
Abstract
Purinergic P2X7 receptors may affect neuronal cell death through their ability to regulate the processing and release of interleukin-1β (IL-1β), a key mediator in neurodegeneration. The authors tested the hypothesis that ATP, acting at P2X7 receptors, contributes to experimentally induced neuronal death in rodents in vivo. Deletion of P2X7 receptors (P2X7 knockout mice) did not affect cell death induced by temporary cerebral ischemia, which was reduced by treatment with IL-1 receptor antagonist (IL-1RA). Treatment of mice with P2X antagonists did not affect ischemic or excitotoxic cell death, suggesting that P2X7 receptors are not primary mediators of experimentally induced neuronal death.
Purinergic receptors have important actions in the CNS (Ralevic and Burnstock, 1998). The P2X7 receptor subtype, expressed primarily in microglia and macrophages, is activated by ATP and regulates the release of the proinflammatory cytokine interleukin-1 (IL-1) (Ferrari et al., 1997; Sanz and Di Virgilio, 2000; Le Feuvre et al., 2002a,b). Interleukin-1, which exists as IL-1α and IL-1β, is a key mediator of experimentally induced neurodegeneration (Allan and Rothwell, 2001; Rothwell, 1999; Touzani et al., 2002). Thus, inhibition of IL-1 action (by IL-1 receptor antagonist [IL-1RA]) or deletion of genes for IL-1α and IL-1β dramatically reduces neuronal injury induced by cerebral ischemia and excitotoxicity (Boutin et al., 2001, and see Allan and Rothwell, 2001). Recently, we have shown that activation of P2X7 receptors by ATP regulates cleavage and release of both IL-1α and IL-1β in primary cultures of microglia and macrophages (Brough et al., 2002; Le Feuvre et al., 2002b). P2X7 receptors may thus play an important role in neuronal death and could be an important therapeutic target.
The objective of the present study was to determine whether activation of P2X7 receptors contributes to experimentally induced neurodegeneration in rodents in vivo. Our preliminary data (see review by Le Feuvre et al., 2002b) showed that the absence of the P2X7 receptor did not significantly affect neuronal damage after temporary occlusion of the middle cerebral artery (MCA) or exposure to the excitotoxin cis,-2,4-methanoglutamate (Mglu). The present study investigates the role of IL-1 in the ischemic damage in P2X7 knockout (KO) mice and the effects of P2X antagonists on ischemic- and excitotoxic-induced damage.
MATERIALS AND METHODS
Experiments were performed on male, adult (25 to 30 g) wild-type (WT; B6D2[C57BL6xDBA/2] F1) and P2X7 receptor gene KO mice (P2X7 KO) littermates (Solle et al., 2001; breeding pairs donated by Pfizer, Groton, CT, U.S.A.). Absence of the P2X7 receptor was verified by polymerase chain reaction analysis (Le Feuvre et al., 2002a). Male C57/SV129 mice (25 to 30 g; Charles River, Margate, U.K.) were used to investigate the effects of P2X7 antagonists and agonists on MCA-occlusion (MCAO)–induced damage. Animals were maintained in a 12-h light/dark cycle at 22°C, and all experiments were performed in accordance with U.K. legislation under the 1986 Animals (Scientific Procedures) Act.
Focal cerebral ischemia was induced by right MCAO using the intraluminal filament technique (Hata et al., 1998). Briefly, a filament (7/0 suture; Ethicon, Edinburgh, U.K.), thickened to 180 μm for 1.5 mm at its tip, was inserted into the common carotid artery and advanced up to the origin of the MCA via the internal carotid artery. Reduction in cerebral blood flow (approximately 70%) was confirmed by laser Doppler flowmetry (Moor Instruments, Axminster, U.K.), and body temperature was monitored and maintained at 36.5°C to 37.5°C. Reperfusion of the tissue was achieved by withdrawing the filament (under brief anesthesia) after 30 or 60 minutes. Cerebral vasculature, checked by perfusion of animals with paraformaldehyde containing 10% india ink, appeared identical between WT and P2X7 KO mice.
Administration of vehicle (0.9% sterile saline), interleukin-1 receptor antagonist (IL-1RA, 5 μg; Amgen, Thousand Oaks, CA, U.S.A.), the P2X agonist benzoylbenzoyl-ATP (BzATP, 30 mmol/L) or P2X antagonists oxidized ATP (o-ATP, 3 mmol/L) or pyridoxalphosphate-6-azophenyl-2′,4′-disulphonic acid (PPADS, 30 μmol/L) in a volume of 1 μL, was performed for a 5-minute period via indwelling right lateral ventricular cannulae (implanted under anesthesia 1 week before MCAO), immediately before and again 30 minutes after MCAO.
Excitotoxic lesions were induced by infusion (0.2 μL/min) of the N-methyl-d-aspartate (NMDA) receptor agonist cis-2,4-methanoglutamate (Mglu, 0.5 μL of 10 mmol/L; Tocris Cookson, St. Albans, U.K.) directly into the striatum (0.7 mm anterior, 1.8 mm lateral, and 3.0 mm dorsoventrally relative to bregma; Franklin and Paxinos, 1997) of halothane-anesthetized WT and P2X7 KO mice. Immediately before MGlu infusion, mice were infused (loaded in the same syringe as MGlu) with either 0.5 μL of the P2X antagonists PPADS (10 mmol/L) or [N,O-bis(5-isoquinolinesulphonyl)-N-methyl-l-tyrosyl]-4-phenylpiperazine (KN-62, mmol/L; Sigma Chemicals, Poole, U.K.) or their respective vehicle (0.9% sterile saline or 3.5% dimethyl sulfoxide [DMSO] respectively).
Twenty-four hours after MCAO or excitotoxic damage (when damage is maximal), mice were killed and brains were removed and frozen in isopentane. Viable tissue was identified on frozen brain sections (20 μm) stained with cresyl violet, and lesion volume was quantified by automated image analysis. For MCAO damage, infarct areas were corrected for edema as described previously (Boutin et al., 2001). Data are presented as mean + SD, and statistical differences were tested using ANOVA with Tukey post hoc test for multiple groups, or Student's t-tests for two groups.
RESULTS
Occlusion of the MCA for 60 minutes in vehicle-treated P2X7 KO mice resulted in reproducible infarcts in the striatum and cortex, which were similar in size to those previously observed in noncannulated animals (Le Feuvre et al., 2002b). Infarct volumes were significantly reduced (58%, P < 0.01) in animals treated with IL-1RA (vehicle-treated mice, 34 + 18 mm3 vs. IL-1RA-treated mice, 14 + 10 mm3; n = 7 in both groups; Fig. 1). Both groups had similar body temperature (37.7 + 0.6°C and 37.3 + 0.6°C vehicle- and IL-1RA-treated mice, respectively) and reductions in cerebral blood flow on MCAO (74.1 + 10.2% and 69 + 7.3% reductions on MCAO, vehicle and IL-1RA, respectively).
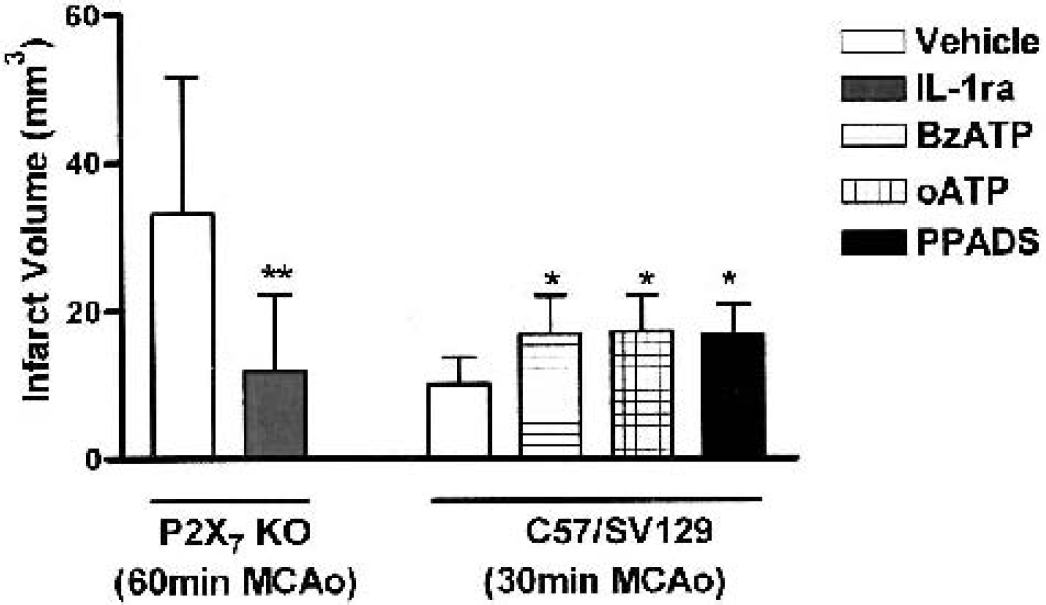
Effect of interleukin-1 receptor antagonist (IL-1RA) on total lesion volume after temporary (60-minute) middle cerebral artery occlusion (MCAO) in wild-type and P2X7 knockout (KO) mice was measured 24 h after MCAO (n = 7 per group). The effects of injections (1 μL intracerebroventricularly) of the P2X receptor agonist BzATP (30 μmol/L) and the antagonists oATP (3 mmol/L) and PPADS (30 μmol/L) on lesion volumes after 30-minute reperfusion MCAO in C57/SV129 mice are also shown. Values are mean + SD. **P < 0.01 versus vehicle (Student's t-test) for the IL-1RAdata, and *P < 0.05 versus vehicle, ANOVA for the P2X antagonist/agonist study. BzATP, benzoylbenzoyl-ATP; o-ATP, oxidized ATP; PPADS, pyridoxalphosphate-6-azophenyl-2′,4′-disulphonic acid.
Middle cerebral artery occlusion for 30 minutes in vehicle-treated (C57/SV129) mice produced compact striatal lesions (10 + 4 mm3) as shown in Fig. 1. Treatment of mice with the P2X receptor agonist (BzATP, 30 mmol/L) or the receptor antagonists, oATP (3 mmol/L) and PPADS (30 μmol/L), produced small increases in lesion volumes compared with vehicle-treated controls (16 + 5 mm3, 17 + 6 mm3, and 17 + 4 mm3, respectively; n = 7 to 9, P < 0.05; Fig. 1).
Administration of 0.9% saline or DMSO (3.5%, the vehicle for KN-62 experiments) with MGlu in P2X7 WT mice induced excitotoxic lesions of the striatum that spread into the adjacent cortex (lesion volumes: 24 + 13 and 22 + 6 mm3, respectively; Fig. 2). Infusion of either PPADS (10 mmol/L) or KN-62 (2 Mmol/L) with MGlu in WT mice had no effect on lesion volume compared with their appropriate vehicle-treated controls (24 + 9 mm3 and 27 + 9 mm3, respectively; Fig. 2). Treatment of P2X7 KO with saline and MGlu produced similar lesions to those observed in WT mice (32 + 3mm3). Infusion of PPADS with MGlu did not affect lesion volumes in P2X7 KO mice (33 + 5 mm3; Fig. 2).
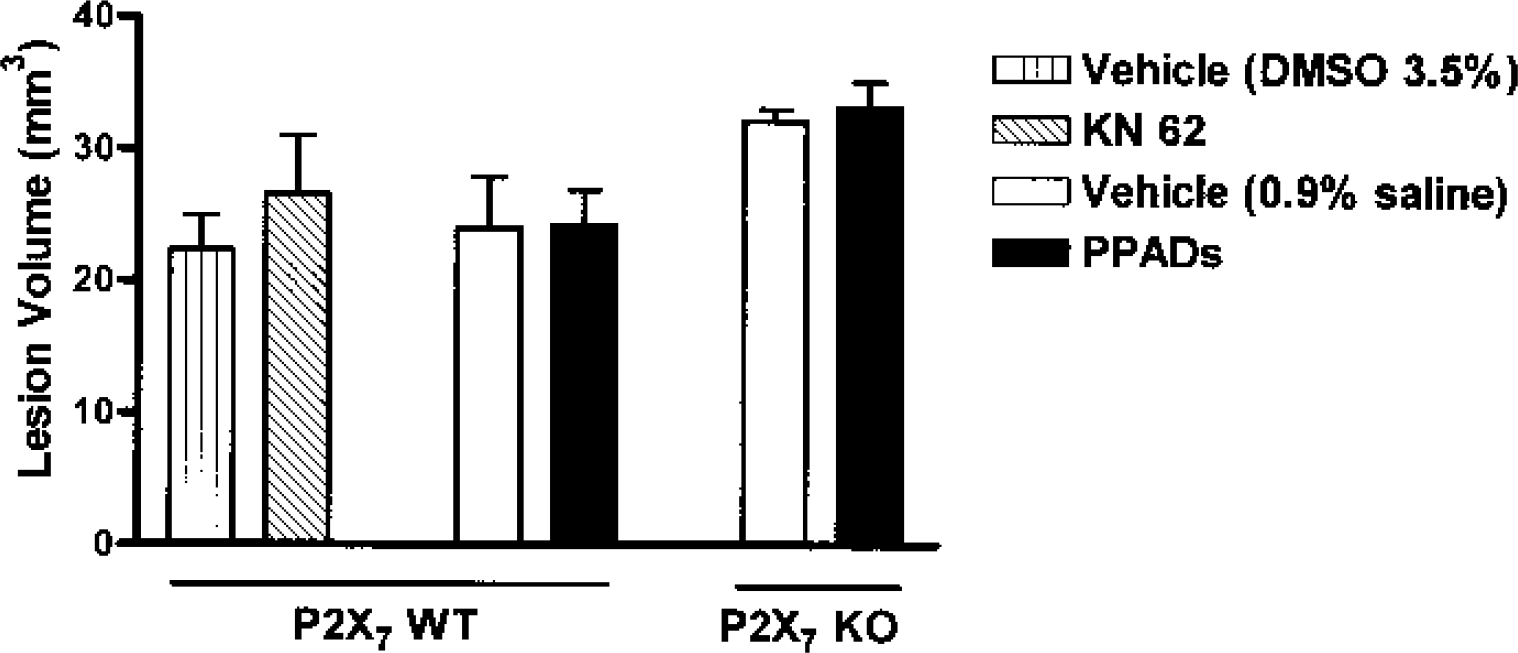
The effects of infusion (0.5 μL) of P2X receptor antagonists, KN-62 (2 mmol/L) and PPADS (10 mmol/L) or their respective vehicle (3.5% dimethyl sulfoxide [DMSO] and 0.9% sterile saline, respectively) immediately before infusion of cis,-2,4-methanoglutamate (0.5 μL of 10 mmol/L) on lesion volumes are shown. Values are mean + SD (n = 7 to 11 for PPADS and vehicle groups, n = 4 for KN-62, and n = 6 for DMSO vehicle). No significant differences were observed between groups. KN-62, [N,O-bis(5-isoquinolinesulphonyl)-N-methyl-l-tyrosyl]-4-phenylpiperazine; PPADS, pyridoxalphosphate-6-azophenyl-2′,4′-disulphonic acid.
DISCUSSION
There is now considerable in vitro evidence to suggest that extracellular ATP is involved in inflammatory responses and neuronal death induced by ischemic stress (Di Virgilio et al., 1998; Nieber et al., 1999; Cavaliere et al., 2002). Whereas extracellular ATP is normally present at low concentrations in the brain (Dubyak and el-Moatassim, 1993), levels can increase significantly under inflammatory conditions in vivo or in vitro (Sikora et al., 1999; Ferrari et al., 1997; Lazarowski et al., 2000), and ATP may be released from dying cells, in response to tissue trauma (e.g., ischemia–hypoxia [Nieber et al., 1999]), or co-released with glutamate from presynaptic terminals (Sperlágh and Vizi, 1996). Increased P2X7 receptor messenger RNA expression has been observed around the penumbra of ischemic damage (Collo et al., 1997). This, together with the well-documented effects of P2X7 receptor activation on IL-1 processing and release (Ferrari et al., 1997; Sanz and Di Virgilio, 2000), suggests that the P2X7 receptor could regulate IL-1 release and thus neuronal death in vivo. The role, however, of ATP and P2X7 receptors in neurodegeneration in vivo has not been widely investigated.
In our previous studies (Le Feuvre et al., 2002b), we showed that deletion of the P2X7 receptor (P2X7 KO mice) does not affect neuronal death induced by 30- to 60-minute MCAO, permanent MCAO, or excitotoxic injury, compared with responses in WT littermates. These studies suggested that, although IL-1β release is absent in microglia isolated from P2X7 KO mice challenged with ATP (Brough et al., 2002), it is possible that IL-1 is secreted from microglia via alternative pathways activated by cerebral ischemia or excitotoxic challenges.
Despite the important role of IL-1 β in neurodegenerative conditions (Rothwell, 1999), IL-1β KO mice do not exhibit reduced ischemic brain damage, whereas deletion of both IL-1α and IL-1β causes a substantial reduction in damage (Boutin et al., 2001). These data suggest that IL-1α can compensate for IL-1 β in its absence. P2X7 KO mice could therefore exhibit “normal” responses to ischemia. However, ATP activation of the P2X7 receptor stimulates release of IL-1α as well as IL-1β in vitro, and neither form of IL-1 is released from P2X7 KO macrophages in response to ATP (LeFeuvre et al., 2002a). The data presented here (Fig. 1) show that IL-1RA dramatically reduces infarct volume in P2X7 KO mice, suggesting that IL-1 is still involved in the damage in P2X7 KO mice and that factors other than ATP may be responsible for IL-1 processing or release in response to cerebral ischemia. Sixty-minute MCAO was chosen for this study because the strain of mice used shows little overt signs of behavioral disturbances with this protocol, and because it results in a lesion that is optimal in size for observing neuroprotection.
The effects of P2X antagonists and agonists on ischemic and excitotoxic damage were investigated. We tested the effects of BzATP, a stable analog of ATP, which is a potent agonist of the P2X7 receptor, and the P2X receptor antagonist's oATP and PPADS on ischemic damage induced by 30-minute MCAO in C57/SV129 mice (Fig. 1). Thirty-minute MCAO was chosen because this time point was well characterized in this mouse strain, known to produce discrete lesions with no behavioral disturbances, and with an increase in damage as a possible outcome. Both the antagonists and the agonist produced small increases in neuronal damage induced by ischemia.
The effects of higher doses of the P2X receptor antagonist PPADS and KN62 (thought to have high specificity at P2X7 receptors) were investigated on excitotoxic cell death induced by infusion of MGlu and the appropriate vehicle. Although the doses chosen were high (PPADS, 5 nmol) these were comparable to those used in other in vivo studies (Gourine et al, 2002; Kittner et al, 2001), and a high dose was considered necessary to block the direct actions of the glutamate agonist. Preincubation with the antagonists was not possible without prolonged or repeated anesthesia, which would influence damage and is associated with increased mortality. Furthermore, IL-1β expression in microglia is not increased 1 to 4 h after excitotoxic injury in the rat (Pearson et al, 1999). Neither of the P2X antagonists affected excitotoxic damage (Fig. 2). Although these studies suggest that P2X antagonists do not reduce cell death in vivo, the selectivity of these agents is still not fully established, and full dose responses and time courses were not investigated. Neurotoxic actions of oATP on cerebellar granule neurones in vitro have been reported previously by our group (Craighead et al., 2001). Other reports, however, have not reported toxic actions in a number of cell types (Murgia et al., 1993), and no overt cell death was observed when the antagonist was injected directly into the brain of control animals (data not shown). The small stimulatory action of the P2X agonist, BzATP, may also reflect an ability of this compound to stimulate tumor necrosis factor-α release from microglia (Hide et al., 2000).
Previous studies in vitro have described neuroprotective actions of P2X receptor antagonists, particularly suramin and reactive basilin blue. These are both nonselective antagonists, however, and may exert additional actions at NMDA receptors (Peoples and Li, 1998). Selective antagonists of P2X7 receptor function, GSK883 and GSK671 (provided by GlaxoSmithKline, Harlow, U.K.) (which have 50% inhibitor concentration effects of 735 nmol and 1 μmol/L, respectively, in the P2X7 YO-PRO assay [Michel et al., 1999] and confirmed by electrophysiologic methods [Chessell et al., 1998] without affecting P2X4) also failed to reduce excitotoxic damage in vivo (data not shown).
In conclusion, although ATP can stimulate processing and release of IL-1β in vitro, through actions at P2X7 receptors, these receptors do not seem necessary for IL-1 actions in cerebral ischemia. Although these studies suggest that P2X7 receptors do not contribute significantly to ischemia or excitotoxic damage in rodents in vivo, further experiments are required when more selective compounds become available.