical fashion (Figures 2C-2D). The nerve fiber presents flat, discoidal, more or less fringed regions randomly situated along the nerve course, usually one per corpuscle (Figure 2E). These formations are not easily noticeable on single optical sections and are better appreciated after 3D reconstruction, particularly when the “nerve fluorescence channel” (NF 70-200) is observed alone (Figures 2C-2D).
On the other hand, labeled lamellar cells, which are arranged in stacks intercalated with the tortuous annulospiral nerve course, are more evident on a single optical section (Figure 1C) than in a 3D reconstruction, in which they have a more diffuse appearance (Figures 2A and 2B). During tomography sessions, the use of only one laser beam (argon) that activates both FITC and TRITC labels provides no blurring of the image. In addition, appropriate object rotation provides vision from another angle which modifies and corrects image interpretation (Figure 2E). Apparent discontinuities are probably artifacts caused by incomplete labeling of the sample or by photonic reflection defaults rather than a real structural defect (Figures 2A and 2B). Despite the quality of labeling that is sufficient to distinguish fibers with diameters of less than 0.8 μm, no extracorpuscular network has been identified in the vicinity of observed corpuscles (Figures 2A-2D). Measurements whose mean and SEM refer only to the optical 3D resolution give a nerve diameter that varies from 2 to 5 (mean diameter 4 ± 1 μm), with size differences which are more easily appraised on a stereoscopic view (Figures 2C-2D) whereas discoidal expansions have a mean diameter of 15 ± 1 (from 7 to 30 μm) (Figure 2E). Corpuscle size, measured from five samples (not shown), varies from 30 to 140 ± 1 μm in length and from 20 to 60 ± 1 μm in diameter.
Discussion
This study shows that double immunolabeling of thick samples observed by confocal microscopy allows reliable imaging of Meissner corpuscles. Many LM and TEM studies have demonstrated their fine structure, but analysis of this sophisticated framework could remain difficult (Cauna 1956a,b; Cauna and Ross 1960; Chouchkov 1973; Haschimoto 1973; Castano 1974; Iggo 1974; Halata 1975; Breatnach 1977). Reconstructions from 2D serial TEM observations were a first step to a better morphofunctional understanding, but at present CSLM coupled to 3D reconstruction appears to be the most reliable and accurate technique (Ventura and Castano 1977; Castano and Ventura 1978, 1979; Shotton 1989; Mayhew 1992; Parazza et al. 1993).
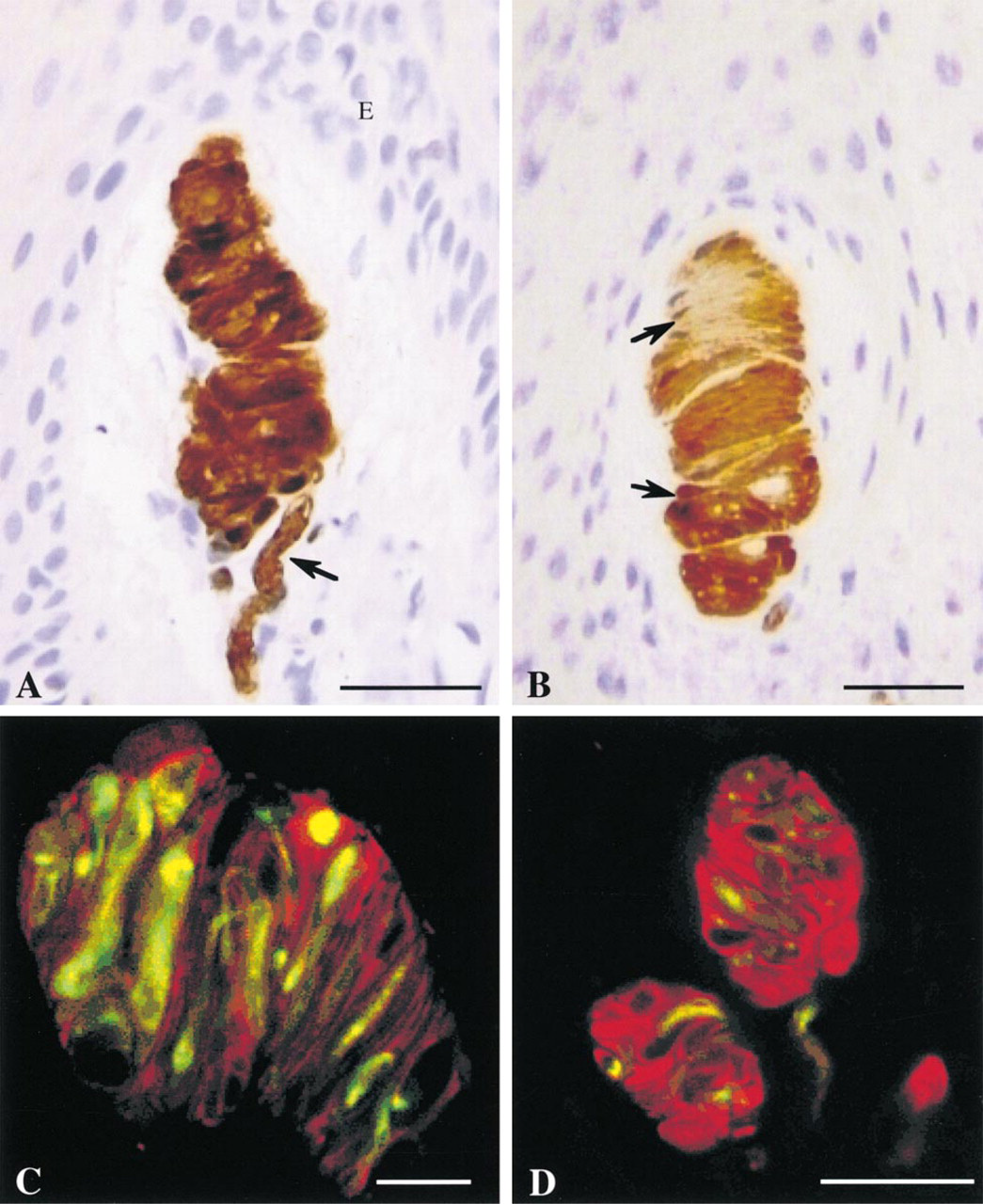
(A) Meissner corpuscles labeled with anti-PS-100 antibodies (peroxidase immunostaining). The complex spiral organization is evident. The corpuscle is connected to an afferent nerve at its basal extremity (arrow). The corpuscle is oriented vertically in a dermal papilla. E, epidermis. Thin paraffin sections (7 μm) of human digital pulp. Hematoxylin counterstaining. Light microscopy. Bar = 40 μm. (
Our 3D reconstructions are consistent with the results of Cauna (1956b) who stated that the nerve fiber, after having lost its myelin sheath on entry to the corpuscle, branches repeatedly without interconnections along its course. Despite the high image resolution, analysis was sometimes difficult, but in case of doubtful or difficult interpretation, elimination of one fluorescence channel (Figures 2C and 2D) reduced image complexity and enhanced anatomic analysis. However, it is impossible to be sure of complete marking of the neural structure, owing to the thickness of the sample and to random impregnation by antibodies. In addition to a possible anatomic interruption, nerve discontinuities can also be explained by the fact that thin regions, which may be poorer in neurofilaments, are less reactive to incident light and/or because of high absorption of the reflected light during its course through the skin sample (Cheng and Summers 1990; Carlsson 1991; Rumio et al. 1995b).
According to the labeling technique used, consistent visual differences can exist and image interpretation may become a matter of debate (Ramieri et al. 1992; Castano et al. 1995; Navarro et al. 1995). Comparing two labels, Rumio et al. (1995a) reported in a confocal study that the flattened and discoidal portions identified along the nerve course within the corpuscle looked different. With a PGP 9.5 label these regions appeared discoidal, and silver impregnation (Winkelmann technique) revealed them as slightly dilated segments or varicose elements that were less visible. The authors stated that these differences in aspect could be explained by the difference of staining mechanisms because these flattened discs and varicosities, which are rich in closely packed mitochondria, are less impregnated by the silver-lipoprotein precipitate that preferentially labels areas rich in neurofilaments. However, these expansions that appear as discoidal areas in TEM reconstructions are probably the same structures that have been identified with CSLM and immunolabeling (Ventura and Castano 1977; Castano and Ventura 1978, 1979).
Castano et al. (1995) reported with PGP 9.5 labeling that Meissner corpuscles can present several discoidal formations. However, we identified, from a sample of five (not shown), only one flattened disc per corpuscle with a smaller mean size (Figures 2C and 2D). This difference may be partly explained by the different staining precision between PGP 9.5 (neuroplasm protein) and NF 70-200 (neurofilaments from 70 to 200 kD) or, more probably, by individual variations. Several quantitative and qualitative studies have demonstrated that changes in Meissner corpuscle shape and size, nerve aspects, and lamellar cell organization are common with occupation, gender, aging, or nerve impairment in humans (Cauna 1956b; Bolton et al. 1966; Schimirgk and Rüttinger 1980; Matsuoka et al. 1983). Meissner corpuscles have also been commonly described as multiply (double or more) innervated in humans but our five observations were all simply innervated (not shown) (Cauna 1956b; Cauna and Ross 1960; Munger and Idé 1988; Saxod 1996). According to Matsuoka et al. (1983), 54% of Meissner corpuscles in human fingers are supplied by only one myelinated fiber, and the remainder are connected by two or more.
In 1896, Timofeew described thin accessory fibers accompanying human Meissner corpuscles in dermal papillae (Ruffini 1902; Cauna 1956b). In a primate TEM study, Castano et al. (1991) confirmed the presence of unmyelinated nerve fibers associated with Meissner corpuscles, but in a later human confocal study with PGP 9.5 immunolabeling they were unable to identify this extracorpuscular network. Rumio et al. (1995a) demonstrated thin fibers close to human Meissner corpuscles that were identified as intraepidermal nerve fibers (INFs). According to Björklund et al. (1986), these may be thin sympathetic nerves in the vicinity of Meissner corpuscles on the basis of a neuron-specific enolase-like (NSE-Li) and neurokinin A-like (NKA-Li) immunoreactivity. Several descriptions of such unmyelinated nerve fibers have been given in animal species, but convincing evidence is lacking in humans (Castano 1974; Idé 1976; Castano and Ventura 1978; Munger and Idé 1988; Castano et al. 1991). Despite the fact that very thin fibers of less than 0.8 μm in diameter can be isolated with CSLM, optical resolution limits may be one explanation of their absence in our reconstructions (Carlsson 1991). Nevertheless, the fact that neither of the antibodies against NF 70-200 and PGP 9.5 specific for myelinated and nonmyelinated fibers applied in the present and other confocal studies showed any thin extracorpuscular neural network does cast a serious doubt on its existence (Castano et al. 1995; Rumio et al. 1995a).
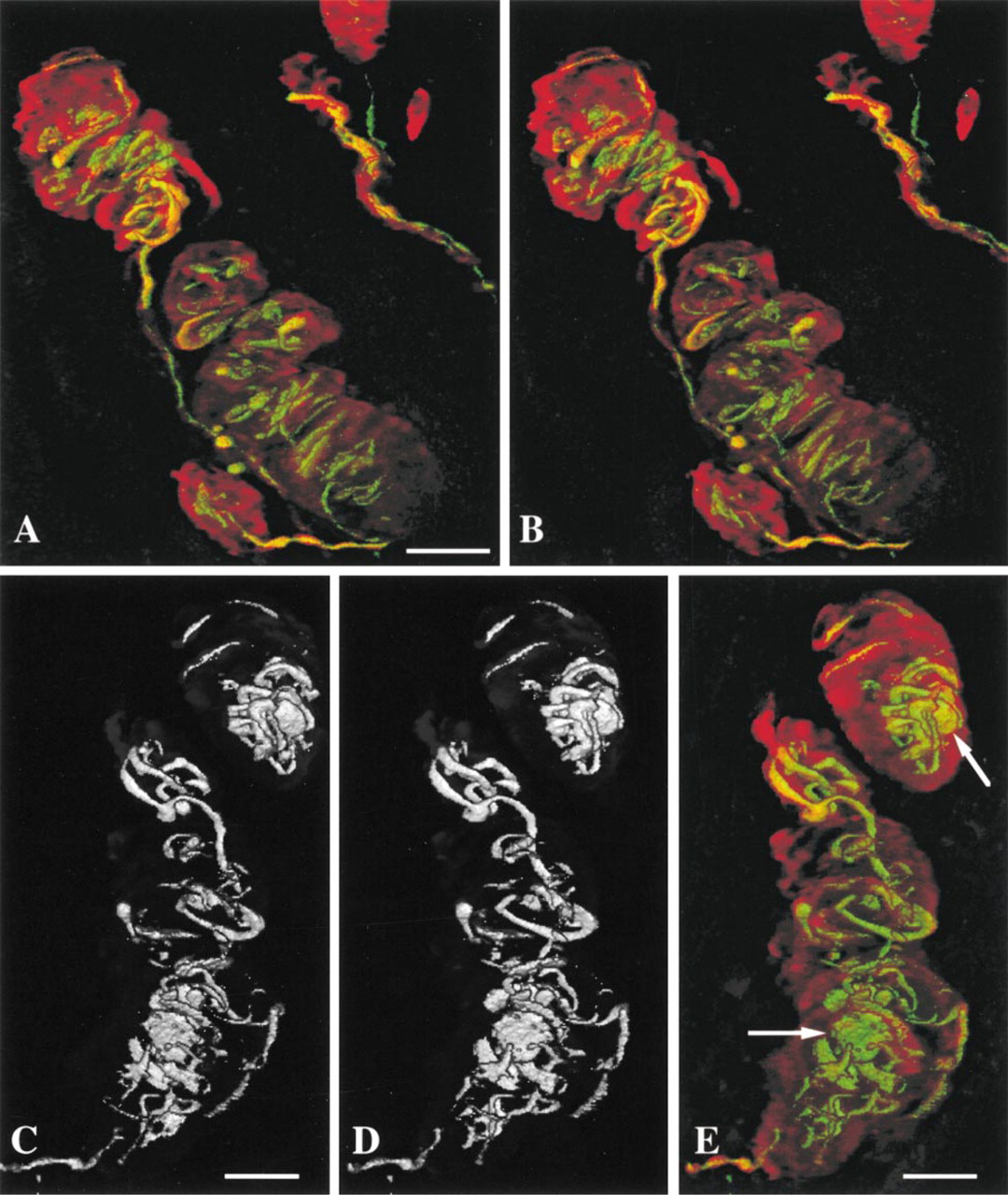
(A,B) 3D reconstruction of Meissner corpuscles labeled by fluorescent-conjugated antibodies against NF 70-200 (yellow-green) and PS-100 (red) obtained from 50 1-μm serial optical sections (CSLM, laser argon 488 nm).
In addition to the neural component, lamellar cells are another important part whose functional role is not yet completely understood. They are considered to be Schwann cell derivatives because they are labeled by specific Schwann cell markers (Iwanaga et al. 1982; Hachisuka et al. 1984; Munger and Idé 1988; Saxod 1996; Vega et al. 1996). The cell membranes are in close contact with the nerve fiber all along its course, but this complex stacking is sometimes difficult to interpret even on a clearly bi-colored 3D reconstruction (Figures 2A and 2B). Cauna and Ross (1960) showed that the nerve endings enter into a close appositional relationship with the flattened portions of lamellar cells and, in some instances, the cell and axolemmal membranes are thickened, with small packed vesicles on both sides. TEM descriptions have shown axonal spines along the longitudinal axis of the axon, microvesicules resembling synaptic vesicles, consistently electron-opaque membranes, localized Ca2+ in terminal axoplasm, and a concentration of packed mitochondria in nerve expansions (Cauna and Ross 1960; Haschimoto 1973; Castano 1974; Munger and Idé 1988; Tachibana et al. 1992). This particular intimate relationship between these flattened discoidal expansions and lamellar cells plausibly argues in favor of a transduction process during mechanical stress of the corpuscle at this level.
Footnotes
Acknowledgments
We wish to thank Dr F. Labat-Moleur MD for technical assistance, Prof F. Moutet (Hand Surgery Unit at Grenoble University Hospital, France) who provided us with human samples, and Prof J. Carlsson for his translation contribution.