
Abstract
Farber's disease (FD) is a rare genetic disorder caused by ceramidase deficiency, which results in ceramide accumulation in lung, liver, colon, skeletal muscle, cartilage, and bone. Although this disease has been symptomatically characterized, little is known about its molecular pathogenetic process. Because recent studies reported that ceramide accumulation induces GD3 ganglioside formation and apoptosis, we investigated, in tissue obtained via colonoscopy from seriously involved patients, the possible involvement of ceramide in FD colonocyte destruction. Histochemical and TUNEL analyses of paraffin-embedded sections revealed that 45 ± 4.3% of FD colonocytes showed morphological signs of apoptosis compared with the 8 ± 2.3% of constitutive epithelial cell death. Importantly, immunohistochemical study for pro-apoptotic factors showed that GD3 accumulation co-localized with active caspase-3 and cleaved K18 in FD colon tissue. These findings provide evidence for a role of the apoptotic ceramide pathway in the pathogenesis of FD.
F
Although joint pain is the most common symptom, more severely involved patients may have respiratory, gastrointestinal, and neurological impairments (Qualman et al. 1987; Levade et al. 1995; Moser 1997). Symptomatology is strictly related to histopathological alterations arising early in life. Granulomatous nodular neoformations appear at muscular and osteocartilagineous levels and cause joint deformities (Moser 1997; Qualman et al. 1987).
Ceramide accumulation results from impaired ceramide hydrolysis due to loss of function mutations of the acid ceramidase gene (Koch et al. 1996). This leads to cellular alterations characterized by lysosomal inclusions containing lamellar and curvilinear membrane (Farber bodies) (Chen and Decker 1982; Abenoza and Sibley 1987). This process is followed by granulomatous reaction associated with the presence of infiltrating macrophages, lymphocytes, and fibroblasts (Tanaka et al. 1979; Burck et al. 1985; Van Echten Deckert et al. 1997).
Ceramide is a cell signaling molecule involved in a variety of biological effects in different cell types, including cell proliferation and differentiation, inflammation, and apoptosis (Testi 1996; Green and Reed 1998). Recently, it has been shown that, during the apoptotic process, ceramide is recruited to the ganglioside pathway and converted into GD3 ganglioside by a GD3 synthase (De Maria et al. 1997). Accumulation of GD3 ganglioside is responsible for the generation of apoptotic signals in a caspase-dependent or -independent manner (De Maria et al. 1997). Moreover, cell-permeable exogenous ceramide has been demonstrated to induce caspase-3 (CPP32/yama/apopain) activation (Tepper et al. 1997; Anjum et al. 1998; Genestier et al. 1998).
Caspase-3 is one of the most specific proteases involved in the apoptosis mechanism (Patel et al. 1996). Caspases are initially produced as inactive precursors, which are activated after cleavage of inactive proenzymes into heterotetrameric subunits to initiate the proteolytic cascade (Schwartz 1998; Thornberry and Lazebnik 1998). Caspase-3 is activated early during apoptosis, and the activated form is a marker of cells undergoing apoptosis (Ballou et al. 1996; Schwartz 1998). Furthermore, keratins, in particular K18, a major component of intermediate filaments of epithelial cells, have been shown to be a target of caspases at the early phases of apoptosis (Caulìn et al. 1997).
In this study we investigated whether ceramide accumulation in FD triggers the apoptotic machinery in colon epithelial cells, thus contributing to the pathogenesis of this genetic disorder.
Materials and Methods
Specimens
Colon mucosal biopsies were obtained at the time of colonoscopy. The diagnosis of FD was based on clinical criteria and was confirmed by acid ceramidase deficiency in cultured white blood cells (Chatelut et al. 1996). Tissue was obtained from two patients with FD and from three healthy adults (age 42 ± 7.5 years).
Histochemistry
Tissues from each subject were fixed and stained with hematoxylin and eosin (H&E) and periodic acid-Schiff (PAS; Sigma-Aldrich, St Louis, MO) (Spicer 1987).
Immunohistochemistry
Immunohistochemical stains were performed on paraffin-embedded tissue sections (7 μm) obtained from FD colon and normal human colon specimens. Sections were dewaxed in xylene and dehydrated through graded alcohols (100% to 30%). Endogenous peroxidase was inhibited by exposing sections to 3% H2O2 for 10 min at RT. Then sections were rinsed for 10 min in distilled water. Before starting the immunostaining procedure, sections were treated with citric acid (2 g/liter, pH 6.0) for 15 min in the microwave for antigen retrieval.
Sections were incubated for 10 min with TBS containing 1% BSA to block aspecific staining. After elimination of excess serum, sections were exposed for 1 h to specific antibodies anti-Fas (DX2, mouse IgG1; PharMingen, San Diego, CA), anti-GD3 (S2-566, mouse IgM; Seikagaku, Tokyo, Japan), anti-CPP32 (clone 19, mouse IgG2a; Transduction Laboratories, Lexington, KY), anti-caspase-3, active form (polyclonal rabbit; kindly provided by Merk Frosst, Centre for Therapeutic Research, Merck Research Laboratories, Kirkland, Quebec, Canada), anti-M30 cytodeath (mouse IgG2b; Boehringer Mannheim, Mannheim, Germany) (Leers et al. 1999) or isotype-matched control at appropriate dilutions. After two washes in TBS, sections were treated with biotinylated anti-rabbit and anti-mouse immunoglobulins and subsequently washed in TBS and incubated with streptavidin-peroxidase (Dako LSAB 2 Kit; Dako, Carpinteria, CA). Detection was revealed using 3-amino-9-ethylcarbazole (AEC) as a colorimetric substrate. Counterstaining of tissue sections was performed using aqueous hematoxylin. Sections were rinsed in water, mounted in aqueous mounting medium, and analyzed by light microscopy.
In Situ Apoptosis Detection
TdT-mediated dUTP nick end-labeling (TUNEL) reaction was detected in serial paraffin-embedded sections. After dewaxing, sections were washed twice with PBS and cells permeabilized with 0.1% Triton X-100, 0.1% sodium citrate for 2 min on ice. Then, in situ apoptosis detection was performed with an In Situ Cell Death Detection AP Kit (Boehringer Mannheim; Indianapolis, IN). DNA strand break detection was done with an antifluorescein antibody conjugated with alkaline phosphatase (AP) and revealed by 5-bromo-4-chloro-3-indolyl-phosphate (BCIP; Dakopatts, Glostrup, Denmark). Eosin was used as counterstain. A positive control was carried out by pretreating the specimens with DNase I (1 μg/ml) to introduce nonspecific strand breaks (not shown). By contrast, the negative control was subjected to the same staining for TUNEL without terminal deoxynucleotidyl transferase (TdT) (Stassi et al. 1997a,b,1999). Photography was performed with a light microscope.
Quantification of Apoptotic Events
H&E-, PAS-, and TUNEL-stained sections were examined for the scoring of cells undergoing apoptosis, detected morphologically as membrane-bound apoptotic bodies and enzymatically as dark blue staining. The number of apoptotic events was randomly counted in 40 high-power microscopic fields (X40). Approximately 500 nuclei per colon mucosal biopsy were counted. The number of apoptotic cells was expressed as number per 100 nuclei. Comparison between the two groups was performed by Student's t-test.
Results
Ceramide Accumulation
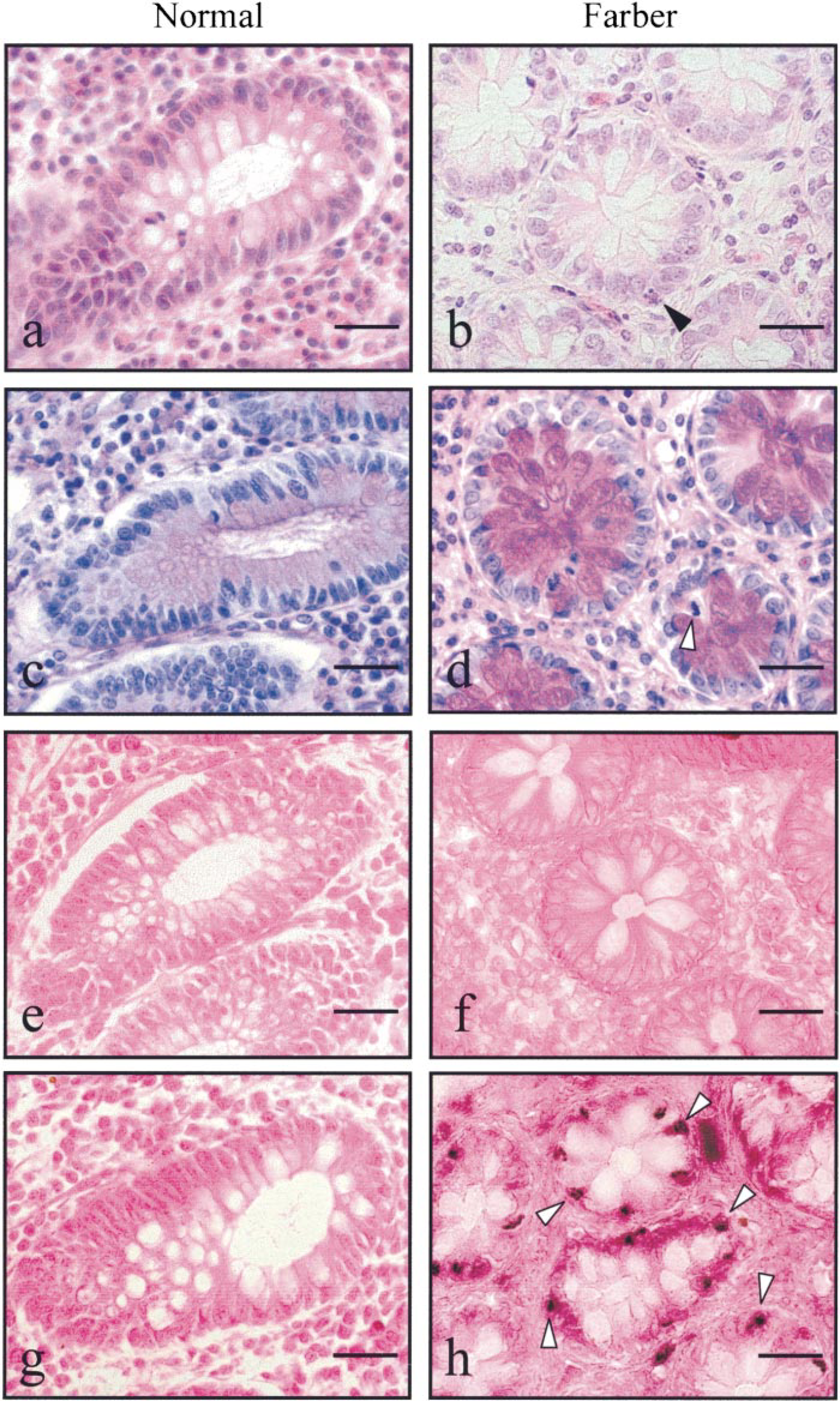
H&E staining showed normal crypt architecture in both normal and FD colon tissues, whereas lymphocytes, plasma cells, and eosinophils were numerous in the lamina propria of FD colon mucosa compared to colon of healthy individuals (Figures 1a and 1b).
In FD tissue, PAS staining showed an abundant accumulation of polysaccharides, lipids, and glycosphin-golipids containing ceramide precursors that are represented by strong purple-red staining (Figure 1d).
Ceramide Accumulation Induces Abundant Morphological Changes Associated with Apoptosis in FD Colonocytes
H&E and PAS scoring analyses showed a large number of late-stage apoptotic cells in FD colon (Figures 1b and 1d, arrowheads). In situ TUNEL detection revealed massive early- and late-stage DNA fragmentation stained in dark blue (Figure 1h, arrowheads) compared to a few apoptotic epithelial cells found in normal colon (Figure 1g). The mitotic activity of stem cells at the base of the crypts produces a continuous supply of new cells, which progress to the mucosal surface where they mature before degeneration by apoptosis.
Colon mucosa from normal and FD subject stained for (
Most of the normal colon epithelial cells showing morphological evidence of apoptosis (8 ± 2.3% of the total) were localized to the mucosal surface (Figure 2). In contrast, whereas only 0.8 ± 0.2% of crypt cells were apoptotic in normal colon, a massive number of colonocytes (45 ± 4.3%) were apoptotic in crypts from FD colon (p<0.001) (Figures 1g and 1h, arrowheads, and Figure 2). These findings suggest that ceramide accumulation and triggering of apoptosis machinery may play an important role in FD cell destruction.
Active Caspase-3 Cleaves K18, Leading to Cell Death In Vivo
We then investigated whether ceramide-induced apoptosis in FD colonocytes showed involvement of caspase activation. Interestingly, analyses of serial colon sections shows that the active form of caspase-3 in colonocytes was mostly evident in colon crypt areas containing abundant reactivity for M30 (Figures 3b-d and 4c,d), an antibody that specifically recognizes a caspase cleavage site in K18 that is not detectable in native K18 of normal cells (Caulìn et al. 1997). In contrast, normal colon mucosa did not show any immunoreactivity evidence for caspase-3 (Figure 3a) or an active form of caspase-3 or M30 (not shown). These data suggest that the active form of caspase-3 determines the proteolytic cleavage of K18, leading to actin cytoskeletal disruption, chromatin condensation, and packaging of apoptotic bodies in ceramide-induced apoptosis in FD colonocytes.
GD3 Gangloside Co-localizes with Active Caspase-3 and Cleaved K18
Accumulation of GD3 ganglioside is an early event in Fas- and ceramide-induced apoptosis, responsible for apoptotic signal generation (De Maria et al. 1997). To determine the relative amount of Fas- and ceramide-induced pre-apoptosis in FD, we evaluated by immunohistochemistry the number of GD3-positive cells. FD colonocytes express negligible amounts of Fas, as shown by anti-Fas staining (Figure 3e). In contrast, strong immunoreactivity for GD3 ganglioside co-localized in serial sections with FD colonocytes that were positive for TUNEL, the active form of caspase-3, and M30 (Figures 3d,f and 4a–c), suggesting the rare involvement of the Fas/FasL system and the central role of GD3 in ceramide-induced apoptosis in FD.
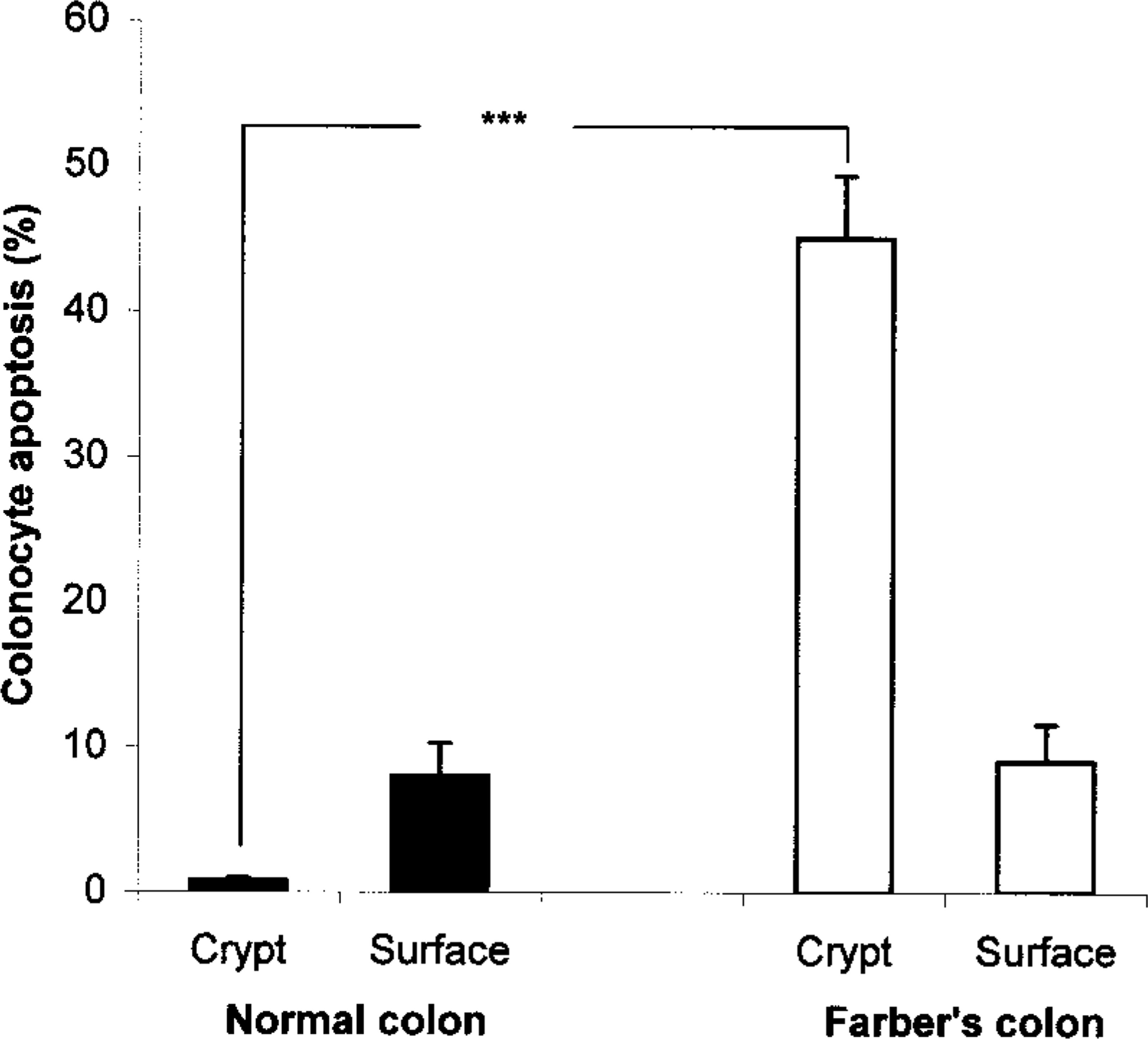
Percentage values of apoptotic cells in normal colon (▪) and FD colon (▪). Bars represent mean values ± SD. ∗∗∗p<0.001 for differences between the two groups.
Discussion
Although FD is genetically characterized, the pathogenic mechanisms determining cell tissue destruction are still not clear. Ceramide accumulation in FD cells has been classically regarded as the major pathogenic factor correlated with the severity of the disease (Qualman et al. 1987). This concept was suggested after the observation that ceramide is accumulated in the lysosomal fraction of FD skin fibroblasts showing membranous inclusions and cell destruction (Chen and Decker 1982). Furthermore, studies have reported that liver, spleen, and lymph nodes contained massive histiocytic infiltrates in association with elevated ceramide levels (Qualman et al. 1987; Fujiwaki et al. 1992; Zappatini-Tommasi et al. 1992; Moser 1997). However, although ceramide is believed to play a critical role in tissue damage, there is no evidence for a role of ceramide in the death pathway activated in FD.
Apoptosis has been proposed as a key pathogenic mechanism in several genetic, degenerative, and autoimmune diseases (Giordano et al. 1997; Stassi et al. 1997a,b,1999). The data presented here suggest that the ceramide pathway is responsible for triggering of apoptosis in FD epithelial cells. A substantial number of FD colonocytes undergo apoptosis in crypts. In fact, about 45% of FD colonocytes resident in the glands are apoptotic and show clear TUNEL reactivity. However, in normal subjects only a few mucosal surface colonocytes undergo spontaneous apoptosis, probably because of the continuous supply of new cells at the base of crypts. Moreover, we provide evidence that ceramide initiates the apoptotic program by generating GD3 neosynthesis, resulting in GD3 accumulation in FD cells.
The interaction of Fas and its ligand is a major apoptosis-inducing mechanism. Fas is a surface receptor expressed on many different cell types (Giordano et al. 1997; Stassi et al. 1997a,b,1999). We and others have demonstrated that triggering of the Fas molecule results in a cascade of metabolic events, including acidic sphingomyelinase activation and ceramide generation, which is a potent inducer of apoptosis (Cifone et al. 1995; De Maria et al. 1998).
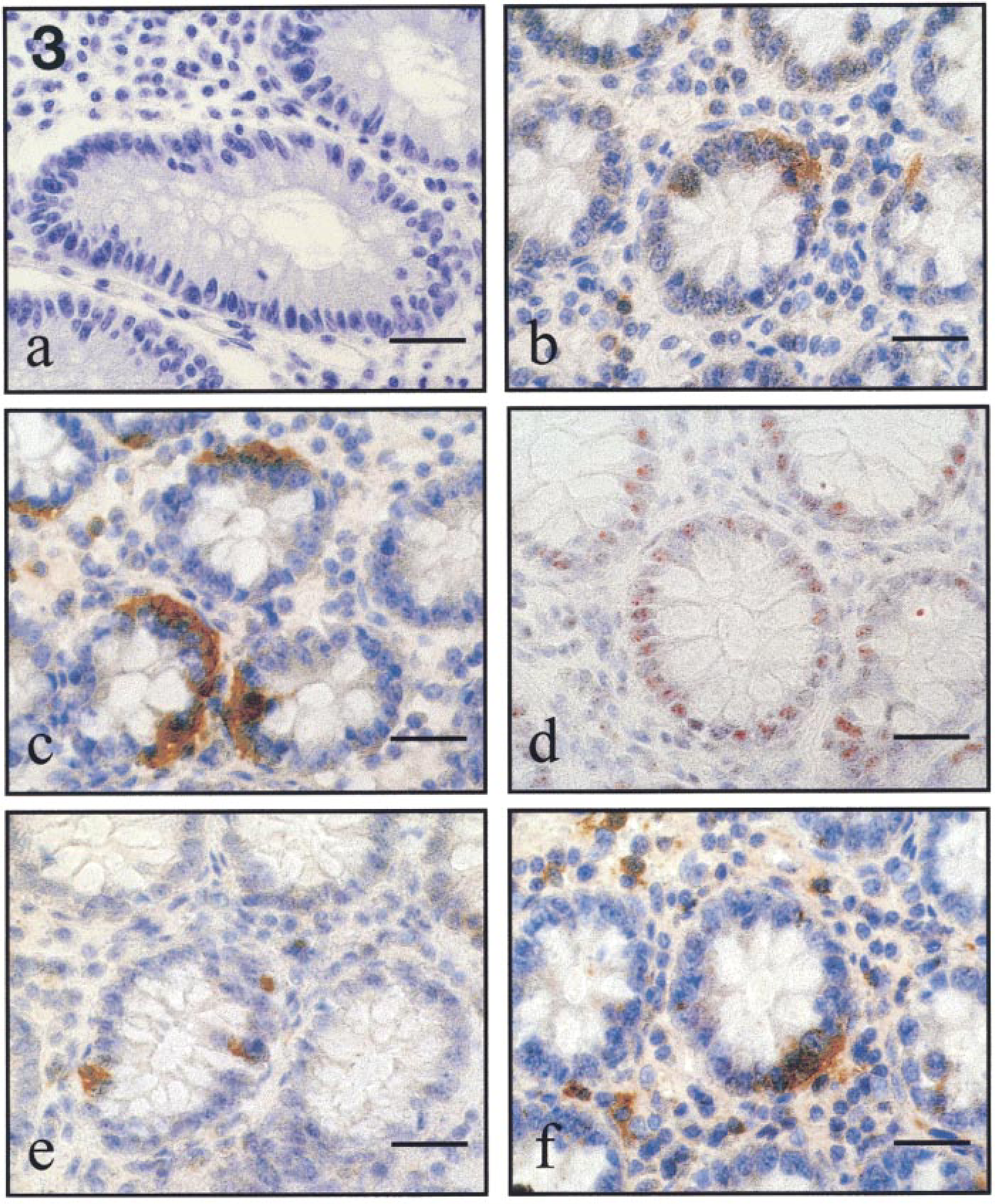
Representative experiment of normal and FD colon mucosa immunostaining. Normal subject: (
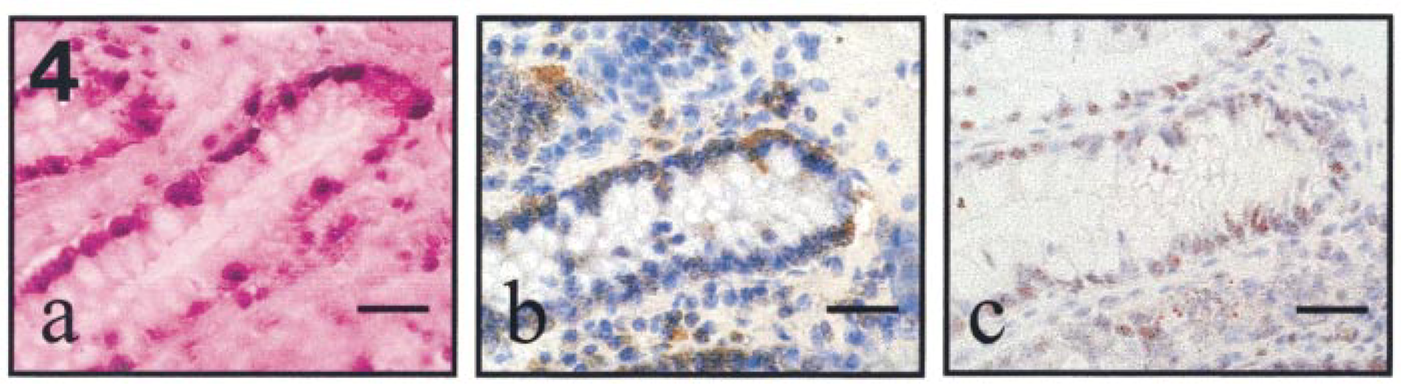
Co-localization between TUNEL-positive area and active form of caspase-3 or cleaved keratin 18 in FD colonic mucosa. (
Fas is rarely expressed in FD colonocytes, suggesting that is not involved in colon epithelial cell apoptosis. In contrast, GD3 ganglioside is abundantly accumulated, probably due to the deficient acid ceramidase activity. It is likely that in these cells GD3 acts as an inducer of caspase activation, promoting cleavage of a number of substrates that eventually results in cell death.
Recent studies support the idea that caspase-3 activation is responsible for fragmentation of keratins in apoptotic epithelial cells by specific cleavage (Caulìn et al. 1997). Accordingly, we found high levels of active form of caspase-3 co-localized with cleaved K18 in FD colonocytes.
In summary, GD3 synthesis induced by a high concentration of ceramide may alter mitochondrial function and promote cell destruction in a caspase-dependent manner, explaining the pathogenic mechanism of cell injury in FD colonocytes. Apoptosis along the entire colon epithelium down into the crypts may be a key pathogenetic factor in the persistent diarrhea and severe gastrointestinal lesions that occasionally occur in FD (Fujiwaki et al. 1992; Koga et al. 1992). It is likely that other epithelia of the digestive system, such as oropharyngeal and hepatic epithelia, are similarly affected, thus giving rise to the oropharyngeal lesions (Haraoka et al. 1997) and obstructive jaundice (Nowaczyk et al. 1996) frequently observed in FD.
Footnotes
Acknowledgements
Supported by Telethon-Italy Grant E.735 (to GS) and MURST 60% (to FF and GZ).
We are grateful to Drs S. Accomando (Pediatric Institute, University of Palermo), V. Marcianò, and L. Licata (Human Anatomy Section, University of Palermo) and G. Balsano (Institute of General Surgery, University of Palermo) for providing surgical specimens and helpful discussions and advice.