
Abstract
Spreading depression (SD) is considered to play a role in pathologic conditions of humans such as in the evolution of ischemic brain injury and migraine aura. Because many studies have demonstrated spreading hypoperfusion in patients with migraine and persistent hypoperfusion in nonprimate animal models of SD, these changes in cerebral blood flow (CBF) were regarded as an epiphenomenon of SD. However, there is no direct evidence of the occurrence of SD in primates. The authors attempted to elicit SD by applying 3.3 mol/L potassium chloride to the cerebral cortex of nine male cynomolgus monkeys. The CBF was monitored by positron emission tomography in five animals. Propagated direct-current shifts were found by the two neighboring microelectrodes only in one animal. The direct-current wave propagated at a speed of 4 mm/min and its amplitude was 20 mV, being consistent with the SD findings. Except in one animal with 6 SD episodes, SD waves were recorded infrequently at the rostral site (none in three animals, once in three, and twice in two). Focal hyperemia accompanied SD. Neither spreading hypoperfusion nor persistent hypoperfusion was found. These unique features of SD in primates raise a doubt as to whether the role of SD in nonprimate animals is the same as that in stroke and migraine in humans.
Keywords
Cortical spreading depression (SD), which was first reported by Leao (1944), is characterized by reversible depression of cortical electrical activity in the brain that spreads like a wave at a speed of 2 to 5 mm per minute. Spreading depression can be induced by a variety of experimental stimuli such as electrical, chemical, and mechanical stimuli. Spreading depression has been observed in rats (Lauritzen et al., 1982; Iijima et al., 1992; Gardner-Medwin et al., 1994; Hasegawa et al., 1995; Takano et al., 1996), cats (Piper et al., 1991; Saito et al., 1995; Kuge et al., 2000), and a patient with severe head injury (Mayevsky et al., 1996).
There is strong experimental evidence that repetitive pathologic SDs in the periinfarct border zone contribute to the evolution of infarcts in rat and cat models (Gill et al., 1992; Iijima et al., 1992; Chen et al., 1993; Nedergaard and Hansen, 1993; Back et al., 1994; Hossmann, 1994; Takano et al., 1996; Ohta et al., 2001). Because ischemia-related SDs are considered to exacerbate the preexisting energy depletion in the periinfarct zone (Hossmann, 1994; Takano et al., 1996), they are regarded as a target of pharmacologic intervention. Although most neuroprotective agents that block the occurrence of SDs in rodents (Gill et al., 1992; Iijima et al., 1992; Chen et al., 1993) work well during brain ischemia in such small animals (del-Zoppo et al., 1997), the majority of potential neuroprotective therapies modifying ischemic cell death cascades did not have beneficial effects in patients who had had a stroke.
Transient cortical hyperemia followed by persistent hypoperfusion was reported to be an epiphenomenon of SD in rat- and cat-SD models (Lauritzen et al., 1982; Piper et al., 1991; Kuge et al., 2000). Spreading depression has been speculated to underlie migraine visual aura based on indirect evidence from human neuroimaging studies (Olesen et al., 1981; Lauritzen et al., 1983; Lauritzen and Olesen, 1984; Woods et al., 1994; Hadjikhani et al., 2001). Because spreading hypoperfusion was demonstrated during a migraine attack in these previous studies (Olesen et al., 1981; Lauritzen et al., 1983; Lauritzen and Olesen, 1984; Woods et al., 1994; Hadjikhani et al., 2001), it is regarded as an attendant change in cerebral blood flow (CBF) on SD in patients with migraine attacks. However, the hypothesis that aura underlain by SD phenomenon might be the trigger for migraine, particularly being pivotal to pain, was indicated to be implausible from the clinical point of view (Goadsby, 2001). Recently, Ebersberger et al. (2001) did not see any proof of neurogenic inflammation in migraine headache. Furthermore, there are no reports providing direct evidence that SD, characterized by the reversible depression of cortical electrical activity, can be elicited in primates, although a few studies suggested the occurrence of SD by measuring the changes in cortical fluorescence (Strong et al., 2000), the kinetics of resolution of extracellular potassium activity (Branston et al., 1977), and recording the slow potential of lissencephalic cortex in squirrel monkey (Robert, 1970).
The participation of SD in various neuronal disorders in humans has been a matter of speculation. In the current study, we attempted to clarify whether SD can be evoked in primates. In addition, we examined the attendant changes in the CBF during the provocation of SD using positron emission tomography (PET).
MATERIALS AND METHODS
Animal preparation
All procedures in this study were approved by our Institutional Animal Research Committee and were performed in accordance with the standards published by the National Research Council (Guide for the Care and Use of Laboratory Animals). In compliance with these standards, every effort was made to ensure that the animals were free from pain or discomfort. The principal investigator and primate handling staff were present for all procedures. The monkeys were individually in cages larger than 63 cm (width) × 76 cm (height) × 76 cm (depth) and maintained on a 12-hour light/dark cycle (lights on at 8:00 a.m.). All animals were neurologically normal and had no evidence of infections or inflammation immediately before the experiments.
Nine adult, male cynomolgus monkeys (nos. 1 through 9) were anesthetized with pentobarbital (50 mg/kg, intraperitoneally), followed by tracheal intubation. Then, the anesthetic state was maintained with N2O/O2 (70%:30%) gas mixture inhalation under artificial ventilation. Any other anesthetic agents were not added during the experimental period. The rectal temperature was monitored continuously with a rectal probe and maintained at approximately 36°C with a heating pad. An arterial catheter was used for continuous monitoring of heart rate and arterial pressure.
Electrophysiologic measurement
The anesthetized animal was mounted on a custom-made stereotaxic instrument in a prone position, and the head was restrained using teeth-and ear-bars. After the frontoparietal cranium was exposed by a midsagittal incision, three to four small burr holes were made in the left parietal skull bone and the dura was carefully excised; one burr hole was used for KCl application and two to three were used for direct current (DC) potential recordings. Of these, two burr holes were made rostral to the hole for KCl application in all animals, for recording the DC potential through microelectrodes inserted into the cortical surface. Another hole for recording the DC potential was made caudal to the hole for KCl application, in two animals (nos. 4 and 5). These burr holes were made along a line at 7-mm intervals. We also recorded the DC shift in the right parietal skull in one monkey (no. 4), although it has been reported that SD waves do not spread to the contralateral hemisphere (Marshall, 1959; Hasegawa et al., 1995).
The DC potential was monitored with an amplifier (Iso-DAM8; World Precision Instruments, Sarasota, FL, U.S.A.) by microelectrodes that had been inserted into the cortex to a depth of 1 mm. Physiologic saline followed by KCl solution was applied to the cortex through one of the burr holes. The KCl solution was applied up to 3 times at intervals of 10 minutes until a SD was elicited by the KCl. The concentration of KCl of 3.3 mol/L was chosen, because lower concentrations such as 0.15 mol/L and 1 mol/L did not induce SD in our preliminary study with monkeys (data not shown).
At 120 minutes after KCl application, the brain tissues of animals that did not undergo the PET study described in the next section were perfused with cold saline and the animals were killed by exsanguination under pentobarbital anesthesia. The brain was quickly removed, and several samples were dissected out and stored at −80°C for future analyses. A sagittal slice through the insertion sites of the two microelectrodes was obtained from one monkey (no. 7) to measure the length of the cortical surface between the two microelectrodes.
Positron emission tomography study
Positron emission tomography was performed with a multislice PET scanner (ECAT EXACT HR/47; Siemens/CTI, Knoxville, TN, U.S.A.) (Wienhard et al., 1994), which provides 47 tomographic images at 3.1-mm intervals per frame. The spatial resolution at the center of the field of view was 3.7 mm in-plane at full width at half maximum and 4.1 mm axially.
Five of the nine animals (nos. 1 to 5) underwent the PET study simultaneously with the recording of SD waves. The animal was placed in the field of view in the prone position and the head was restrained by teeth-and ear-bars. In the PET experiments, the brain position was standardized with the aid of laser beams. This enabled us to correlate PET images with the outer orbital point and the meatus auditorius. Positron emission tomography images were then correlated with the locations of stimulation and DC potential measurement. Blood gas and blood sugar analyses were performed in blood samples obtained from the femoral artery before and 120 minutes after the KCl application. The CBF was measured by PET with an intravenous bolus injection of 15O-labeled water. Dynamic PET scanning (0 to 2 minutes) was initiated at the time of the 15O-H2O injection. A baseline CBF measurement (Pre) was made before the topical application of 3.3 mol/L of KCl solution in all animals. In three animals (nos. 1 to 3), the CBF was also measured before the administration of physiologic saline to confirm that the mean hemispheric CBF before administration of KCl solution was in the same range as that before the administration of physiologic saline. In the animals with SD occurrence, the CBF was measured within 10 minutes after the first SD was elicited (CBF1), followed by 3 to 5 PET scans at intervals of approximately 10 to 20 minutes. If a SD was not elicited, the CBF1 was measured 10 minutes after the third KCl application, followed by 3 PET scans that were obtained in the same manner as in the other animals that had SDs.
After the PET study was completed, the brain tissues were perfused with cold saline and the animals were killed in the same manner as those that did not undergo the PET study. Several samples obtained from the brain were stored at −80°C for future study.
Data analysis
The PET images were reconstructed according to a standard filtered back-projection procedure using a Hanning filter. Each activity image was globally normalized by relative ratio to regional counts of the cerebellar cortex ipsilateral to the site of KCl application. Time courses of CBF changes for every frame in either transaxial or coronal images were ascertained by the visual inspection by two of the authors (Y.K. and N.E.), whether there were any areas of persistent hypoperfusion as compared with the contralateral hemisphere, and also spreading hypoperfusion was observed following primary hyperperfusion after SD. In addition, a total of 14 circular regions of interest were positioned in each hemisphere symmetrically on three transaxial brain slices as shown in Fig. 1. Considering the spatial resolution of the PET apparatus in the present study, the diameter of regions of interest (ROIs) was 8 mm (0.5 cm2 in area), twice as much as the value of full width at half maximum. Because of various problems as pointed by Heiss et al. (1994), that is, with repeated withdrawal of blood during prolonged experimental periods, partial volume effects, and excluding the effects associated with variations in levels of anesthesia, we used the asymmetry index, which was defined as the ratio of the value of regional CBF in the ROI in the hemisphere ipsilateral to the site of KCl application to that in the contralateral homologous ROI.
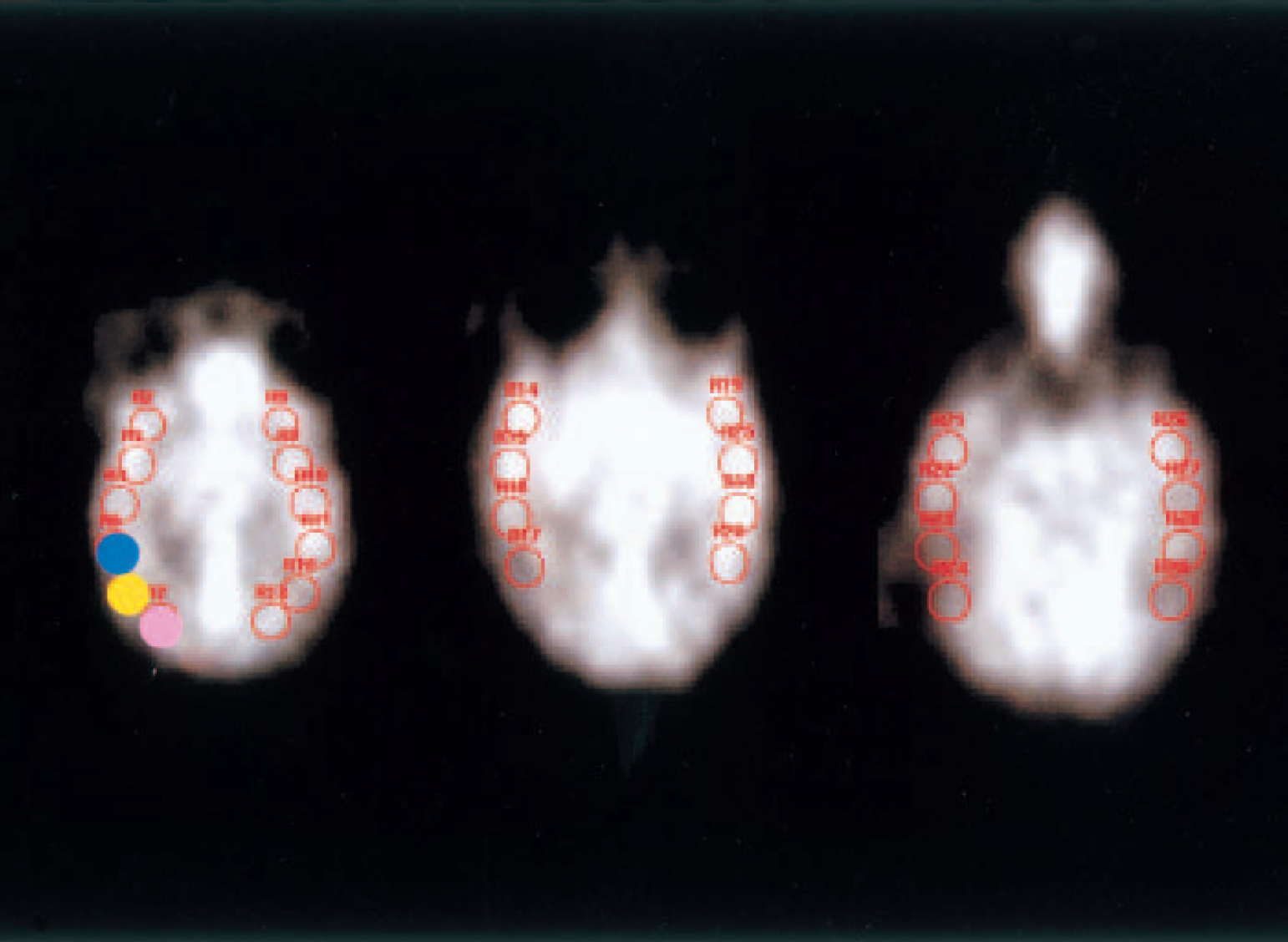
Examples of regions of interest (ROIs; each ROI had an area of 0.5 cm2) placed on three transaxial images. The slices are approximately 2.5 cm, 3.2 cm, and 4.2 cm, respectively, from the top of the precentral gyrus in the field of view. Fourteen circular ROIs were placed on the cerebral cortex ipsilateral to the site of KCl application, and 14 additional ROIs were placed symmetrically in the contralateral hemisphere. The ROI rostrally adjacent to the site of KCl application is indicated with a blue circle; the one caudally adjacent to the KCl application site is shown in pink; and the KCl application site is indicated with a yellow circle.
Data are expressed as the mean + SD, unless otherwise noted. The significance of differences in the sequential change of each physiologic parameter was assessed using repeated-measures one-way analysis of variance and Fischer's PLSD post hoc test. A two-tailed P value less than 0.05 was considered to be significant.
RESULTS
Electrophysiologic study
During the experimental period, physiologic parameters of the monkeys including body temperature, heart rate, mean arterial pressure, blood sugar, and blood gas levels did not change significantly (Table 1).
Serial changes in physiologic variables
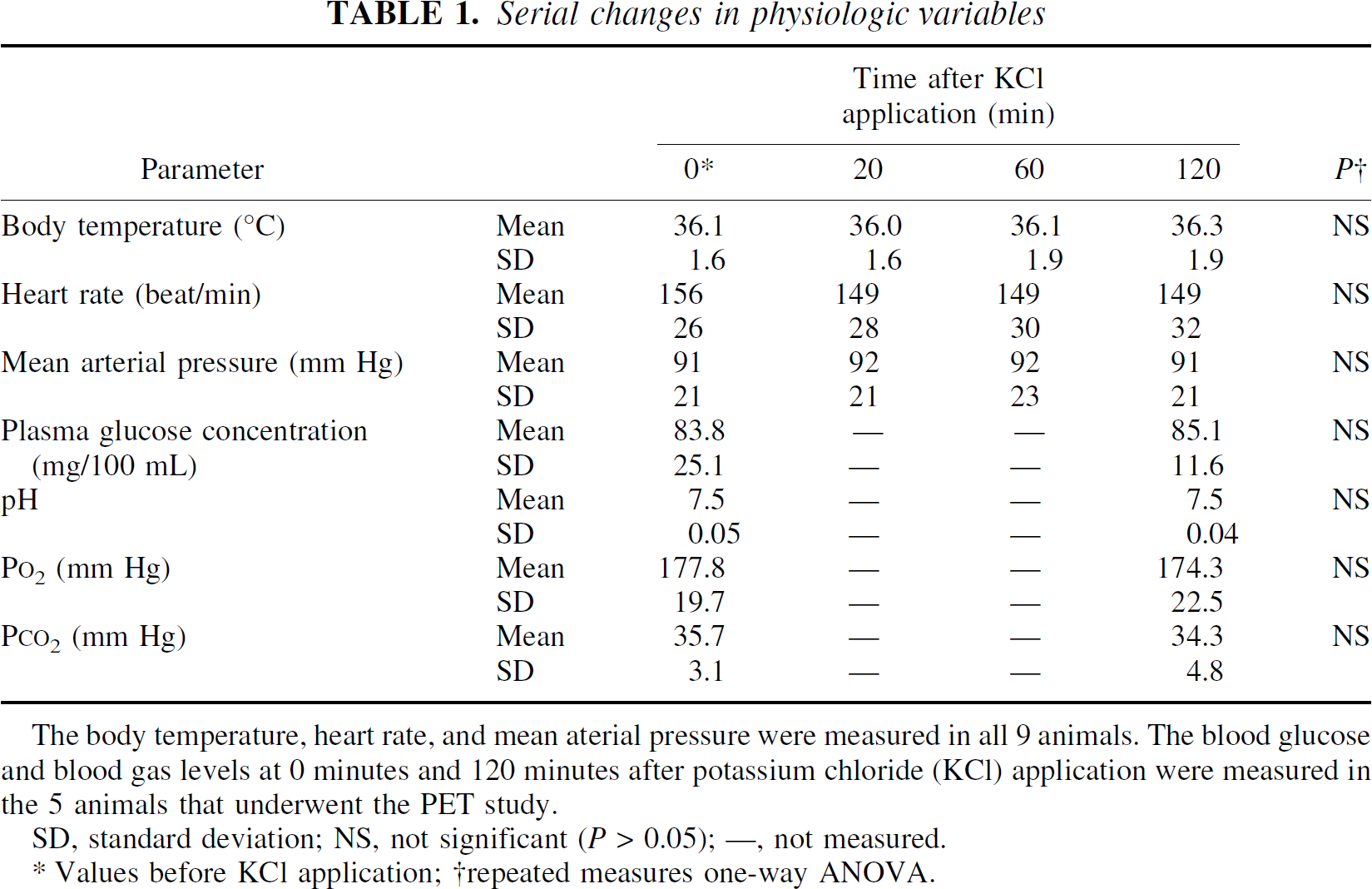
The body temperature, heart rate, and mean aterial pressure were measured in all 9 animals. The blood glucose and blood gas levels at 0 minutes and 120 minutes after potassium chloride (KCl) application were measured in the 5 animals that underwent the PET study.
SD, standard deviation; NS, not significant (P > 0.05); —, not measured.
Values before KCl application;
repeated measures one-way ANOVA.
The number and location of DC shifts elicited by the KCl in each monkey are listed in Table 2. Shifts of DC potential were recorded through the microelectrode just rostral to the site of chemical stimulation in only five of the nine monkeys: a single episode was recorded in three monkeys (nos. 3, 6, and 7), two episodes in one monkey (no. 2), and six episodes in one monkey (no. 9). Of the two animals with the caudal hole, animal 5 had eight episodes in the site caudal to the site of chemical stimulation, but no DC shift at the rostral sites. The other animal (no. 4) had two DC shifts, each of which was rostrally adjacent to the site of chemical stimulation, and six episodes at the caudal site, but no episodes at the contralateral site.
Number of direct-current shifts detected by the microelectrodes at each site
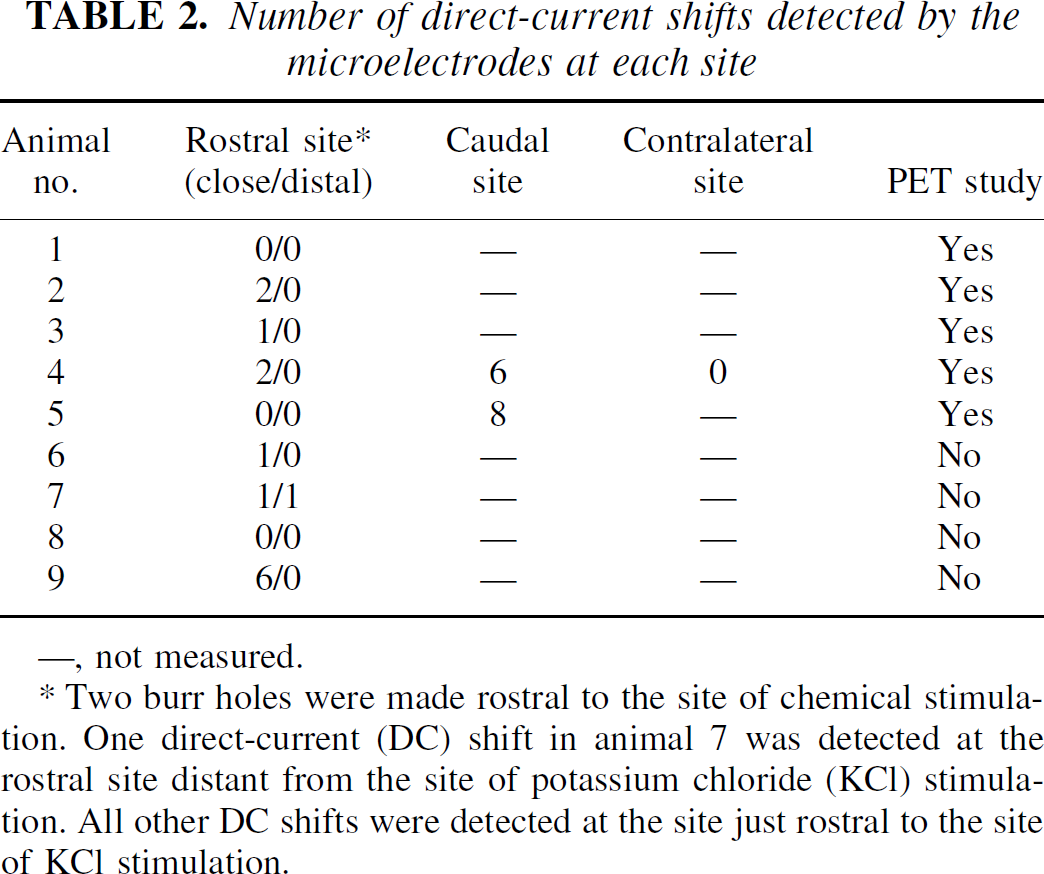
—, not measured.
Two burr holes were made rostral to the site of chemical stimulation. One direct-current (DC) shift in animal 7 was detected at the rostral site distant from the site of potassium chloride (KCl) stimulation. All other DC shifts were detected at the site just rostral to the site of KCl stimulation.
A propagated DC shift was found on the recordings of DC potentials in two neighboring microelectrodes 7 mm apart in only one animal (no. 7). In this monkey, the length of the cortical convolution between the 2 microelectrodes was 48 mm. The DC shift spread between the two microelectrodes in 12 minutes (Fig. 2A), resulting in a propagation speed of 4 mm per minute. The amplitude was 20 mV. These values are consistent with the SD findings reported in previous studies (Leao, 1944; Marshall, 1959). The representative image of repetitive SD waves in animal 9 is shown in Fig. 2B. Propagation of SD waves was not detected in the rostral hole distant from the site of KCl application.
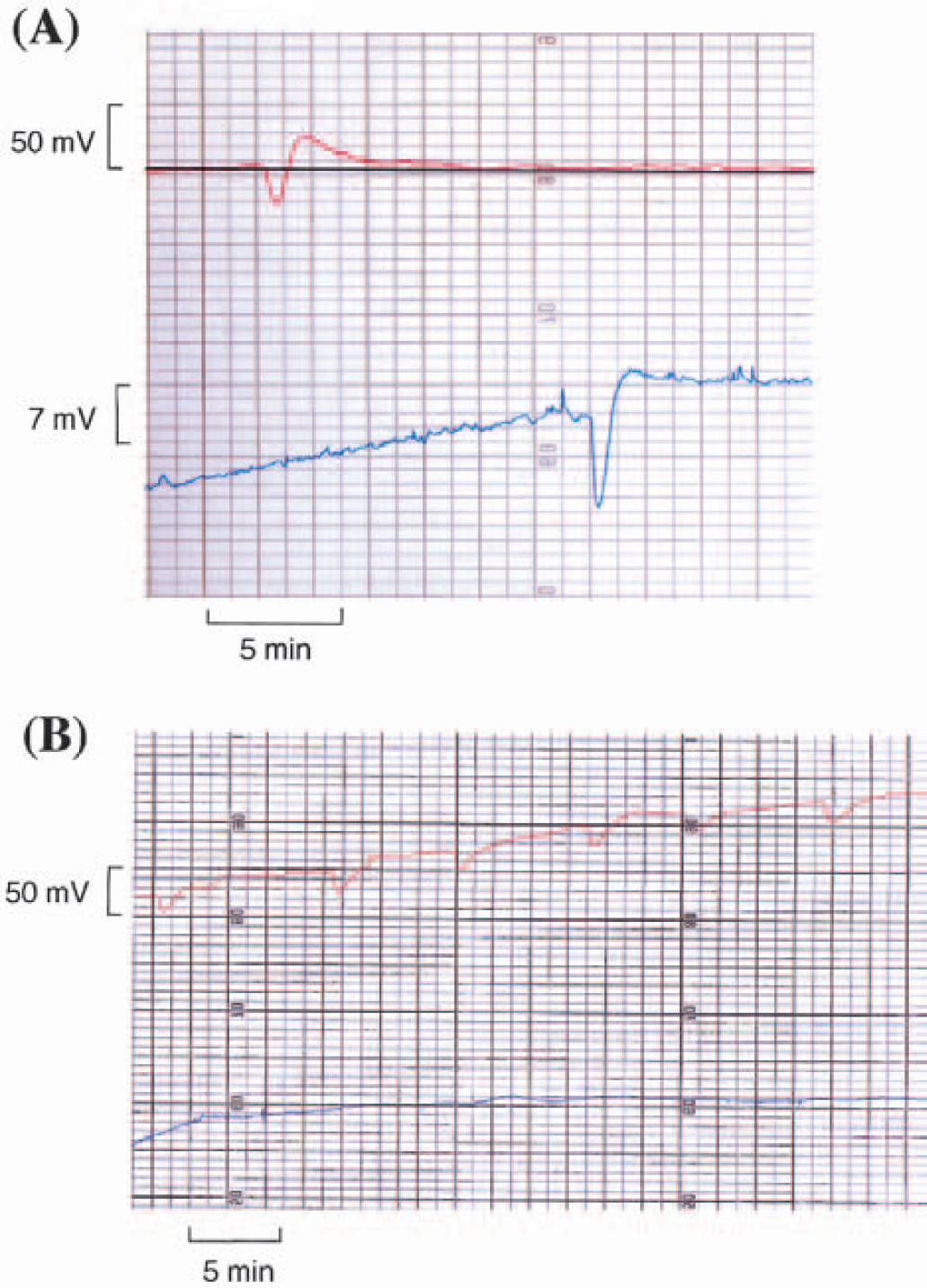
Characteristics of spreading depression waves.
Positron emission tomography study
By the visual inspection of all images of the PET study including transaxial and coronal ones, there were neither areas of persistent hypoperfusion nor spreading hypoperfusion following primary hyperperfusion after SD in any animals. Representative, multislice PET images of animal 4 in which SD was recorded twice at the rostral site and six times at the caudal site are shown in Fig. 3. Six episodes at the caudal site and one episode at the rostral site were detected immediately after KCl application, and another rostral episode occurred between the measurement of CBF1 and CBF2. The PET images obtained after the occurrence of SD demonstrated transient focal hyperemia in the occipital cortex caudally adjacent to the site of KCl application. In the hemisphere contralateral to the site of KCl application, there were no remarkable changes in regional CBF throughout the PET study in any of the animals.
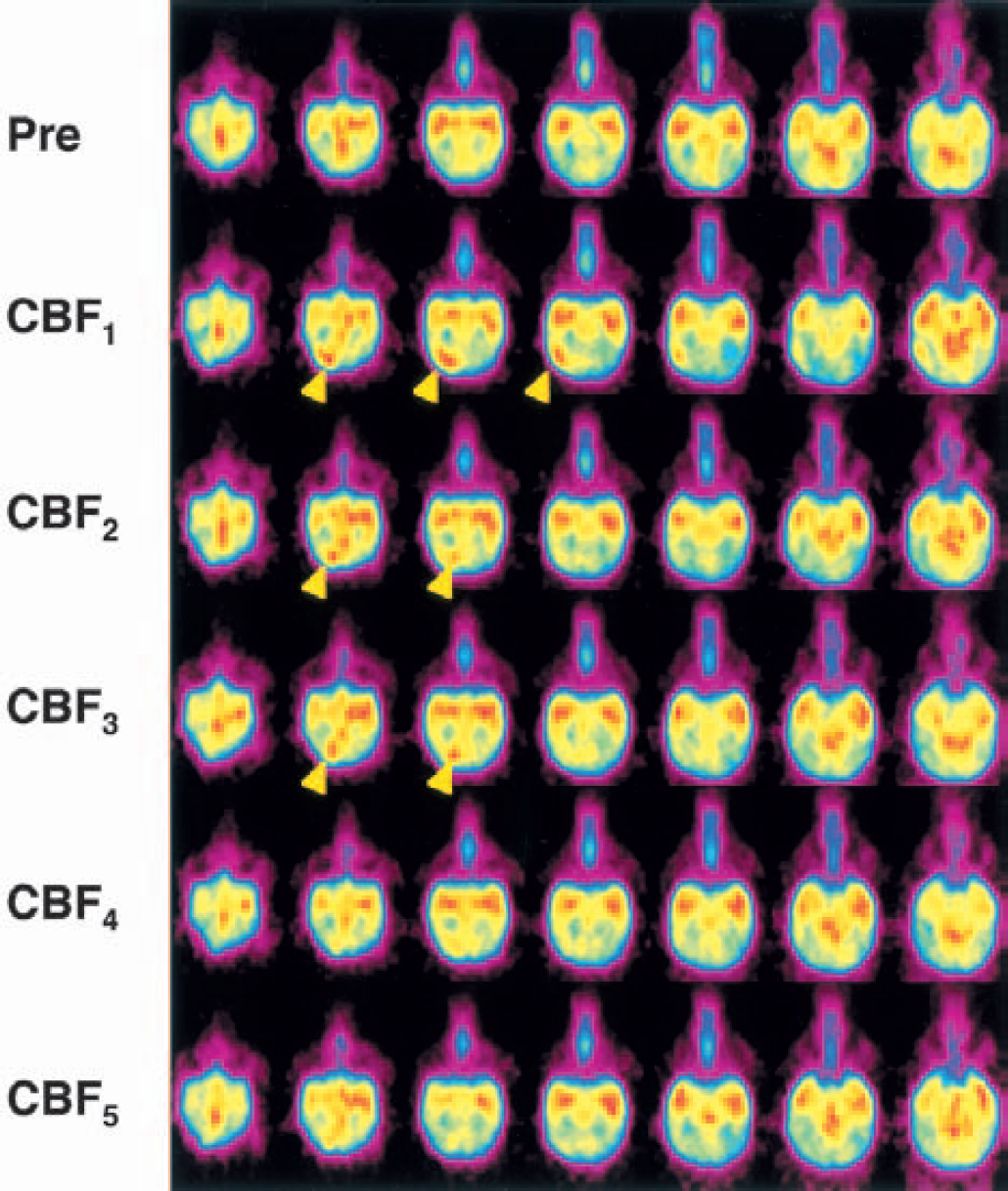
Representative positron emission tomography (PET) images showing the time course of cerebral blood flow (CBF) changes. The time course of the change in CBF in transaxial slices of 3.1-mm width before (Pre) and at various time points after KCl application in animal 4 is shown. The first PET scan after KCl application (CBF1) was taken within 10 minutes after the first spreading depression was detected. Thereafter, 4 PET scans at intervals of 10 to 20 minutes (CBF2, CBF3, CBF4, CBF5) were obtained. The PET scans in a column represent those taken at the same transaxial slice. The multislice PET images showed transient focal hyperemia (arrowhead) in the occipital cortex caudally adjacent to the site of KCl application.
Figure 4 shows the change in the asymmetry index of CBF in the 14 ROIs over time before and after the SDs in each of the 5 animals in the PET study. Focal increases in regional CBF were observed in accordance with the occurrence of SD in each animal. During the experiment, the asymmetry index increased by a maximum of 1.2 to 1.7 times in various ROIs in the five animals. In animal 1, in which no SD episodes had been detected at the rostral sites, focal hyperemia was observed after the chemical stimulation. However, none of the animals had persistent hypoperfusion in any ROIs.
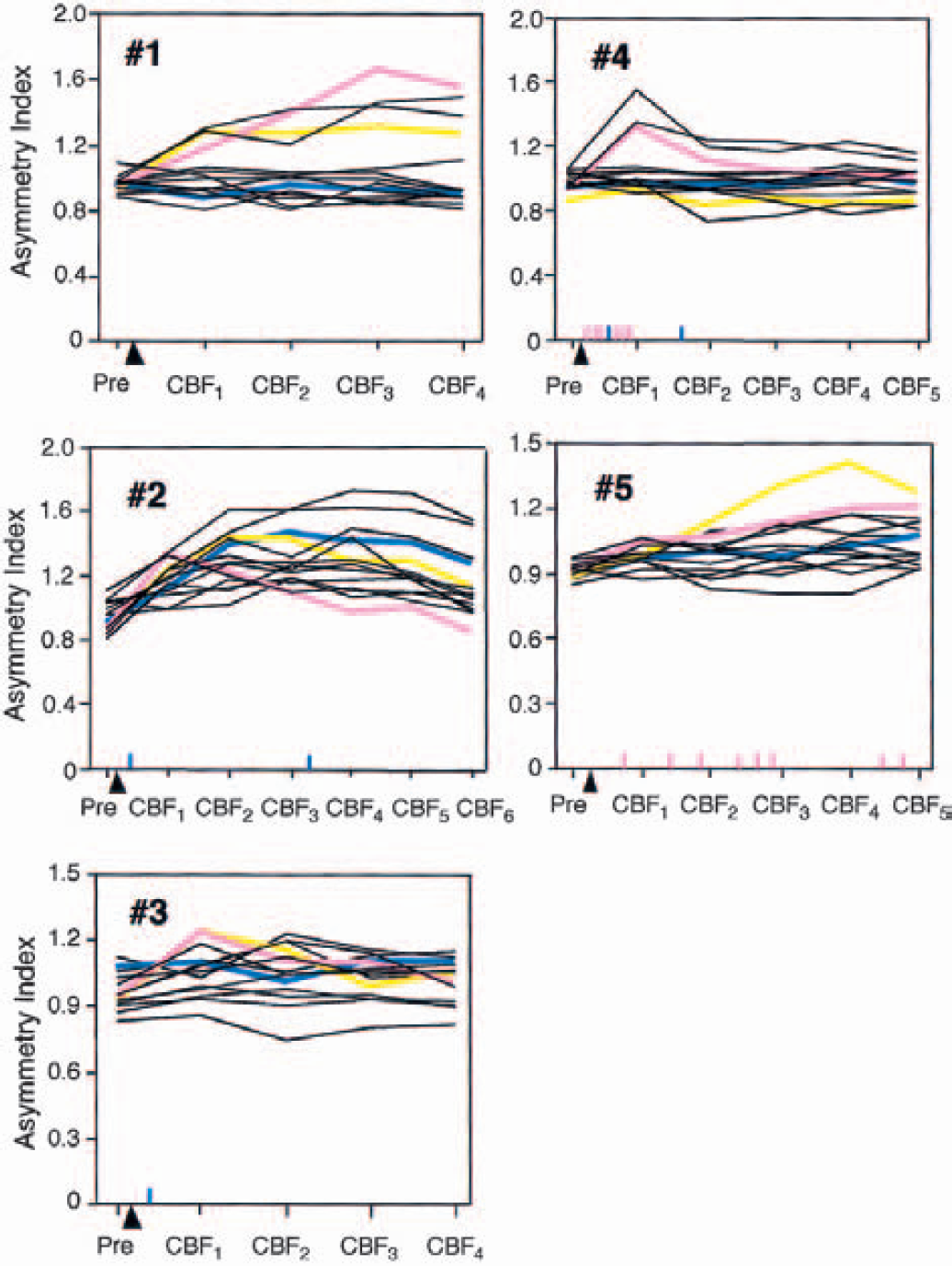
Course of the asymmetry index of cerebral blood flow (CBF) in the 14 regions of interest (ROIs) in each of the 5 animals that underwent the positron emission tomography study. The blue, pink, and yellow lines show the changes in the asymmetry index in the ROIs indicated by blue, pink, and yellow, respectively, in Fig. 1. The arrowhead indicates the time point of KCl application for eliciting spreading depressions (SDs). The time points at which SD was detected are indicated with vertical short lines above the x axis: a blue line indicates SD to the rostral direction, and a pink line indicates SD to the caudal direction. Focal increases in regional CBF were observed in accordance with the occurrence of SD in each animal. However, persistent hypoperfusion was not seen in any of the animals.
DISCUSSION
This is the first report that provides direct evidence that SD can be evoked in primates. It is known that SD waves propagate predominantly toward the caudal direction (Leao, 1944), and the present results were consistent with this. However, the measurements of SD at caudal positions were performed only in two of nine monkeys in the present study. Thus, the question as to whether SD preferentially propagates in a posterior direction remains obscure. Spreading depression waves can readily be elicited in the lissencephalic cortices (Saito et al., 1995).
In a rat model, SD waves propagated throughout a hemisphere (Hasegawa et al., 1995). In the present study, however, SD waves were infrequently detected at the rostral site close to the source of stimulation. Only a single SD was detected at the site distant from the stimulation. Because measurements of SD waves were examined by a limited number of electrodes in the present study, it may be difficult to confirm that no SD waves were detected. The detection of SD was also reported to depend on several factors, including the depths of electrode insertion into cortical tissue (Richter and Lehmenkuhler, 1993). However, we reported previously that more than 13 SD episodes could be evoked in the rostral direction in cats under the same experimental conditions as the present study (Kuge et al., 2000). SD waves can hardly be elicited, and propagate within a narrow range in primates. Differences in the cytoarchitecture among species, which are considered to reflect differences in the number and length of dendritic and axonal ramifications (Rockel et al., 1980), would influence the propagation of SD.
The CBF pattern obtained in the present study differs from those obtained in rat-and cat-SD models (Lauritzen et al., 1982; Piper et al., 1991; Kuge et al., 2000). In this study, we could not observe the prolonged hypoperfusion after the focal hyperemia. Although the size of ROIs was determined considering the spatial resolution of the PET apparatus, the possibility cannot be completely excluded that the PET would be insufficiently accurate in measuring CBF changes because of calculation of the limited number of ROIs, and the possibility of the mismatch between the ROI and the mirror region in the contralateral hemisphere. Therefore, we also ascertained by visual inspection of all PET images that there were no areas of persistent hypoperfusion as compared with the contralateral hemisphere, and also no spreading hypoperfusion was observed following focal hyperemia after SD. KCl application has been widely used because it is one of the most reliable methods of evoking SD (Marshall, 1959; Hasegawa et al., 1995; Saito et al., 1995; Takano et al., 1996; Kuge et al., 2000). An increased extracellular potassium concentration per se may increase blood flow. However, the pattern of the serial change in CBF after KCl application varied depending on the SD profile in the five animals that underwent the PET study. Although animal 1 did not show SD to the rostral direction, the pattern of the serial CBF change in this animal was similar to that in animal 5 that had eight SD episodes to the caudal direction but no episodes to the rostral direction. The dynamics of the CBF after chemical stimulation appear to be determined by the timing as well as the number of SD episodes.
Duckrow (1991) advocated a hypothesis using a rat model that hyperemia after SD would be observed only in anesthetized animals, whereas long-lasting hypoperfusion would occur after SD in both the conscious and anesthetized conditions. We previously demonstrated that a long-lasting CBF reduction followed a transient increase in CBF after a single SD in the cortex ipsilateral to the site of KCl application in cats (Kuge et al., 2000), which is consistent with a rat-SD model (Lauritzen et al., 1982). Flow-metabolism uncoupling with persistent hypoperfusion in the cortex of cats with SD indicated that cerebrovascular reactivity was impaired in association with SD (Kuge et al., 2000). In this study, persistent hypoperfusion was not seen in any ROI in the five animals. Wahl et al. (1987) demonstrated that the vascular responses to SD differed between cats and rats. Based on our study, the vascular responses to SD in primates may also differ from those in cats.
The changes in CBF during an SD phenomenon in primates also differed from those in patients with migraine (Olesen et al., 1981; Lauritzen et al., 1983; Lauritzen and Olesen, 1984; Woods et al., 1994; Hadjikhani et al., 2001), although the scanning procedures and data analysis of PET were the same as those of the previous report (Woods et al., 1994). Because a pain stimulus results in a reduction in global CBF in humans (Coghill et al., 1998), the migraine headache itself may influence the dynamics of CBF in patients with a migraine attack. It was also indicated that SD may not be associated with migraine headache (Ebersberger et al., 2001). Furthermore, SD in the human brain, if present, would propagate much more slowly than observed in a previous migraine study (Woods et al., 1994), because SD waves propagate on the cortical convolution.
One should be cautious about projecting our results to the ischemic brain of primates and questioning the role of SDs in the evolution of brain infarction. Spreading depression is a fully reversible process in the normal brain and does not cause neuronal damage (Nedergaard and Hansen, 1988). However, in moderately ischemic brain as is present in the ischemic penumbra, SD waves, namely, periinfarct depolarizations, may be associated with episodic energy failure in the acute ischemic penumbra in primates, as demonstrated in rats (Back et al., 1996). We have reported recently that a CBF and cerebral metabolic rate of glucose uncoupling was observed in the parietal cortex and several regions surrounding the ischemic core in a primate model of thromboembolic stroke (Kuge et al., 2001). Although the mechanisms underlying the elevated glucose metabolism and the process of uncoupling of flow and metabolism remained to be clarified, anaerobic glycolysis and SD are proposed as factors contributing to elevated glucose metabolism in early phases of ischemia (Ginsberg et al., 1977; Shinohara et al., 1979; Tanaka et al., 1985; Hasegawa et al., 1990; Back et al., 1995; Kuge et al., 2000). The contribution of SD to pathophysiology of ischemic stroke cannot be resolved by the findings obtained from the present study. However, the poor migration of SDs and rare evidence of migrating SD waves in primates may account for the negative result in patients with stroke (Back et al., 2000).
The present study suggests that one must be careful when applying the experimental results of SD obtained from nonprimate animals to the pathogenesis of brain diseases in humans. The dynamics of SDs may vary among species because of differences in convolutional and cytoarchitectural structure, and the vascular responses to SDs. First, SD occurred less frequently in our primate model than in nonprimate models. Its propagation was limited to areas near the site of stimulation in the cynomolgus monkeys, although it propagated to the entire hemisphere in rats. Second, immediately after the first SD was elicited, focal cortical hyperemia was detected, but it was not followed by spreading hypoperfusion or persistent hypoperfusion in the hemisphere ipsilateral to the site of chemical stimulation. These unique features of SD in this primate demand reappraisal of the hypothesis that SD contributes to the pathogenesis of human brain diseases.