
Abstract
Deep brain stimulation of the subthalamic nucleus (STN-DBS) is a highly effective surgical treatment in patients with advanced Parkinson's disease (PD). Because the STN has been shown to represent an important relay station not only in motor basal ganglia circuits, the modification of brain areas also involved in nonmotor functioning can be expected by this intervention. To determine the impact of STN-DBS upon the regional cerebral metabolic rate of glucose (rCMRGlc), we performed positron emission tomography (PET) with 18-fluorodeoxyglucose (FDG) in eight patients with advanced PD before surgery as well as in the DBS on- and off-conditions 4 months after electrode implantation and in ten age-matched healthy controls. Before surgery, PD patients showed widespread bilateral reductions of cortical rCMRGlc versus controls but a hypermetabolic state in the left rostral cerebellum. In the STN-DBS on-condition, clusters of significantly increased rCMRGlc were found in both lower thalami reaching down to the midbrain area and remote from the stimulation site in the right frontal cortex, temporal cortex, and parietal cortex, whereas rCMRGlc significantly decreased in the left rostral cerebellum. Therefore, STN-DBS was found to suppress cerebellar hypermetabolism and to partly restore physiologic glucose consumption in limbic and associative projection territories of the basal ganglia. These data suggest an activating effect of DBS upon its target structures and confirm a central role of the STN in motor as well as associative, limbic, and cerebellar basal ganglia circuits.
Functional neurosurgery for the management of advanced Parkinson's disease (PD) has experienced a remarkable renaissance in the past decade. Moreover, the new technique of high-frequency stimulation is increasingly replacing lesional surgery. Deep brain stimulation of the subthalamic nucleus (STN-DBS) has been shown to result in the most effective improvement of motor function and is therefore an increasingly used treatment option in advanced PD patients (The Deep Brain Stimulation for Parkinson's Disease Study Group, 2001; Limousin et al., 1998). The principle aim of both lesional and DBS procedures is to restore the equilibrium of neuronal activity in the basal ganglia network and to release thalamocortical projections from overinhibition exerted by hyperactive basal ganglia output nuclei in PD (Benazzouz and Hallett, 2000). However, the antiparkinsonian DBS effects are adaptable with the adjustment of stimulation parameters and completely reversible after cessation of stimulation, which implicates unique features of the DBS technique associated with the administration of high frequency electrical stimuli.
The exact mechanism of antiparkinsonian DBS action is largely unknown as yet. Both the effect of stimulation on its neuronal target structure, such as inhibitory or excitatory, and the target tissue, such as cell bodies, fiber tracts, or both, are currently under debate. Moreover, the impact of DBS upon complex neuronal networks, which are involved in motor and nonmotor brain function and become integrated in the basal ganglia system, is not known. In the past, neuropsychologic side effects have been recognized to accompany the motor benefit in some patients with STN-DBS (Alegret et al., 2001; Ardouin et al., 1999; Bejjani et al., 1999; Jahanshahi et al., 2000; Krack et al., 2001; Saint-Cyr et al., 2000). Because the STN plays an important role not only in motor but also in limbic and associative basal ganglia circuits (Obeso et al., 1997; Rodriguez-Oroz et al., 2001), it is a matter of current debate to which extent changes of mood and behavior are related to stimulation-induced alterations of neuronal activity in extramotor basal ganglia networks. To address these questions, we measured the resting cerebral metabolic rate of glucose (rCMRGlc) with [18F]fluorodeoxy-glucose (FDG) and positron emission tomography (PET) in eight patients with advanced PD before surgery and 4 months after implantation in the STN stimulator off- and on-conditions. Measurements of regional cerebral glucose consumption with the use of FDG-PET provide a qualitative index of changes in neuronal activity (Sokoloff, 1977) and have been previously used to evaluate the effects of stereotactic interventions upon brain function in PD (Eidelberg et al., 1996; Fukuda et al., 2001a; Su et al., 2001).
SUBJECTS AND METHODS
Study subjects
After obtaining permission from the local ethics committee and written informed consent according to the declaration of Helsinki, eight patients with advanced PD (five men, three women, age [mean ± 61.8 ± 7.7 years, disease duration 14.9 ± 4.7 years) and ten healthy age-matched controls (six men, four women, age 62.6 ± 3.6 years) were included in the study. The mean age of both study groups was not significantly different (P = 0.81, unpaired t-test). All patients with PD were treated in our outpatient movement disorders clinic and included in the study in consecutive order. The clinical characteristics of patients with PD are shown in Table 1. PD was diagnosed according to the UK Parkinson's Disease Society Brain Bank criteria (Hughes et al., 1992) with the Hoehn and Yahr scale (HY) (Hoehn and Yahr, 1967) and the Unified Parkinson's Disease Rating Scale (UPDRS) (Fahn and Elton, 1987) as clinical measures of disease severity. All patients with PD had a sufficient drug response in a preoperative levodopa challenge and did not show any atypical clinical signs. STN-DBS was indicated in these individuals because of parkinsonian symptoms that were refractory to medication, such as severe levodopa-associated on/off fluctuations, peak dose dyskinesias, and severe resting hand tremor.
Patient characteristics, clinical data, and medication before surgery and at the time of FDG-PET scans under STN-DBS conditions
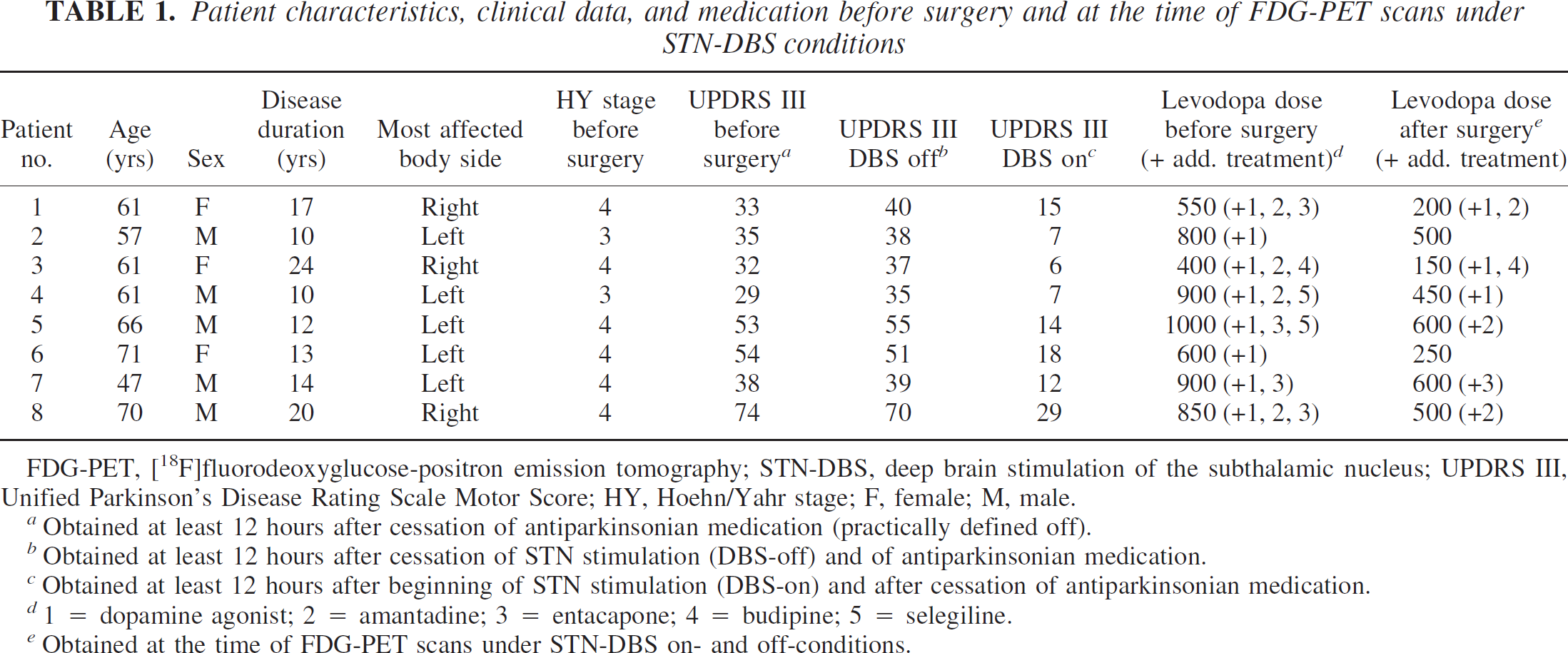
FDG-PET, [18F]fluorodeoxyglucose-positron emission tomography; STN-DBS, deep brain stimulation of the subthalamic nucleus; UPDRS III, Unified Parkinson's Disease Rating Scale Motor Score; HY, Hoehn/Yahr stage; F, female; M, male.
Obtained at least 12 hours after cessation of antiparkinsonian medication (practically defined off).
Obtained at least 12 hours after cessation of STN stimulation (DBS-off) and of antiparkinsonian medication.
Obtained at least 12 hours after beginning of STN stimulation (DBS-on) and after cessation of antiparkinsonian medication.
1 = dopamine agonist; 2 = amantadine; 3 = entacapone; 4 = budipine; 5 = selegiline.
Obtained at the time of FDG-PET scans under STN-DBS on- and off-conditions.
Surgical procedures
Bilateral STN electrodes (Medtronic model 3389, Medtronic, Minneapolis, MN, U.S.A.) and impulse generators (2 Itrel II, 6 Kinetra, Medtronic GmbH) were implanted as described previously (Hilker et al., 2002; Voges et al., 2002). In brief, the STN contour was outlined on axial T2-weighted MR-images. Additionally, the anterior-to-posterior commissural line (AC-PC) was defined on both T1- and T2-weighted MR-images, and the target, if necessary, was readjusted according to the data obtained from the Schaltenbrand brain atlas (2–3 mm behind the mid-commissural point [MCP], 3.7 mm below this point rectangular to the AC-PC line, and 12 mm lateral from the midline of the third ventricle). Finally, the tip of the electrode was positioned inside the dorsal third of the STN. Intraoperatively, the electrode position was verified using macrostimulation, stereotactic teleradiography, and repeated neurologic monitoring.
PET data acquisition
In patients with PD, three FDG-PET scans were performed: one was performed 2 to 4 weeks before DBS implantation, and two were performed in the stimulator on- and off-conditions 2 to 7 months after surgery (mean interval 3.8 ± 1.8 months) on consecutive days in random order. All PET scans were performed in the drug-off condition with the antiparkinsonian medication withdrawn for at least 12 hours before PET imaging. Likewise, the DBS devices were also switched on and off at least 12 hours before scanning. Individual STN stimulation parameters were identical with those programmed before the study (Table 2). One patient (Patient 2 in Table 2) had unusually high stimulation parameters (two adjacent active contacts with 4.5 V amplitudes, 90 μs pulse width, and 145 Hz frequency) without the occurrence of relevant side effects because suppression of severe resting hand tremor could only be reached using this setting, whereas other off-phase symptoms such as rigor, akinesia, and foot dystonia were already well controlled with lower DBS parameters.
STN stimulation parameters at the time of the PET study
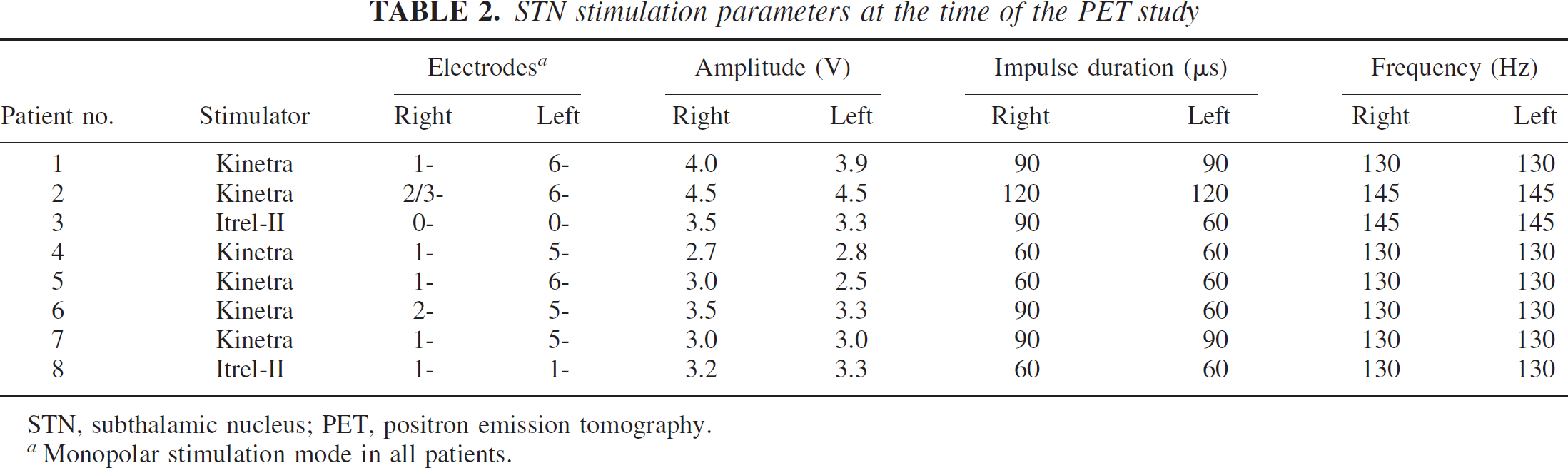
STN, subthalamic nucleus; PET, positron emission tomography.
Monopolar stimulation mode in all patients.
After overnight fasting, PET scans were performed on a 24-detector ring scanner (Ecat Exact HR, Siemens-CTI, Knoxville, TN, U.S.A.) in a 3D data acquisition mode (Bailey, 1992) with correction for scatter and attenuation. Images were reconstructed by 3D filtered back projection providing 47 contiguous transaxial image planes (slice thickness 3.1 mm) with reconstructed resolution of 5 mm (full width at half maximum) (Wienhard et al., 1994). The study subjects were positioned supine in the scanner with 3D laser alignment of the orbitomeatal line in a room with dimmed lighting and low background noise. Cerebral glucose metabolism was measured in resting state with eyes closed and ears unplugged after injection of 370 MBq FDG. Regional cerebral metabolic rates of glucose (rCMRGlc) were calculated pixel-by-pixel as described previously (Wienhard, 1992; Wienhard et al., 1985).
PET data analysis
Data processing was performed on a voxel-by-voxel basis using Statistical Parametric Mapping (SPM99, The Wellcome Department of Cognitive Neurology, London, UK) implemented in MATLAB (Mathworks, Sherborn, MA, U.S.A.). All scans were realigned and normalized into stereotactic space according to the atlas of Talairach and Tournoux (1988). Subsequently, images were scaled proportionally and smoothed using an isotropic 12-mm Gaussian kernel to increase signal-to-noise ratio and to compensate for interindividual anatomical variability. To separate effects of electrode implantation (microlesional effect) from direct STN stimulation effects in patients with PD, the data analysis was undertaken twice with the paired t-test option in SPM99 and a threshold P level of 0.01: first, by comparison of the preoperative state and the postoperative STN-DBS off-condition (determination of the microlesional effect), and second, by comparison of the postoperative STN-DBS off- and on-conditions (determination of direct STN stimulation effects). Clusters were accepted as significant if P values corrected for multiple comparisons were 0.01 or less. In addition, the preoperative and both postoperative rCMRGlc measurements of patients with PD were compared independently with FDG-PET scans of healthy controls by unpaired t-test statistics considering significant differences in brain areas with a corrected spatially restricted P value of 0.001 or less on the cluster level.
Neuropsychologic testing
All patients were examined with an extensive neuropsychologic test battery before surgery (levodopa-on condition) and at the time of the PET study (levodopa-on and stimulator-on conditions). Cognitive test batteries included the Mini-Mental-Status Examination (MMSE) (Folstein et al., 1975) and the Mattis Dementia Rating Scale (MDRS) (Mattis, 1988). In addition, the Beck Depression Inventory (BDI) (Beck, 1987) was administered. Specific tests for verbal short-term, working, and long-term memory were performed using the forward and reverse digit span subtests, a verbal selective reminding task with immediate and delayed recall (Memo-Test) (Schaaf et al., 1992), and the logical memory task of the Wechsler Memory Scale-Revised (WMS-R) (Wechsler, 1987). Moreover, tests for nonverbal long-term memory (delayed recall of the Rey-Osterrieth figure) (Spreen and Strauss, 1991), for executive functions (interference condition of the Stroop Test and verbal fluency tasks with the letters F, A, and S and the category “animals”) (Bäumler 1985; Spreen and Strauss, 1991), for visuoconstructive abilities (copy of the Rey-Osterrieth figure), and for speed of information processing (subtests 1 to 2 of the Stroop Test) were performed.
RESULTS
Clinical outcome
The UPDRS motor rating did not significantly differ between the preoperative state and the postoperative DBS-off condition (pre-DBS off 43.5 ± 15.5 vs. post-DBS off 45.6 ± 12.1). However, a significant clinical improvement was obvious after switching the STN stimulators on, resulting in a 70% reduction of the UPDRS motor score (DBS off 45.6 ± 12.1 vs. DBS on 13.5 ± 7.6, P < 0.001, paired t-test). The administered mean daily levodopa dose significantly decreased from 750 ± 208 mg before surgery to 406 ± 180 mg at time of the PET scans under STN-DBS (P < 0.01, paired t-test). The following transient side effects of STN stimulation were recorded in the first 6 weeks after surgery: paresthesias after turning the device on in 6/8 patients, DBS-induced choreiform dyskinesias in 5/8, severe depression in 2/8 (treatment with reboxetine 8 mg and amitryptiline 150 mg daily), and paranoid delusions in 1/8 (treatment with quetiapine 300 mg daily). At the time of the postoperative PET study, all stimulation side effects had been completely remitted. Individual data on clinical outcome are summarized in Table 1.
Determination of microlesional effects of STN electrode implantation
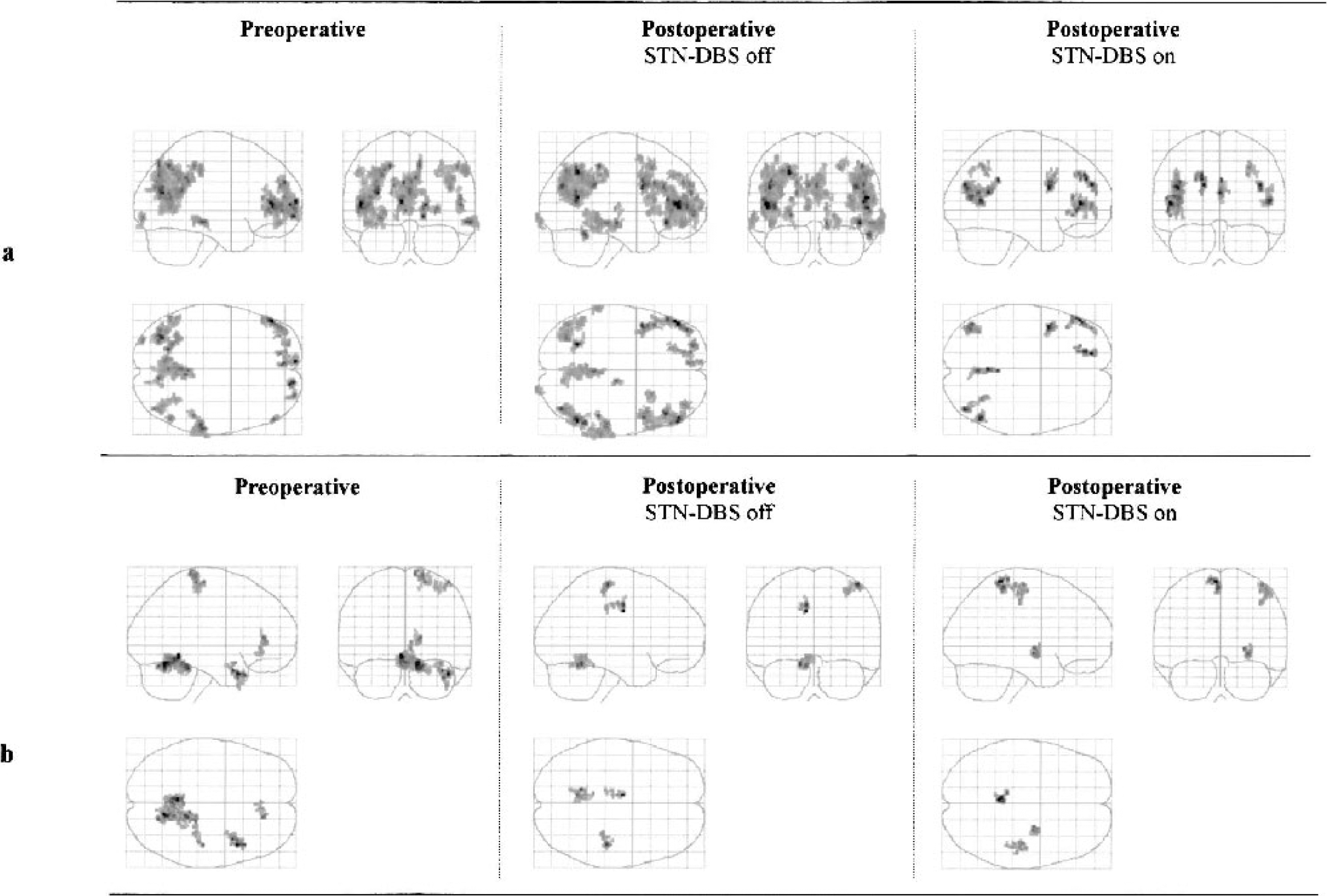
Regions with significantly decreased (A) and increased (B) rCMRGlc in eight patients with PD compared with ten age-matched healthy controls (P < 0.001 on cluster-level corrected for multiple comparisons, SPM99). Left panel, before surgery; middle panel, STN-DBS off-condition; right panel, STN-DBS on-condition. rCMRGlc, regional cerebral metabolic rate of glucose; PD, Parkinson's disease; STN-DBS, deep brain stimulation of the subthalamic nucleus.
Determination of direct STN stimulation effects
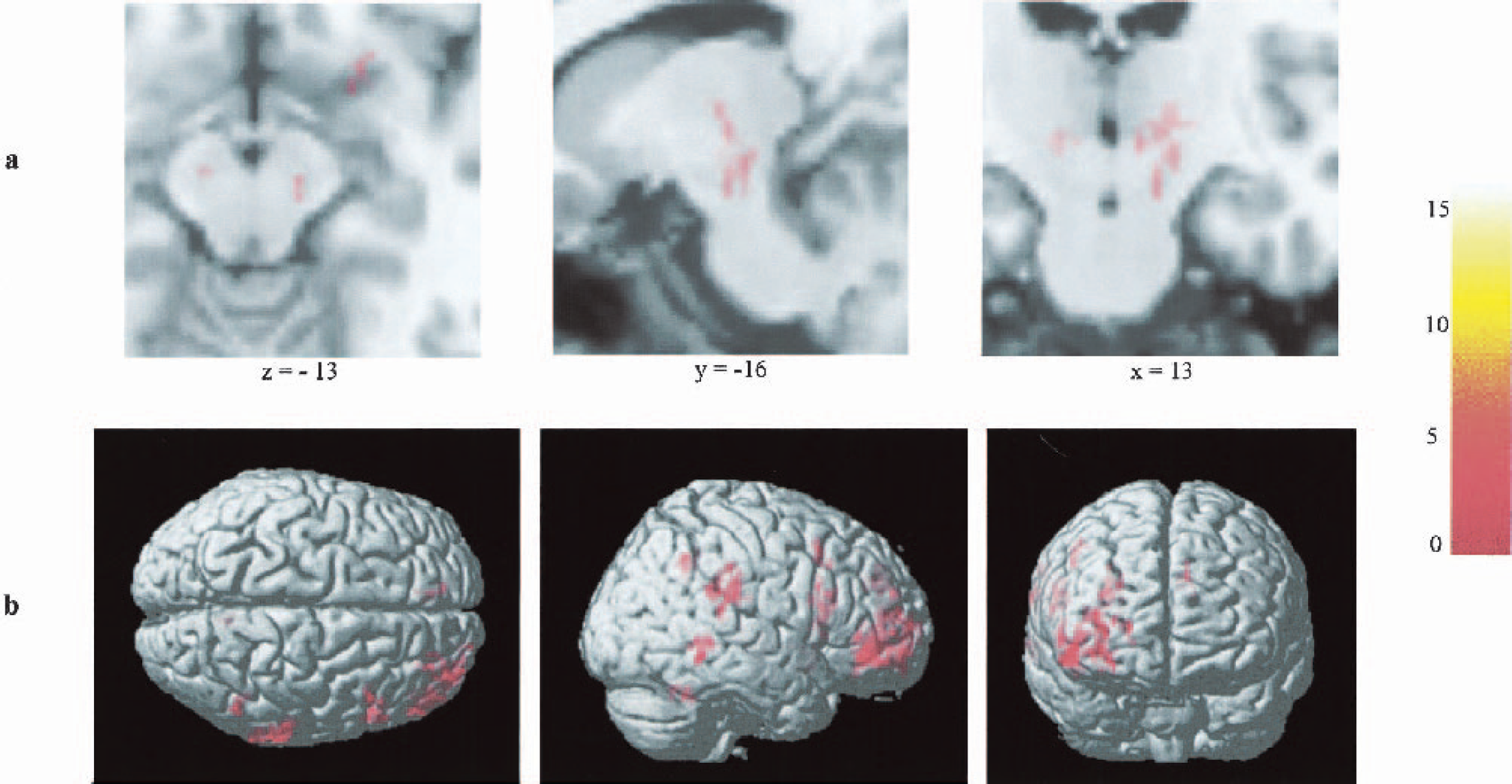
Regions with significantly increased rCMRGlc in the stimulator on- versus stimulator off-condition measured with SPM 99 (P < 0.01 on cluster-level corrected for multiple comparisons, color bar represents t-values). Transversal, sagittal, and coronal views (from left to right) in projection onto a standard MRI, depicted as brain slices (A, x/y/z coordinates according to Talairach atlas) and as 3D surface projection (B). rCMRGlc, regional cerebral metabolic rate of glucose; MRI, magnetic resonance imaging.
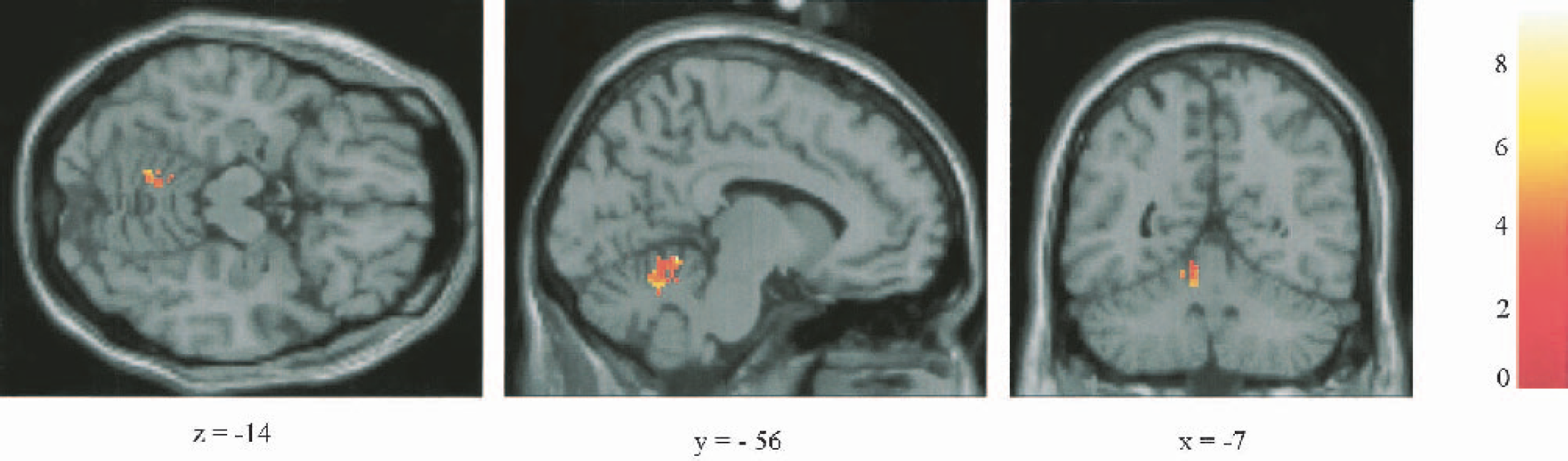
Significantly decreased rCMRGlc in the left anterior lobe of the rostral cerebellum in the stimulator on- versus stimulator off-condition measured with SPM 99 (P < 0.01 on cluster-level corrected for multiple comparisons, color bar represents t-values). Transversal, sagittal, and coronal views in projection onto brain slices of a standard MRI (x/y/z coordinates according to Talairach atlas). rCMRGlc, regional cerebral metabolic rate of glucose; MRI, magnetic resonance imaging.
Brain regions with significantly altered rCMRGlc in FDG-PET during bilateral STN-DBS ordered according to their peak z-scores
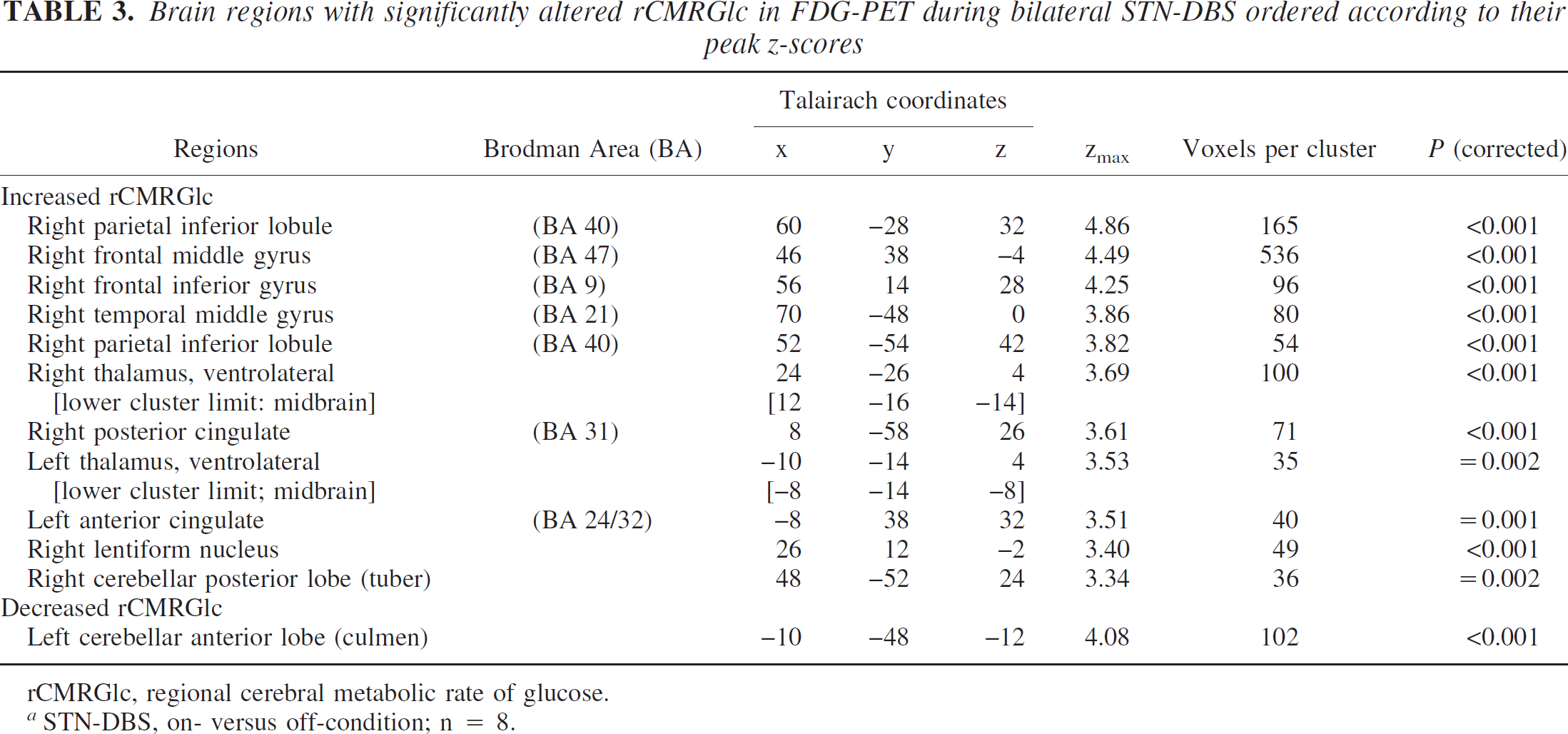
rCMRGlc, regional cerebral metabolic rate of glucose.
STN-DBS, on- versus off-condition; n = 8.
Neuropsychologic testing
Before surgery, patients with PD were not demented (MMSE 28.2 ± 1.2, Mattis score 141.0 ± 3.5) but scored below normal ranges in verbal short- and long-term memory, nonverbal long-term memory, letter fluency, and visuoconstructive abilities (Table 4). Clinically relevant depression was not observed, despite the slightly increased BDI score (10.1 ± 5.7). At the time of the postoperative PET study, the PD group significantly improved both in verbal and nonverbal longterm memory (59% in delayed recall of Memo-Test, P < 0.05; 44% in Rey-Osterrieth figure, P < 0.01), but verbal long-term memory was still slightly below the normal range (Table 4). No other significant differences between pre- and postoperative test performances were observed.
Neuropsychologic test performance of eight advanced patients with PD treated with STN-DBS before and at the time of the FDG-PET study after surgery
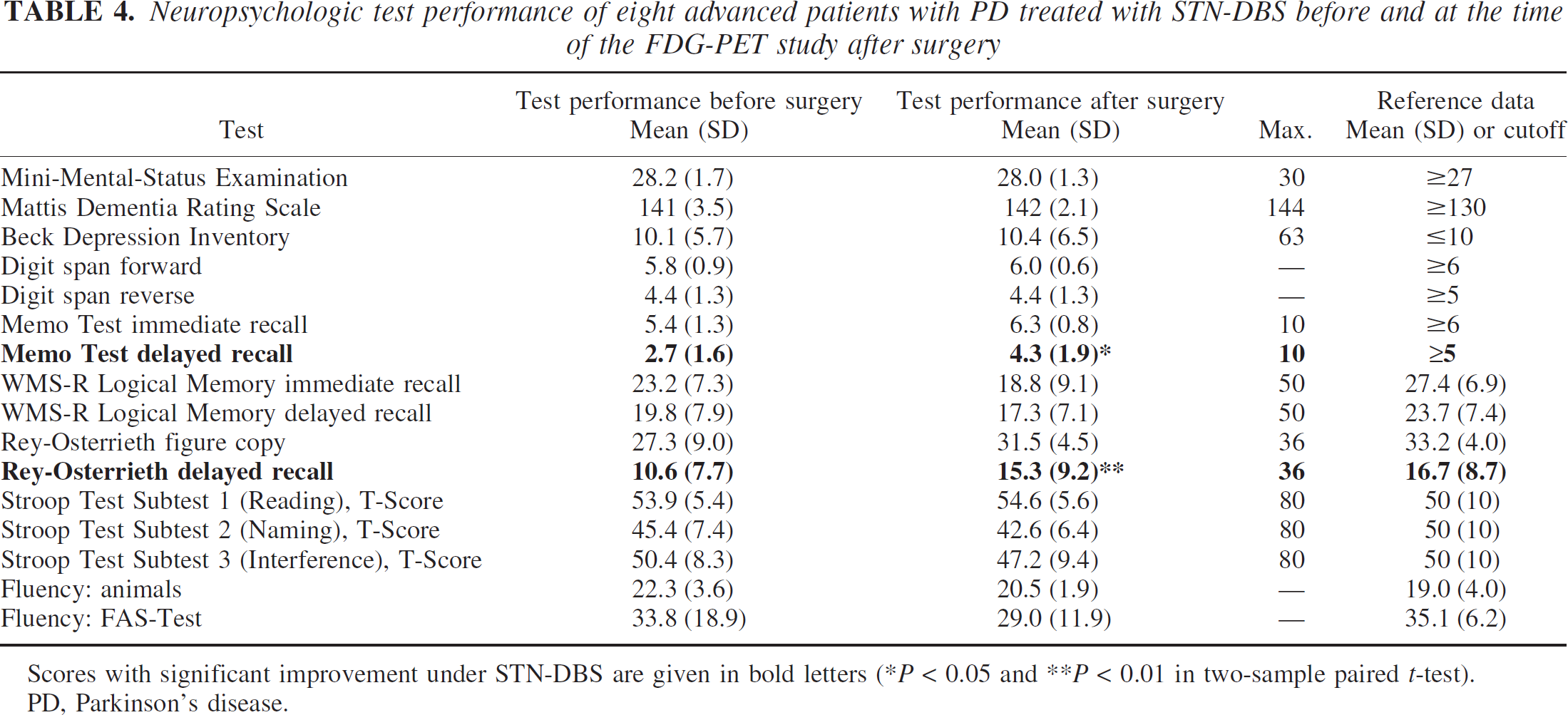
Scores with significant improvement under STN-DBS are given in bold letters (*P < 0.05 and **P < 0.01 in two-sample paired t-test). PD, Parkinson's disease.
DISCUSSION
In this study, we demonstrated that STN-DBS is able to restore glucose metabolism in widespread cortical areas of patients with PD within hours because regions with significantly decreased glucose use were distinctly restricted in the STN on-condition as compared with the preoperative and the DBS-off states. These metabolic changes were almost completely reversible after turning the devices off, indicating a functional and temporary phenomenon in line with the relapse of severe parkinsonism in the DBS off-condition. Our data indicate that impaired cortical FDG uptake in patients with advanced PD, which has been described by several groups in frontal and temporo-parietal lobes (Berding et al., 2001; Eidelberg et al., 1994; Hu et al., 2000; Turjanski and Brooks, 1997), should be considered as an at least partly reversible phenomenon presumably reflecting suppressed thalamocortical processing in the abnormal parkinsonian brain network. In contrast, we could not demonstrate any long-term microlesional effects of electrode implantation on the brains metabolic network, which became, however, obvious in patients with PD after unilateral subthalamotomy (Su et al., 2001).
Therefore, our study results indicate a characteristic stimulation effect fundamentally different from lesional procedures in the STN target. This hypothesis is particularly corroborated by our PET findings in subcortical brain regions showing small clusters of significantly increased FDG uptake during STN stimulation in the right lentiform nucleus and in both lower thalami reaching down to the midbrain DBS target area. These findings were unexpected because the rCMRGlc has been shown to be primarily determined by afferent synaptic activity (Auker et al., 1983), and, according to the common theory of DBS as an inhibitory intervention that blocks neuronal membranes, the reduction of STN hyperactivity by DBS should decrease the local PET signal in the pallidum and subsequently in the ventrolateral thalamus by reducing the STN excitatory glutamatergic output. However, previous PET studies on internal pallidum (GPi) stimulation did not show a DBS-induced decline of thalamic or pallidal rCMRGlc values (Eidelberg et al., 1996; Fukuda et al., 2001b) but demonstrated a significant increase of regional cerebral blood flow (rCBF) in the putamen ipsilateral to stimulation (Davis et al., 1997). During chronic STN stimulation, recent animal studies showed increased glutamate levels in the internal pallidum of normal rats (Windels et al., 2000) and elevated firing rates of GPi neurons in parkinsonian monkeys (Hashimoto et al., 2001). Thus, vascular and metabolic activation in the DBS target regions seems to occur, which suggests rather activating than inhibitory stimulation effects on neuronal cell bodies or axons leading to an increased output from the stimulation site onto afferent structures via ortho- or antidromic fiber conduction. Long-standing high-frequency activation of the STN target region might replace an abnormal phasic firing pattern in the pallidal outflow pathway, which was identified as a key feature of the parkinsonian state (Miller and DeLong, 1988), with a high-frequent, tonic activity state that no longer implies abnormal sense for the generation of parkinsonism (neuronal jamming) (Miller and DeLong, 1988; Vitek, 2002).
It is unknown as yet whether STN cell bodies or axons within the white matter encompassing the nucleus or even both are the primary target structures of STN-DBS. Ashby et al. suggested that DBS effects are most likely mediated by stimulation of large myelinated axons rather than cell bodies (Ashby, 2000; Ashby et al., 1999). A recently published study by Voges and colleagues found the best improvement of PD motor symptoms per administered energy unit in electrode contacts, which were located outside the STN (Voges et al., 2002). The authors concluded that blocking or activation of large fiber connections arising in the STN or running nearby is more likely the active principle of DBS than electrical interference with cell bodies inside the nucleus. Important fiber systems surrounding the STN are the pallidothalamic bundle and the zona incerta. Animal tracer studies revealed that the zona incerta located closely above the STN represents a broad system of descending and ascending fibers connecting the subthalamic region with other basal ganglia nuclei and widespread areas of the neocortex (Nicolelis et al., 1992; Power et al., 1999; Ricardo, 1981). We propose that our PET findings of stimulation-induced metabolic activation in both midbrain areas reaching bilaterally up to the basal ventrolateral thalami and to the right lentiform nucleus most likely reflect neuronal jamming of these fiber bundles and their projections sites, which hold a central position within information processing through the basal ganglia network.
The marked neocortical metabolic activation under STN-DBS indicates a divergent influence of the STN region upon widespread cortical projection areas. The basal ganglia system can be subdivided into three main territories, namely the sensorimotor projecting to the motor cortices, the associative projecting to the prefrontal, and the limbic projecting to the ACC and the medial orbito-frontal cortex (Parent, 1990). Our PET finding of a strongly activated right-sided glucose metabolism in the frontal limbic and associative territory implicates STN functioning as an important relay not only in motor but also in basal ganglia loops, which process cognitive and emotional information. This hypothesis agrees with the identification of motor, associative, and limbic territories in an orderly sequence from the dorsolateral to the ventrolateral STN (Alexander et al., 1990). These subunits of the nucleus receive input from the prefrontal cortex and send back projections to the motor, associative, and limbic areas of the frontal lobe (Alexander et al., 1990; Canteras et al., 1990; Feger et al., 1994; Maurice et al., 1998). Further anatomic studies proved reciprocal pathways between the basal ganglia, midline limbic structures, the brainstem, and cortical frontal and parietotemporal association cortices (Carmichael and Price, 1995). These connections provide a morphologic base for the strong rCMRGlc increases in the right inferior parietal cortex and the adjacent posterior middle temporal gyrus, which were found in our study subjects with PD under STN stimulation.
Our data indicate that STN-DBS interacts with basal ganglia loops, which are involved in the regulation of cognition, mood, and behavior. In patients recovering from severe depression, a recent PET study found enhanced glucose metabolism in the right DLPFC and in the inferior parietal, dorsal anterior, and posterior cingulate cortex, whereas a decreased radiotracer uptake was detected in the ventral subgenual cingulate and in the insula (Mayberg et al., 1999). A reverse pattern of rCBF alteration was found during provocation of acute sadness. Therefore, the authors proposed a reciprocal interaction between ventral limbic and dorsal cortical compartments in mood shifts in either direction. It is interesting to note that the dorsal neocortical areas that get activated in patients resolving from depression are widely congruent with those revealing restored glucose metabolism in the STN-DBS on-condition in our study subjects with PD.
Thus our PET findings agree with the clinical observation that PD-related depression tends to improve after STN stimulation (Ardouin et al., 1999; Volkmann et al., 2001). However, transient euphoria and manic psychosis or anhedonia and depression were also reported to compromise the motor benefit of STN stimulation in the first weeks after device implantation (Bejjani et al., 1999; Doshi et al., 2002; Krack et al., 2001, 2002; Volkmann et al., 2001). In our small PD study sample, severe neuropsychiatric side effects were observed in three out of eight patients within the first 6 weeks after surgery. These subjects did not differ from other study patients in terms of preoperative glucose metabolism and medication changes after surgery. Neuropsychiatric side effects remitted completely in all affected individuals, leading to unchanged BDI scores compared with baseline at the time of the PET study. Therefore, STN-DBS can be considered a safe procedure with respect to psychiatric side effects in the long-term course. However, our PET data suggest that STN-DBS induces marked activity shifts in limbic basal ganglia territories within hours, which complies with accelerated normalization of neuronal activity in hypometabolic cortical regions. We propose that this process remains subclinical in the majority of patients, but might also become transiently apparent in others, resulting in a clinical spectrum of affective symptoms ranging from depression to mania.
Our study subjects with PD showed unchanged executive function 4 months after surgery but showed significantly improved verbal and nonverbal long-term memory according to metabolic activation in memory relevant areas, such as the right DLPFC (Fletcher and Henson, 2001). Other groups found conflicting results in detailed neuropsychologic testing ranging from absent relevant effects of STN-DBS on memory and executive functions (Ardouin et al., 1999) to improved working memory and deteriorated Stroop interference tasks (Jahanshahi et al., 2000; Pillon et al., 2000). The latter finding might corroborate the prediction of impaired flexible behavior after stereotaxic surgery in Parkinson's disease (Marsden and Obeso, 1994).
We did not find an increase of rCMRGlc in the supplementary motor area (SMA) under STN-DBS, although significant movement-related increases of blood flow had been reported in this area (Ceballos-Baumann et al., 1999; Limousin et al., 1997). This finding is presumably explained by our study design, which investigated patients at rest in the PET scanner without the execution of a motor activation paradigm. Another possible explanation might also be the use of a rather high statistical threshold in our study, which might lead to false negative results because of the miss of minor metabolic changes in the SMA. Moreover, the strong lateralization of stimulation-induced activations to the right hemisphere despite bilaterally implanted devices is a striking finding of our study in which the pathophysiologic meaning remains speculative. One possible explanation for this phenomenon might be the asymmetric affection of the hemispheres in our PD study population (predominantly affected body side in five subjects on the left and three subjects on the right side), possibly leading to a stronger restoration of thalamocortical processing by STN stimulation ipsilateral to the main focus of dopamine depletion. Another explanation would be that the interaction of STN-DBS with limbic and associative circuits is reflected in right-lateralized metabolic changes in FDG-PET because specialization of the right hemisphere for mood and behavior regulation and lateralization of mood symptoms in pathologic conditions were repeatedly described previously (Carson et al., 2000; Grafman et al., 1986; Sackeim et al., 1982).
Finally, our data demonstrate a clear influence of STN-DBS upon the activity of cerebellar neurons. The rostral cluster of decreased rCMRGlc was largely congruent with the hyper-metabolic area that was obvious in patients with PD before surgery and in the stimulation off-condition but no longer under effective STN-DBS. Thus cerebellar hypermetabolism seems to be a characteristic feature of the parkinsonian off-state, which disappears with STN stimulation. Previous FDG-PET studies also identified the pons and cerebellum as hypermetabolic regions in PD (Eidelberg et al., 1994; Ghaemi et al., 2002). The pathophysiologic meaning of this finding is currently unclear, but it was proposed that the cerebellum might be involved in the generation of parkinsonian tremor (Brooks et al., 1992). In our study subjects, a marked suppression of parkinsonian tremor in the DBS on-condition with more than 50% reduction of the UPDRS tremor items occurred in four out of eight patients. One male subject needed unusually high stimulation amplitudes for sufficient tremor control (4.5 V on both sides, Table 2). In this patient, FDG-PET showed a bilateral rCMRGlc increase in the posterior cingulate as well as a marked cerebellar decrease in the rostral vermis and the right hemisphere, corroborating the concept of cerebellar involvement in parkinsonian tremor generation.
Two previous PET studies demonstrated decreased cerebellar blood flow with successful PD tremor suppression by DBS of the thalamic ventral intermediate nucleus (VIM) (Deiber et al., 1993; Parker et al., 1992). We therefore suggest that both STN- and VIM-DBS interact either in a direct or indirect manner with cerebellothalamic-cortical pathways that mediate PD tremor, such as by deactivating the rostral cerebellum via an antidromic effect in dentate-thalamic fibers. This hypothesis is in keeping with the clinical observation that the efficacy of STN-DBS in the treatment of drug-resistant PD tremor compares with that of VIM stimulation. Moreover, the prefrontal and parietal association cortices, which have been found to be activated by STN-DBS, send out projections to the ipsilateral cerebellum via the pontine nuclei (Haines et al., 1997), which might explain the right-lateralized activation of glucose consumption in the posterior cerebellar lobe.
In conclusion, the results of this PET study demonstrate that STN-DBS is able to widely restore glucose metabolism in the right frontal cortex and temporoparietal cortex and to suppress rostral cerebellar hypermetabolism in patients with PD within hours. Thus STN-DBS seems to interact with widespread cortical and cerebellar pathways in brain regions spatially remote from the stimulation targets. These alterations of neuronal resting energy metabolism were almost completely reversible after turning the devices off, indicating a functional and temporary phenomenon in line with the relapse of severe parkinsonism in the DBS off-condition. Our finding of increased subcortical glucose metabolism, in particular within the subthalamic and midbrain region, suggests an activating effect of DBS upon its target structures unlike lesional procedures such as subthalamotomy. Moreover, our data also indicate a central role of the STN in motor as well as associative, limbic, and cerebellar basal ganglia circuits. From a clinical point of view, we propose to consider long-term mood stabilization but also transient neuropsychiatric side effects as intrinsic characteristics of DBS in the STN target, which must be thoroughly taken into account in the management of patients with PD treated with this intervention.