
Abstract
Studying the regulation of peroxisome (Px) expression could improve our understanding of human peroxisomal disorders. The granulosa of the largest preovulatory quail follicles proved to be a relevant model because (a) Px expression changes according to the follicular maturation stage and (b) Px expression varies regionally according to the distance of the granulosa relative to the germinal disc region containing the female gamete (oocyte). The question was asked whether Px expression is related to the extent of metabolic cell coupling and whether zonal Px variation is causally related to oocytal factors. This was evaluated by the presence of catalase and Cx-43 (marker proteins for peroxisomes and gap junctions, respectively) and by in vitro experiments with granulosa explants. The data obtained show that the expression of Cx-43 and Px is inversely correlated both temporally and spatially. Uncoupling of gap junctions results in an upregulation of α-catalase immunofluorescence. This is in agreement with reports that gap junctions are often negatively affected by Px proliferators. The zonal gradient in Px expression appears to be imposed by the oocyte, as is the case for steroidogenesis and proliferative capacity in the granulosa epithelium.
P
The granulosa layer of the Japanese quail (Coturnix coturnix japonica) is a suitable model for study of Px expression in vivo because a steady decrease in expression was observed during the last days of follicle maturation, as well as a variation in Px expression according to the location of the granulosa cells relative to the germinal disc region, which contains the female gamete (oocyte). Furthermore, the granulosa layer provides a natural monolayer system embedded in its native extracellular matrix, i.e., between the vitelline membrane and the basement membrane. In granulosa explants cultured in vitro, cells remain coupled by extensive gap junctions.
In this study, two main questions were adressed. (a) Is the regional gradient of Px expression imposed by the germinal disc region containing the oocyte? (b) Is variation in Px expression related to the presence of gap junctions as evidenced by immunofluorescence of connexin-43, the most prominent gap junctional protein in the granulosa?
Materials and Methods
Reagents
Triton X-100, glycine, paraformaldehyde, sucrose, OsO4, K3(Fe(CN)6), 3,3′-diaminobenzidine, and all inorganic salts were purchased from Merck (Darmstadt, Germany). M-199 medium, bovine serum albumin fraction V, and 18 α-glycyrrhetinic acid were purchased from Sigma (St Louis, MO). Hepes was purchased from Acros (Geel, Belgium), dimethylsulfoxide (DMSO) from UCB (Leuven, Belgium), titaniumoxysulfate from Riedel-De Haën (Seelze, Germany) and penicillin-streptomycin from Gibco (Paisley, UK).
Peroxisomes were visualized by their marker enzyme catalase using a polyclonal antibody (IgG fraction) raised in rabbits against catalase isolated from bovine liver (code No. 200-4151; Rockland Laboratories, Gilbertsville, PA).
Gap junctions were visualized using mouse anti-connexin-43 (α-Cx-43) monoclonal antibody directed against amino acid positions 252-270 of the native rat connexin-43 (ProBio; Kent, UK); this monoclonal antibody was reported to crossreact with avian Cx-43 (Musil et al. 1990). Normal sera, secondary antibody conjugates, and fluorescent mounting medium were purchased from Dako (Copenhagen, Denmark).
Animals and Granulosa Sheets
The quail were given food [fresh lettuce and complete breeding food (cerafin Dossche, Belgium)] and water ad libitum. The animals were reared under continuous artificial illumination. Animal care procedures were conducted in accordance with the guidelines set by the European Community Council Directives 86/6091 EEC. After decapitation, the three largest preovulatory follicles (F1, F2, and F3 stage, 1, 2, and 3 days before ovulation with diameters of 18, 14, and 10 mm, respectively) (Callebaut 1983) were excised from the ovaries and rinsed in 0.9% saline. Granulosa sheets were removed as described by Gilbert et al. (1977), divided with iris scissors into 4-mm2 explants, and processed uncultured or after 24 hr of culture.
In the granulosa, three regions are defined as follows (see Figure 1). Region A comprises the animal pole (granulosa covering the germinal disc, i.e., the oocyte cytoplasm and nucleus). Region B is the transition zone adjacent to the animal pole and measures approximately one third of the diameter of the germinal disc. Region C includes the remainder of the vegetative pole. Regions B and C cover the yolk-containing part of the oocyte.
Techniques
Culturing Granulosa Explants. Explants were cultured serum-free in filter-sterilized M-199 medium (pH 7.4) supplemented with bovine serum albumin fraction V (BSA 0.1% w/v), penicillin-streptomycin (1% v/v), Hepes (10 mM), and sodium hydrogen carbonate (4 mM) and were incubated at 38C under saturated humidity (D'Herde and Leybaert 1997).
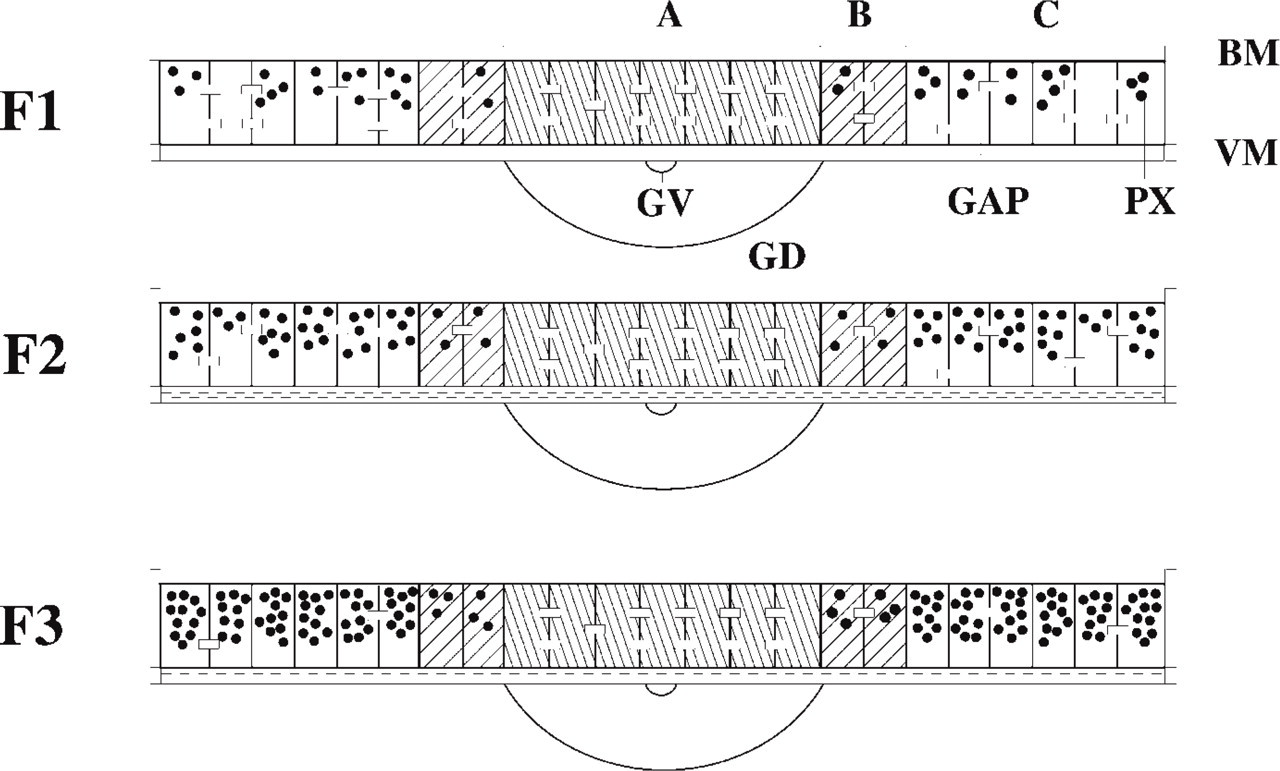
Schematic representation of the distribution of peroxisomes and gap junctions in the three largest preovulatory follicles in vivo. (
Immunofluorescence. Granulosa explants were rinsed three times in PBS (10 mM, pH 7.4), fixed for 20 min in 4% freshly prepared paraformaldehyde in PBS, rinsed again three times in PBS, and processed for immunofluorescence.
Staining of Peroxisomes. Membrane permeabilization was performed with 0.2% Triton X-100 in PBS for 15 min and by 1% glycine in PBS for 10 min (each step was followed by three rinses of 5 min with PBS). After incubating the granulosa explants for 45 min with 10% normal swine serum in PBS to prevent nonspecific labeling, explants were incubated with rabbit α-catalase (1:800) for 60 min at room temperature (RT), followed by five rinses in PBS, and incubated with swine anti-rabbit immunoglobulins-TRITC (1:60) for 45 min at RT. In negative control explants, normal rabbit serum was used in place of the primary antibody.
Staining of Gap Junctions. Granulosa explants were incubated with 3% BSA in PBS for 20 min and with 1% glycine in PBS for 10 min (each step was followed by three 5-min rinses with PBS). After incubating the granulosa explants for 45 min with 10% normal rabbit serum in PBS to prevent nonspecific labeling, explants were incubated with mouse α-Cx-43 (1:3000) for 60 min at RT or overnight at 4C, followed by five rinses in PBS, and incubated with rabbit antimouse immunoglobulins-FITC or -TRITC (1:60) for 45 min at RT. In negative control explants, normal mouse serum was used in place of the primary antibody.
Explants were mounted in fluorescent mounting medium and were examined on a Leica DM IRB/E inverted microscope equipped with epifluorescence optics, suitable filters for FITC and TRITC detection, and a MPS-60 camera.
Catalase Cytochemistry. Granulosa explants were fixed for 16 hr at RT in 4% formaldehyde buffered at pH 7.4 with 0.1 M sodium cacodylate + 1% CaCl2 (Roels et al. 1995). Explants were briefly rinsed in sodium cacodylate + 1% CaCl2 and in 13% sucrose before incubation with 3,3′-diamino-benzidine (DAB) in Theorell buffer, pH 10.5, for 3 hr at 25C. They were then postfixed overnight in 1% OsO4 in sodium cacodylate + 50 mM K3(Fe(CN)6). After dehydration, they were embedded in LX resin. Two-μm sections were examined by phase-contrast microscopy and ultrathin sections by electron microscopy (Jeol JEM-100B; Tokyo, Japan).
Catalase Activity. Granulosa sheets were rinsed twice in homogenization buffer [imidazol-HCl 0.01 M, pH 7.2, containing sucrose (8.5%), EDTA (1 mM), and ethanol (22 mM)] (Geerts et al. 1984). Sheets were resuspended in 500-1000 μl homogenization buffer and disrupted by sonication (six times for 15 sec) on ice using a Vibracell VC-130 sonicator resulting in an output of 4 W. Protein concentrations were measured according to Bradford (1976) using the Bio-Rad (Hercules, CA) protein assay kit. Homogenates were assessed for catalase activity with titaniumoxysulfate according to Baudhuin et al. (1964). Before measurement, samples were centrifuged for 3 min at 10,000 times; g to remove cell debris. Protein concentration and catalase activity measurements were performed with a microtiter plate reader model 550 (Bio-Rad) at 595 and 405 nm, respectively. Catalase activity was expressed as milli-units Baudhuin (Ub) per mg protein.
Experimental Design
Cultured vs Uncultured Granulosa. To study the effect of endo- or exogeneous factors (i.e., co-culture experiment and gap junction blocking experiment in this study) on the expression of Cx-43 and catalase, granulosa cells were kept in culture for 24 hr in serum-free medium. α-Catalase immunofluorescence was done on 27 follicles of 11 animals (uncultured) and on eight follicles of three animals (24-hr cultured).
α-Connexin-43 immunofluorescence was done on six follicles of two animals (uncultured) and on 13 follicles of five animals (24-hr cultured).
Double staining (α-catalase/α-connexin-43) was done on two follicles of one animal (uncultured) and on four follicles of two animals (24-hr cultured).
DAB staining was performed on 19 follicles of seven animals (uncultured) and on 15 follicles of five animals (cultured). The F1, F2, and F3 stages of four animals were used to assess the catalase activity in uncultured granulosa sheets, and the F1 stage of eight animals was used for the cultured sheets.
Co-culture Experiment. To examine the possible influence of the oocyte on the expression of peroxisomes by the granulosa cells, Region B was cultured for 24 h either separated from or still in continuity with the adjacent animal pole (Region A) of the same follicle (Figure 2). Four follicles from two animals were examined by α-catalase immunofluorescence and two follicles from one animal were examined after DAB staining by electron microscopy.
Gap Junction Blocking. To investigate the role of intercellular communication on Px expression, gap junctions were blocked by adding 200 μM 18 α-glycyrrhetinic acid (AGA) or 250 μM octanol during 24 hr of serum-free culture (Rose and Ransom 1997). DMSO was used as vehicle for both gap junction blocking agents. Controls consisted of vehicle-treated granulosa cell explants derived from the same follicle as the treated counterpart. For each agent, the vegetative pole of the F1 stage of four animals was used for α-catalase immunofluorescence, and the entire F1 sheets of four animals were used for catalase activity measurement.
Statistical Analysis
To obtain comparable data, each independent experiment was carried out on granulosa cell explants derived from the same follicle; follicles obtained from the same quail were used to compare the successive preovulatory stages. All experiments were repeated at least three times.
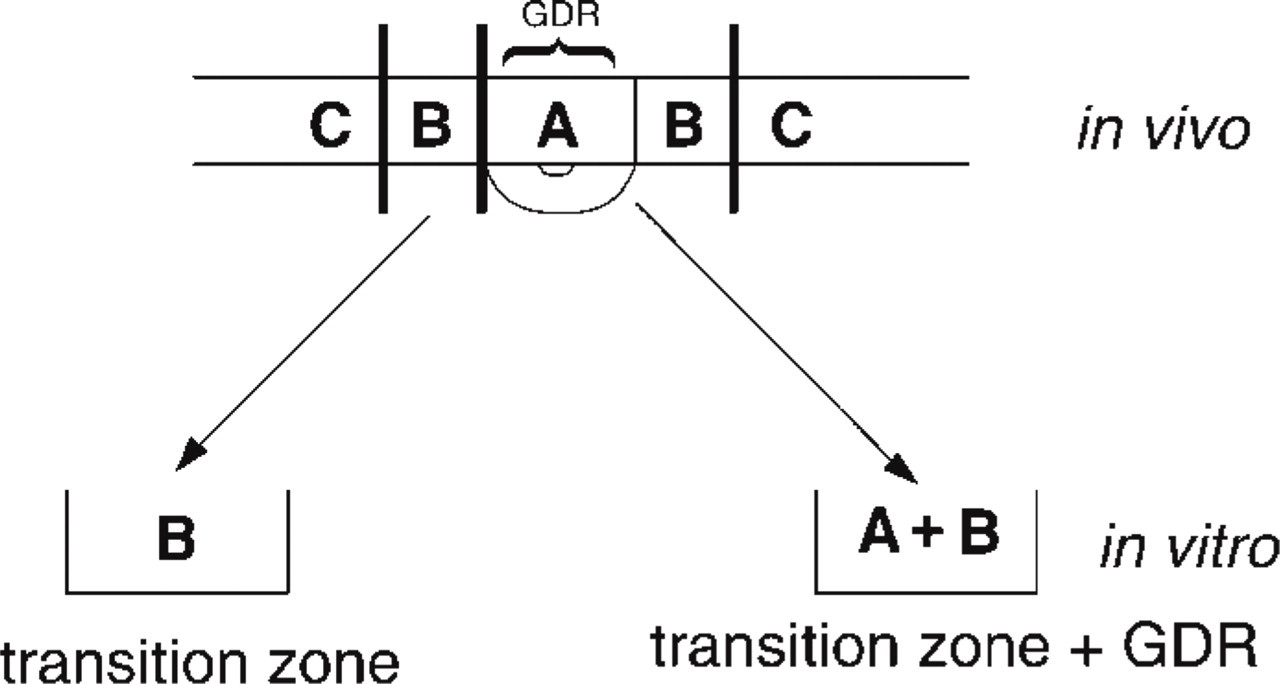
Granulosa from the same follicle cultured in presence or absence of the GDR (germinal disc region). Zone A; granulosa overlying the germinal disc (animal pole); Zone B, transition zone; Zone C, vegetative pole.
Numeric data are presented as means ± SD. The significance of difference among group means of follicles from different quails were analyzed using a t-test for independent samples. A t-test for paired samples was used to compare treated and untreated sheets derived from the same follicles. To compare the gap junction/cell ratio for the different follicle stages, a nonparametric Wilcoxon rank sum test was used. Statistical analysis was done with SPSS 6.1. Differences were considered statistically significant when p<0.05.
Results
Uncultured Granulosa
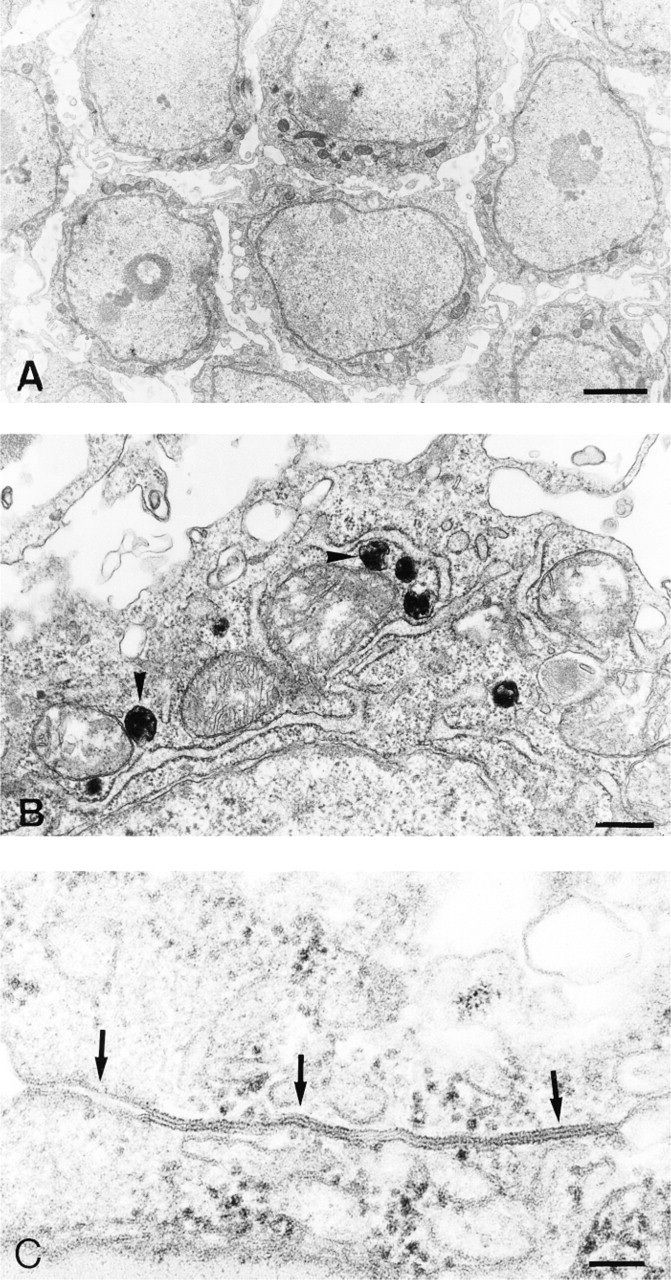
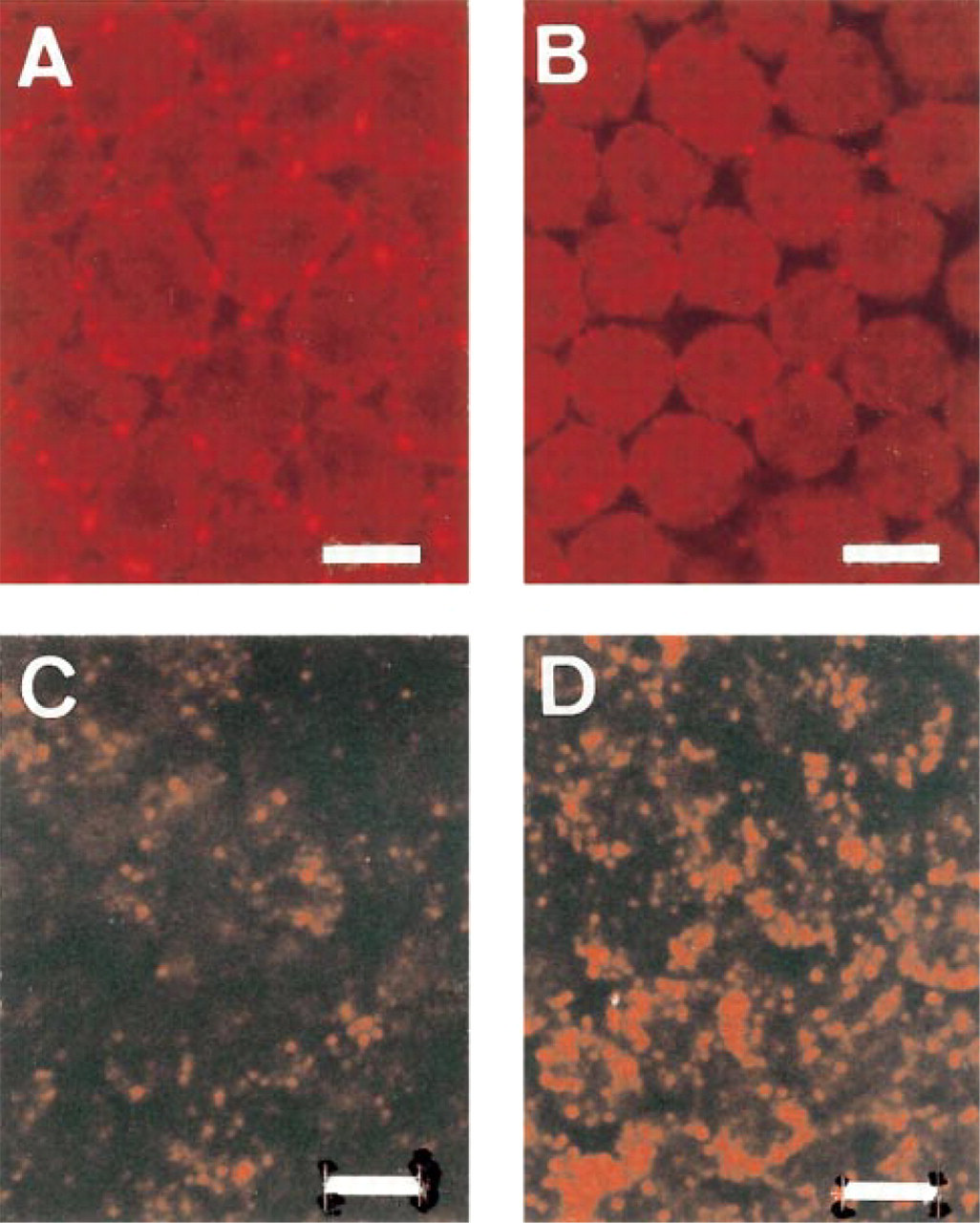
The distribution of peroxisomes and gap junctions in the uncultured granulosa displays a gradient, as summarized in Figure 1. At the animal pole (Region A) no catalase immunoreactivity is present, but in the transition zone B large isolated cells containing many catalase granules indicate the presence of peroxisomes (see Figure 6D). At the vegetative pole (Region C), most cells contain many small peroxisomes. Peroxisomes are concentrated at the basal side of the epithelium. Catalase activity is significantly higher in younger preovulatory follicles in the order F3>F2>F1. The three largest preovulatory follicle stages show statistically significant differences (p<0.02) as determined by a t-test for independent samples (Table 1). These data are also confirmed by α-catalase immunofluorescence (Figures 5C and 5D) and by electron microscopy. Peroxisomes are recognized by their catalase reaction product as small round or oval granules of about 0.2 μm (Figures 3A and 3B, and 4A and 4B).
Gap junctions visualized by α-Cx-43 staining show a completely inverse expression pattern compared to the peroxisome distribution. In an individual follicle, gap junctions are abundant at the animal pole, whereas they are few at the vegetative pole and show an intermediate expression pattern in the transition zone. When granulosa overlying the vegetative pole from three successive follicles (F3, F2, F1) from the same animal is compared, an increased Cx-43 expression pattern is observed during follicular maturation (Figures 5A and 5B), whereas catalase expression is decreased (Figures 5C and 5D). The numbers of both gap junctions and cells were scored by two independent observers on randomly selected color slides. The gap junction:cell ratios were significantly higher in F1 compared to F2 (p<0.05) as determined by the Wilcoxon rank sum test for independent samples (Table 2). The ratios of F2 and F3 were not statistically significantly different. As can be seen in Figure 1, the zonal gradient of gap junction expression is most prominent in the F3 stage. This gradient is less prominent in the F1 stage owing to the increase of Cx-43 in the vegetative pole during follicular maturation.
Double Staining Experiments With α-catalase/α-connexin-43
Double staining also visualized an inverse correlation between the expression of catalase and Cx-43 in individual cells (Figure 6A). Cells with abundant α-catalase immunofluorescence exhibited little or no α-Cx-43 immunofluorescence, and vice versa.
Granulosa After 24 hr of Culture
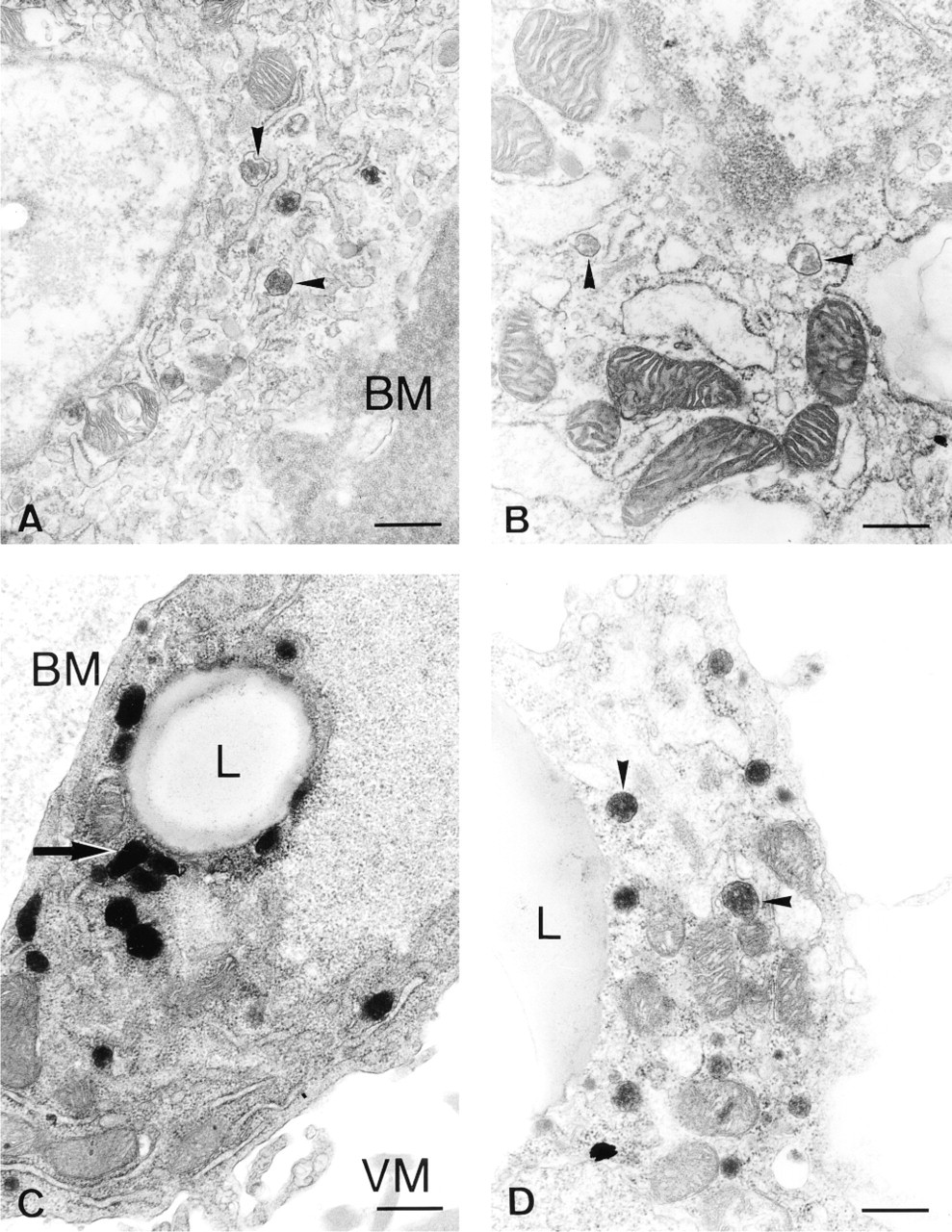
At the animal pole, granulosa cells still showed no catalase labeling similar to uncultured cells, suggesting absence of catalase-containing peroxisomes. Culturing the granulosa overlying the vegetative pole (Region C) led to a more polarized distribution of peroxisomes in the vicinity of lipid droplets, resulting in clusters of fluorescent granules (Figure 6F) compared to the randomly dispersed granules in the uncultured explants (Figures 5C, 5D, and 6E). By electron microscopy, some cells in F2 (Figure 4C) and F3 cultured granulosa showed clusters of peroxisomes with elongated shape or with tails, suggesting proliferation; in F1 such images were rare. The increase in Cx-43 expression at the vegetative pole coinciding with follicular maturation was maintained after 24 hr of culture. At the electron microscopic level, huge gap junctions were observed (Figure 3C). However, no significant increase in catalase activity could be detected in the granulosa of the F1 follicle after 24 hr of culture (control sheets of Table 4c,d) compared to the activity of uncultured F1 (Table 1a) (p>0.05 as determined by a t-test for independent samples).
Co-culture Experiments
Peroxisomes remained scarce in the transition zone of F1 cultured with the animal pole attached to it (Figure 6B), which is comparable to the in vivo situation. In contrast, peroxisomes became more abundant when this zone was cultured for 24 hr in the absence of the animal pole (Figure 6C). This was confirmed in F2 granulosa by counting the peroxisomes in individual cell profiles using electron microscopy (Table 3). Cultures with and without animal pole differed significantly in mean number of peroxisomes per cell (p<0.02) as evaluated by a t-test for independent samples.
Gap Junction Blocking Experiments
Culturing the granulosa explants of the F1 follicle in the presence of either gap junction blocking agent AGA (Figure 6H) or octanol (Figure 6I) increased the catalase immunofluorescence signal compared to the vehicle-treated control explants (Figure 6G). Catalase activity showed no difference between the treated cultures and their untreated counterparts (Table 4).
Regional variation of Px expression; diaminobenzidine cytochemistry for catalase activity in the granulosa layer. (
Influence of culture on Px expression; diaminobenzidine cytochemistry for catalase activity in the granulosa layer. (
Vegetative pole of uncultured granulosa explants. (
Discussion
Peroxisomes in the granulosa show a distinct expression pattern depending on the stage and the region of the follicle. The marker enzyme catalase decreases gradually with follicular maturation (F3>F2>F1) as detected by immunostaining, activity staining, and by activity assays. Within the same follicle, peroxisomes cannot be detected at the animal pole using α-catalase but they are abundant at the vegetative pole, with an intermediate expression at the transition zone between the animal and the vegetative pole.
Because the total catalase amount per sheet also decreases during follicular maturation, we assume that catalase synthesis is blocked; thus, catalase disappears as the result of turnover of the enzyme. However, there is also the possibility that the turnover of catalase is increased while synthesis remains unaffected. In the liver, the half-life of catalase is species-dependent and varies from 1.5 to 5 days (Price et al. 1962; Jones and Masters 1976; Geerts et al. 1984). Degradation of peroxisomes in toto via autophagocytosis was not observed, and inhibition of the enzyme can be ruled out because the antigen is equally disappearing, as detected by the decreasing α-catalase immunofluorescence signal.
The decrease in Px expression coinciding with approaching ovulation, as we have observed in quail granulosa, contrasts with the reported increase of Px expression in granulosa of large preovulatory pig follicles, which is culminating during the luteinization process (Peterson and Stevenson 1992). This reflects the different destiny of the preovulatory granulosa. After ovulation, granulosa in aves degenerates after 24 hr, whereas a corpus luteum develops in mammals.
It is known that the granulosa from the animal and the vegetative pole also differ in yolk transport mechanisms (Perry et al. 1978) and steroidogenesis. The absence of steroidogenesis, as quantified by progesterone production in the granulosa overlying the animal pole described by Tischkau and Bahr (1996), is accompanied by the absence of peroxisomes in this region, as shown in our experiments. In Leydig cells, Px volume is positively correlated with the testosterone secretory capacity of these cells (Mendis-Handagama et al. 1992). The detection of a sterol carrier protein (SCP-2) in peroxisomes of cells of the adrenal cortex (Magalhaes and Magalhaes 1997) also suggests that peroxisomes play a role in steroidogenesis. ACTH (adrenocorticotropic hormone) stimulation, which increases steroidogenesis, is also correlated with an increased number of peroxisomes in the adrenal cortex (Berchtold 1975; Russo and Black 1982). On the other hand, human peroxisomal deficiency disorders, such as X-linked adrenoleukodystrophy, lead to an impairment of steroidogenesis by the adrenal cortex (Van den Branden et al. 1993) and testis (Powers 1985). Furthermore, electron microscopy of hepatocytes indicated a preferential localization of the peroxisomes in the vicinity of lipid droplets (Novikoff et al. 1973), similar to our observations by both immunofluorescence (Figures 6F, 6H, and 6I) and electron microscopy (Figures 4C and 4D) in cultured granulosa cells. Lipid droplets contain cholesterol (produced by peroxisomal enzymes), which serves as a major substrate for steroidogenesis (Magalhaes and Magalhaes 1997).
Catalase activity in uncultured granulosa in relation to follicular maturation ∗
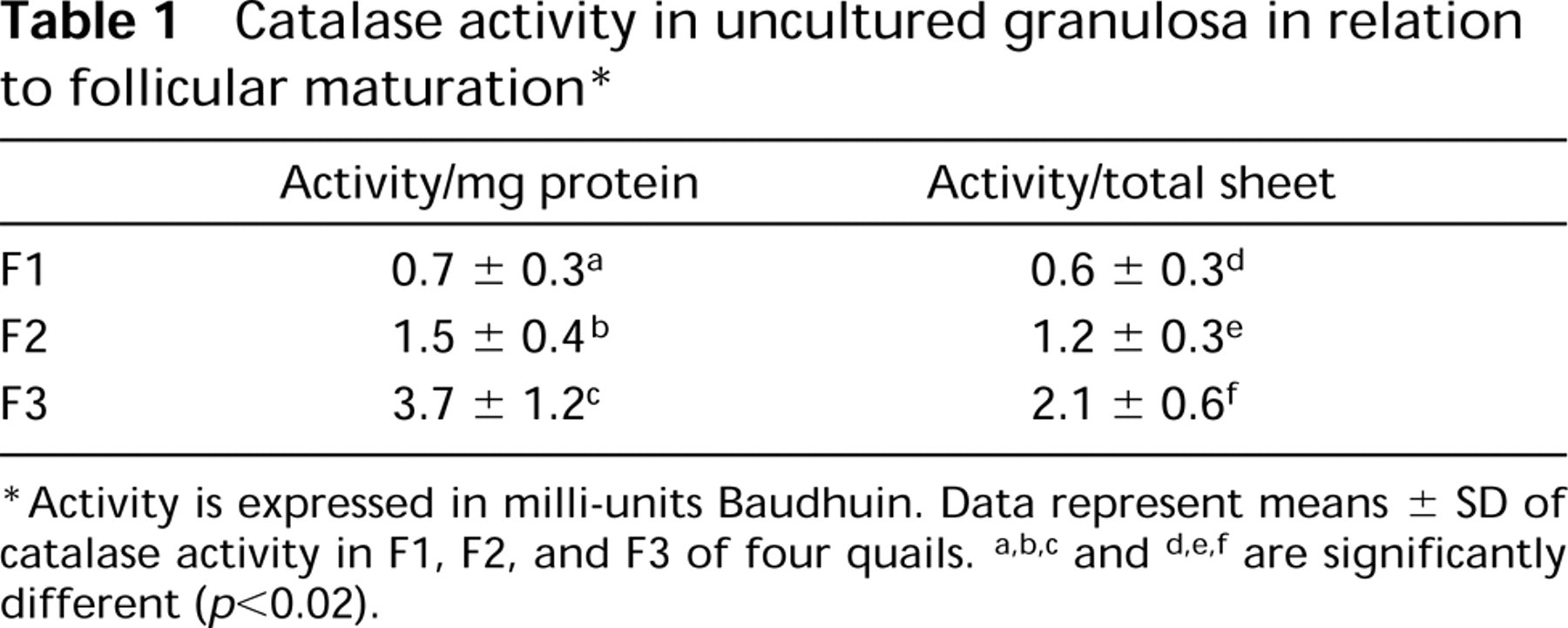
Activity is expressed in milli-units Baudhuin. Data represent means ± SD of catalase activity in F1, F2, and F3 of four quails. a,b,c and d,e,f are significantly different (p<0.02).
Catalase expression remains low in Region B co-cultured with the animal pole (Region A), in contrast to Region B from the same follicle which shows many peroxisomes when cultured without the animal pole. This could be explained by an oocytal factor that inhibits Px expression at the animal pole. The gradient of this factor, which decreases towards the vegetative pole, would explain the rising gradient of peroxisomes in the latter region. This inhibitory factor could be produced by the oocyte itself and might be propagated via gap junctional communication, because the oocyte is connected through gap junctions with the overlying granulosa as well (Dekel 1987; Yoshimura et al. 1993).
Another regulatory influence of the germinal disc (animal pole) on the physiology of the granulosa is proposed by Tischkau and Bahr (1996). Avian granulosa cells at the animal pole are immature, highly proliferative, and exhibit low progesterone production, whereas granulosa cells located distally to the germinal disc (vegetative pole) are less proliferative, more differentiated, and produce higher levels of progesterone. Experiments with germinal disc region conditioned medium disclosed a heat- and protease-sensitive factor that inhibits steroidogenesis and stimulates proliferation of the granulosa in a paracrine manner. Interestingly, mouse oocytes were also found to secrete paracrine factors affecting granulosa cell development and function (Eppig et al. 1993), and mouse oocytes were found to secrete factors that inhibit progesterone synthesis by the granulosa and are secreted in increasing amounts during follicular development (Vanderhyden and Macdonald 1998). These situations are quite analogous to our model in which an oocytal inhibitory factor could, again, be responsible for the absence of Px expression in the granulosa overlying the animal pole. Oocytal factors can easily reach the overlying granulosa through gap junctions (Yoshimura et al. 1993), but we must point out that an inhibitory factor of protein nature cannot be propagated via gap junctions, because these junctions allow passage only of small molecules up to 1 kD. However, small signal molecules inducing or induced by a Px inhibitory protein could be spread via gap junctions. Our experiments cannot exclude or confirm that the inhibitory factor (if any) is propagated through gap junction communication.
Gap junctions in uncultured granulosa in relation to follicular maturation ∗
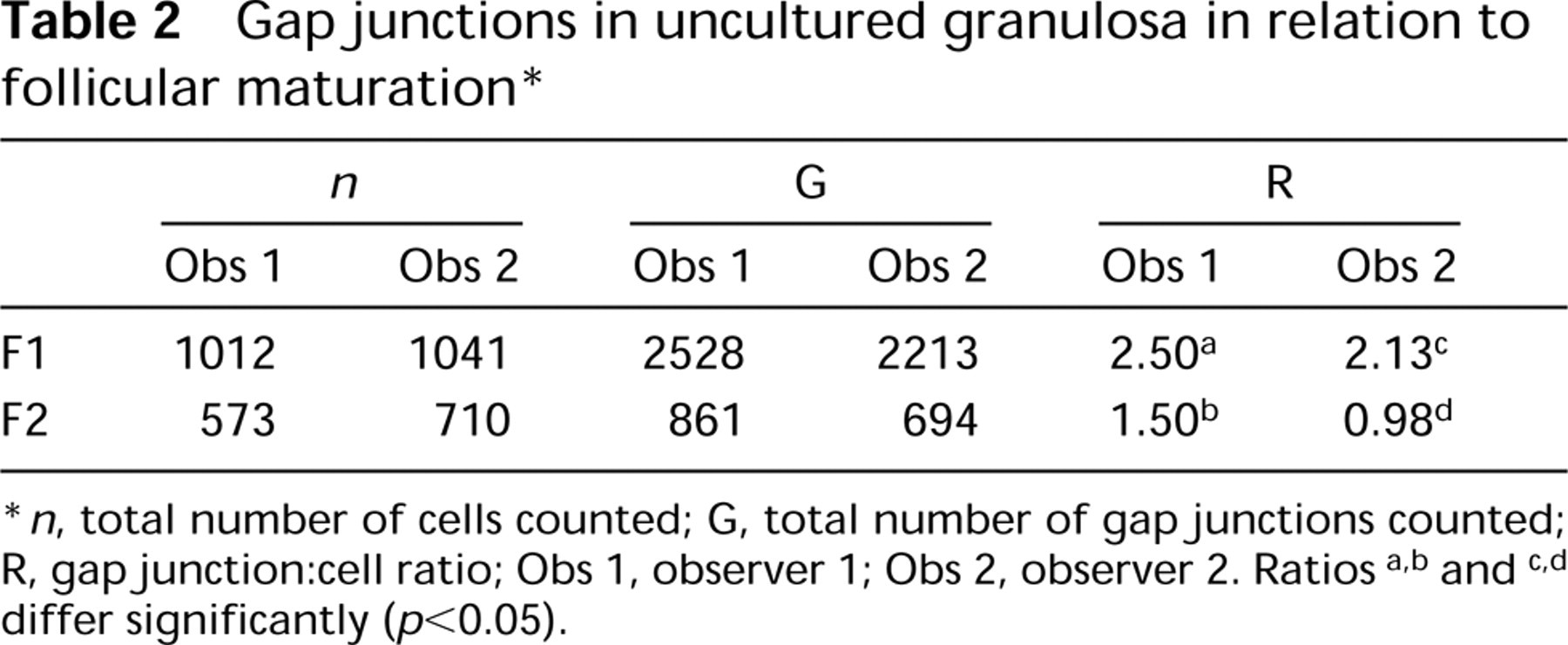
∗ n, total number of cells counted; G, total number of gap junctions counted; R, gapjunction:cell ratio; Obs 1, observer 1; Obs 2, observer 2. Ratios a,b and c,d differ significantly (p<0.05).
The role of gap junctions in the regulation of peroxisome expression is supported by our finding that blocking of gap junction communication by two distinct agents leads to a visible increase in catalase immunofluorescence. That the increase in catalase antigen is not accompanied by a significant increase of enzyme activity could be explained by the presence of newly synthesized catalase monomers lacking catalase activity. Degradation of the native catalase tetramer into its subunits is accompanied by the loss of catalase activity (Tanford and Lovrien 1961; Goldfischer and Reddy 1984), but the monomers preserve their immunoreactivity towards the polyclonal antibodies as shown in immunoblots (Wanders et al. 1995).
It may be significant that several Px proliferators (clofibrate, nafenopin, and perfluorinated fatty acids) have been reported to inhibit gap junctional communication (Leibold et al. 1994; Ito et al. 1996; Cruciani et al. 1997; Upham et al. 1998). This constitutes an independent indication of the inverse relationship between gap junctions and peroxisomes.
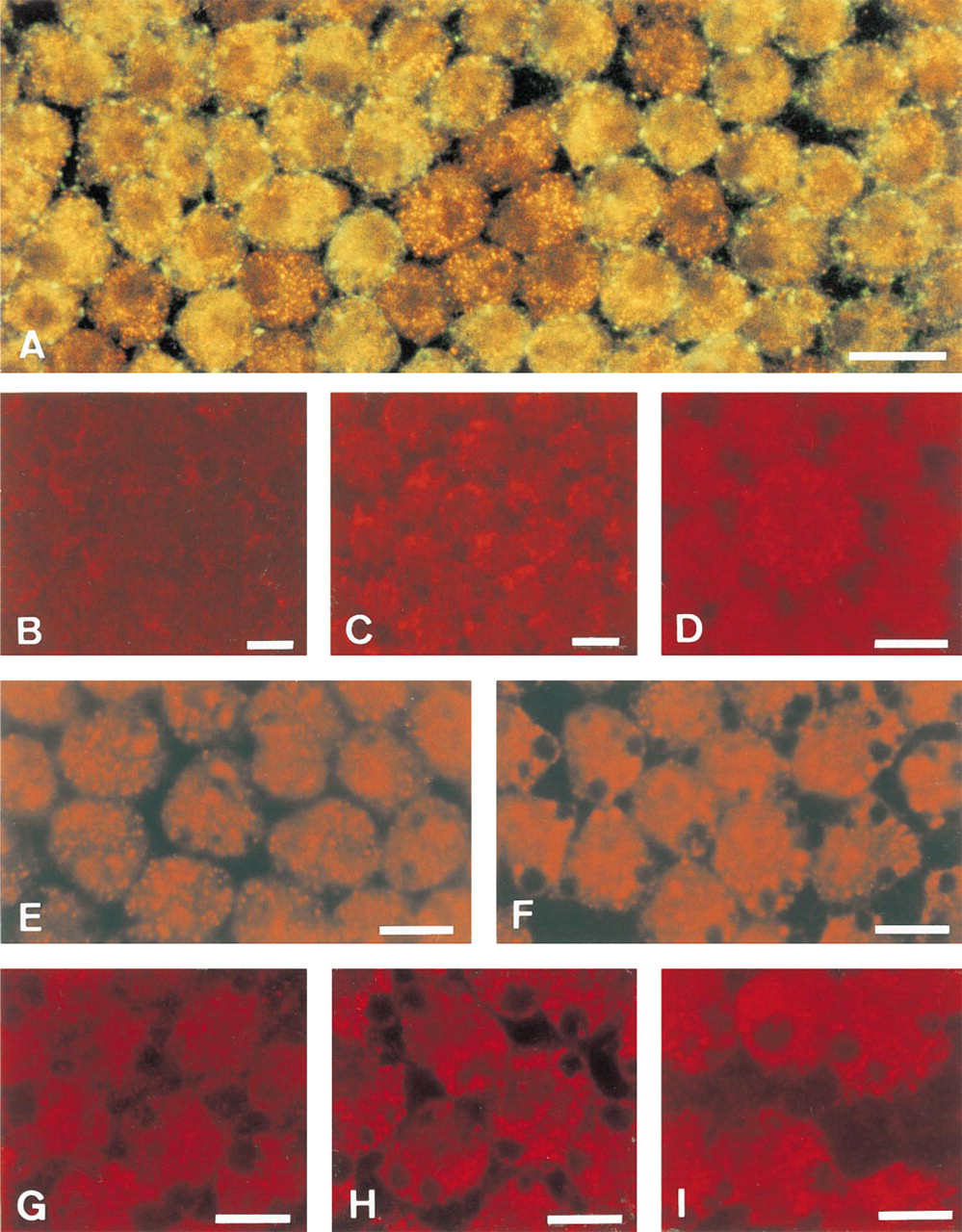
α-Catalase immunostaining (red label) (
Mean number of peroxisomes in individual cell profiles in ultrathin sections after culture in the presence or absence of the animal pole ∗

Region B + A, transition zone cultured with animal pole attached to it; Region B, transition zone cultured without animal pole; n, number of cells examined; Px, mean number of peroxisomes per cell profile ± SD. F2 Cultures witha and withoutb animal pole differ significantly (p<0.02).
Although we have previously suggested a peroxisome inhibitory factor propagating through increasing gap junctional communication, alternative hypotheses cannot be excluded. On the basis of the observation that peroxisomes and gap junctions are also inversely correlated in individual cells (Figure 6A), one could assume that the expression of both catalase and connexin-43 is inversely regulated at the cellular level. Other examples in which altered gap junction communication leads to altered gene expression have been reported. For example, the steroidogenic response of adrenal cells to ACTH is dependent on gap junction communication (Munari-Silem et al. 1995), and restoration of cell coupling by transfection of Cx-43 or Cx-32 increases insulin production in communication-deficient insulinoma (Vozzi et al. 1995) and increases thyroglobulin expression in thyroid cells (Statuto et al. 1997). Moreover, altered gap junction permeability caused by manipulation of the expression of Cx-43 and Cx-45 in osteoblastic cells alters transcriptional activity of osteoblast-specific promotors (Lecanda et al. 1998), although the nature of the mechanisms linking gap junction communication to gene expression remains elusive.
Both models are not contradictory and could even be combined. One or more factors (derived from the oocyte or the overlying granulosa), that have the ultimate effect of upregulating gap junction communication and downregulating Px expression might be propagated through progressing gap junction communication, starting from the animal pole and continuing over the remainder of the follicle.
Effect of gapjunction blocking agents on catalase activity a
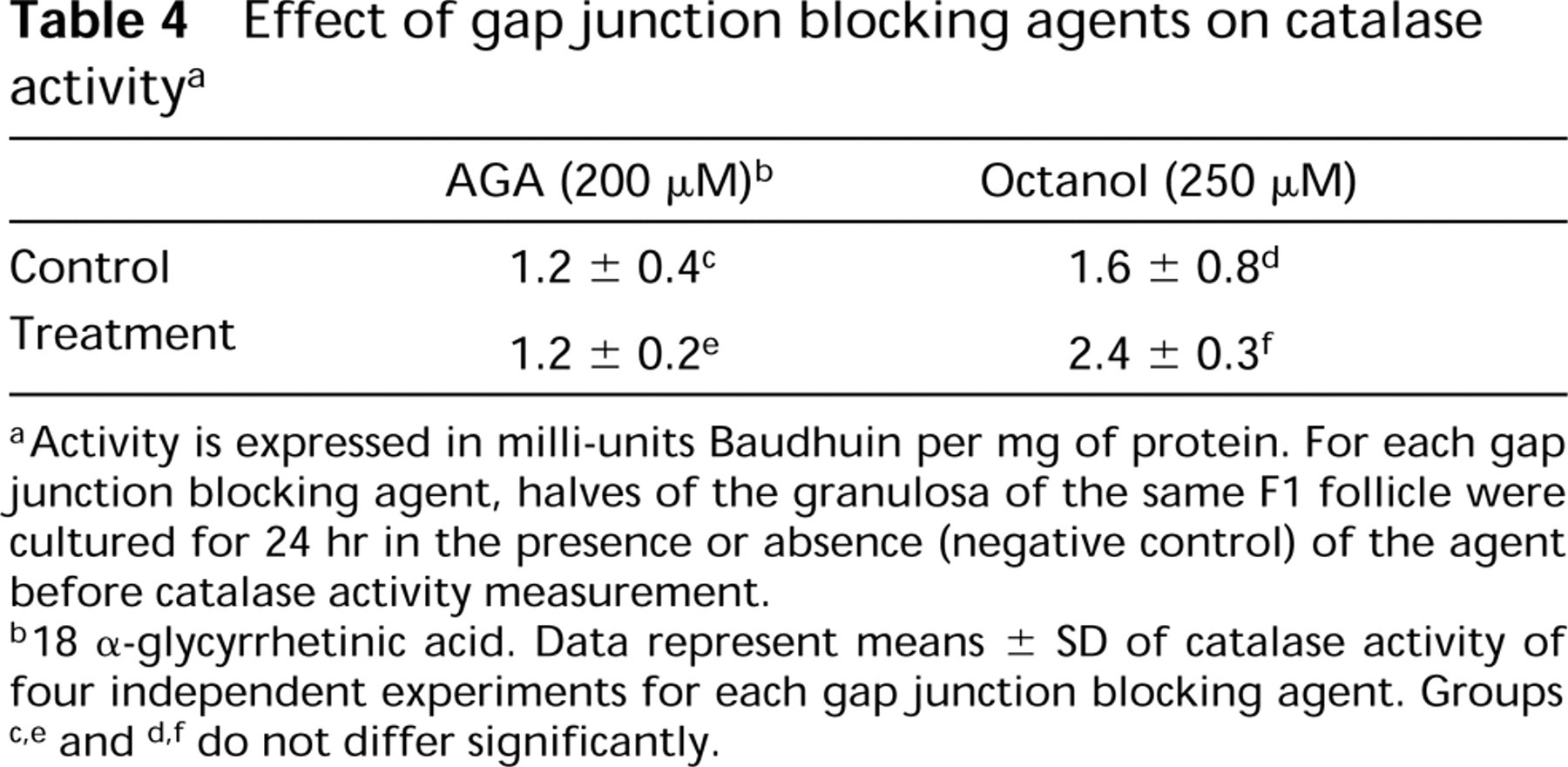
Activity is expressed in milli-units Baudhuin per mg of protein. For each gap junction blocking agent, halves of the granulosa of the same F1 follicle were cultured for 24 hr in the presence or absence (negative control) of the agent before catalase activity measurement.
18 α-glycyrrhetinic acid. Data represent means ± SD of catalase activity of four independent experiments for each gap junction blocking agent. Groups check thisc,e and d,f do not differ significantly.
Footnotes
Acknowledgments
Supported by the European Concerted Action “Peroxisomal Leukodystrophy” BMH4 CT 96-1621, by a grant from University “la Sapienza” (Roma) C.U.N. (SFV), and by the Research Council of the University of Ghent (BOZF No. 011B6197 and 011O6797).
We thank Ir Sylvie Mussche for the statistical analysis and Guido De Pestel, Betty De Prest, Simonne Vanhulle, and Noël Verweire for technical assistance.