
Abstract
The authors previously provided evidence that spreading depression (SD) can be evoked in primates. Cyclooxygenase-2 (COX-2) expression has been found to increase in the rodent cortex undergoing SD, and the authors sought to determine whether this association exists in primate brain. In the present study, neuronal COX-2 expression was induced during SD in the primate cortex. The mean expression ratio of COX-2 messenger RNA in animals with SD was significantly higher than that measured in controls (1.69 vs. 0.5; P = 0.02). Induction of COX-2 in these animals was also detected by human microarray analysis. Results show that, as in rodents, neuronal COX-2 is induced in the primate cortex in response to SD.
Cortical spreading depression (SD), the reversible depression of cortical electrical activity, plays a role in the development of cerebral ischemic injury and migraine (Lauritzen et al., 1983; Olesen et al., 1981; Woods et al., 1994). Repetitive, pathologic SDs were similarly found to play a role in the development of ischemic injury under conditions of focal brain ischemia in rats (Gill et al., 1992; Hossmann, 1994; Iijima et al., 1992; Takano et al., 1996). Changes in cerebral blood flow (CBF) in experimental SD models in rats and cats were characterized by transient focal cortical hyperemia, followed by persistent hypoperfusion (Kuge et al., 2000; Lauritzen et al., 1982; Piper et al., 1991). Recently, we provided the first direct evidence that SD, accompanied by focal cortical hyperemia, can be evoked in primates (Yokota et al., 2002). The long-lasting hypoperfusion that followed this hyperemia in rats and cats has not been observed in primates.
The gene for cyclooxygenase-2 (COX-2), a rate-limiting enzyme in prostaglandin synthesis, was induced in the nonprimate cortex during SD (Koistinaho et al., 1999; Miettinen et al., 1997). Cyclooxygenase-2 appeared to mediate the increase in CBF produced by synaptic activity in the somatosensory cortex in mice (Niwa et al., 2000). The contribution of SD and its associated genes to the pathogenesis of human brain diseases has not been elucidated.
As a first step in determining which genes show altered expression during SD in primates, we examined whether the COX-2 gene was upregulated. Using a human complementary DNA (cDNA) array system, we also examined the effect of SDs on gene-expression profiles.
MATERIALS AND METHODS
Spreading depression model and brain preparation
Nine adult, male cynomolgus monkeys were divided into two groups: a normal control group (group C, n = 3) and a group in which SD was evoked by applying 3.3-mol/L KCl to the cortex (group SD, n = 6). All procedures were approved by our Institutional Animal Research Committee and were performed in accordance with standards published by the National Research Council in the Guide for the Care and Use of Laboratory Animals.
Shifts of direct current potential were measured in four animals in the SD group only with a microelectrode placed just rostral to the chemical stimulation site; a single episode was recorded in three animals, and six episodes were recorded in one animal. Direct current potential was measured in the remaining two animals with a microelectrode that was placed caudal to the chemical stimulation site as well as with micro-electrodes placed rostral to the chemical stimulation site; one of these animals was subjected to eight recording episodes at this site, whereas the remaining animal was subjected to two direct current shifts, each of which was recorded from a point that was rostrally adjacent to the chemical stimulation site, as well as to six episodes at the caudal site.
Two hours after KCl application, the brains from these animals were perfused with cold saline, after which the animals were killed and the brains removed. Several samples from each cortex were harvested and stored at −80°C until use, whereas other samples were embedded in paraffin.
RNA blot analysis
RNA preparation and blot analysis were performed as previously described (Inoue et al., 1995). Cyclooxygenase-2 messenger RNA (mRNA) in each region was expressed as the ratio of the COX-2 mRNA signal to the GAPDH mRNA signal in that region (expression ratio).
Microarray analysis
Microarray analysis was conducted using Genome System (St. Louis, MO, U.S.A.) as described elsewhere (Lyer et al., 1999). The microarray contained 9,182 elements: 8,412 unique annotated genes or expressed sequence tags, and 4,757 characterized human protein genes. Left cortical RNAs from the SD (>5 episodes; n = 3) and control (n = 3) groups were pooled. Poly (A)+ RNAs were purified from these RNAs using oligo(dT)30-latex (Takara, Inc., Shiga, Japan) to use as templates for cDNA synthesis. The cDNA probes were reverse transcribed with 5′Cy3 or fluorescently labeled Cy5 and were hybridized with Human UniGEM V (version 2.0). The average of the total Cy3 and Cy5 signals yielded a ratio that was used to normalize the signals.
Immunoblot analyses
Left temporal cortical samples were weighed and homogenized in 10 volumes of cold 62.5-mmol/L Tris buffer (pH 6.8) containing 1% sodium dodecylsulfate, 10% glycerin, and 5% 2-mercaptoethanol. Tissue homogenates were heated to 90°C for 5 minutes and then centrifuged at 15,000g for 10 minutes at 4°C. The supernatants were separated by SDS-PAGE (5% to 20% acrylamide gradient) and proteins transferred onto a nitrocellulose sheet. The blots were incubated with anti–COX-2 antibody (dilution 1:1,000; Cayman Chemical, Ann Arbor, MI, U.S.A.) for 1 h at 25°C and were then washed in 50-mmol/L phosphate-buffered saline (pH 7.4) containing 0.05% Tween 20.
Immunohistochemistry
A mirror-sectioning technique was used to colocalize COX-2 and microtubule-associated protein 2 (MAP-2), a neuronal skeletal protein. Deparaffinized temporal cortical sections (3 μm) were incubated with a polyclonal anti–COX-2 antibody (dilution 1:100; Cayman Chemical) and a monoclonal anti–MAP-2 antibody (clone HM-2; dilution 1:2,400; Sigma, St. Louis, MO, U.S.A.) overnight at 4°C. The sections were washed with phosphate-buffered saline, and biotinylated goat anti–rabbit immunoglobulin (Vector Laboratories, Burlingame, CA, U.S.A.) or biotinylated F(ab′)2 rabbit anti–mouse immunoglobulin (dilution 1:500; Dako, Carpinteria, CA, U.S.A.) was applied to the sections, which were then incubated for 30 minutes at 25°C. Labeling was visualized using a Vectastain Elite Kit (Vector Laboratories).
Statistical analysis
Data are expressed as the mean ± standard deviation. Comparisons of COX-2 mRNA expression between groups were made using the Mann-Whitney U test. A two-tailed P value less than 0.05 was considered to be significant.
RESULTS
Expression of cyclooxygenase-2 mRNA
Cyclooxygenase-2 mRNA expression increased in the cortex in which SD was induced (left) compared with both the contralateral side and the left cortex in group C (Fig. 1A). The expression ratio of COX-2 mRNA in the left cortex in the SD group (1.69 ± 0.57) was significantly greater (P = 0.02) than in the left cortex in group C (0.5 ± 0.05; Fig. 1B).
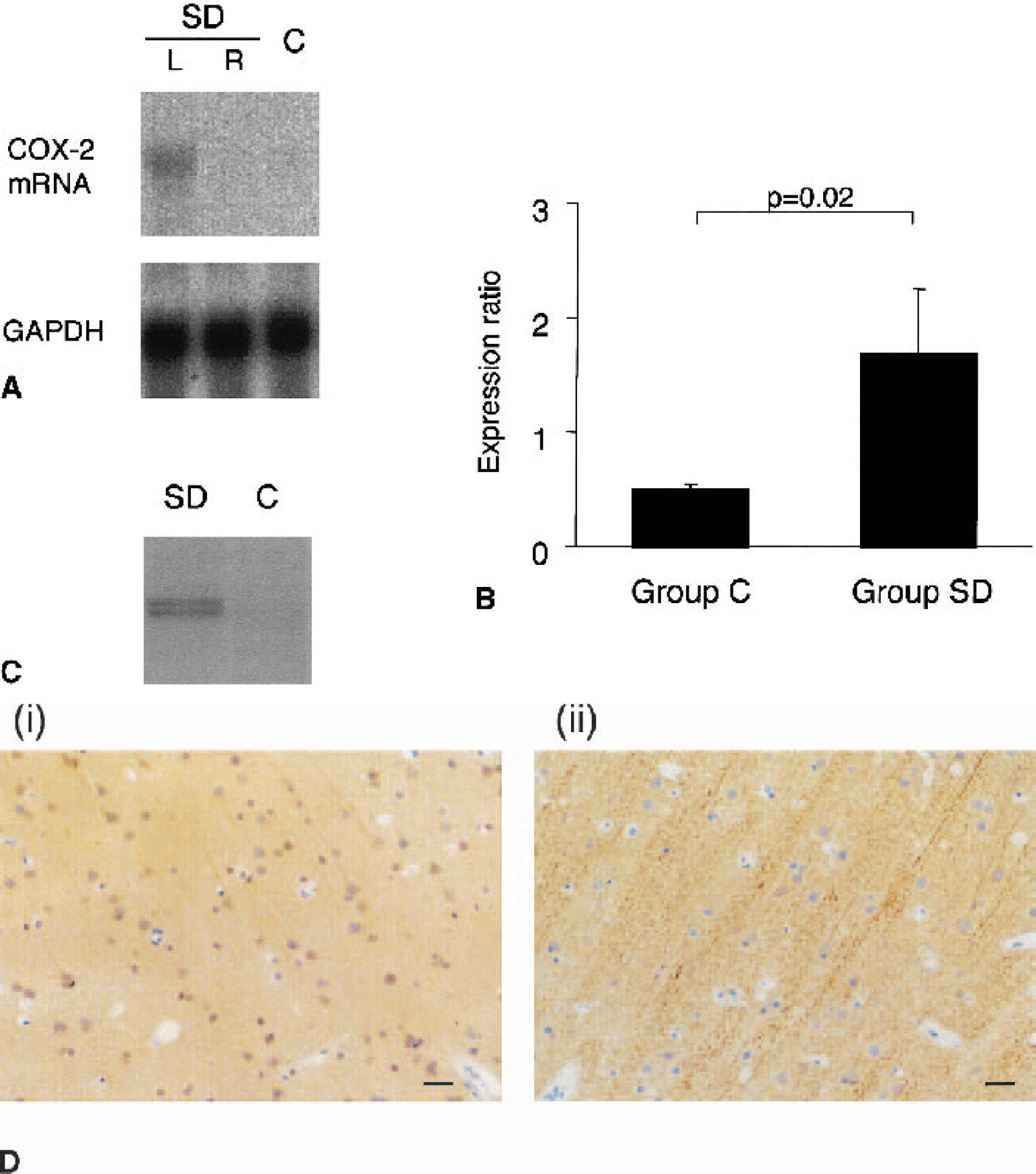
Cyclooxygenase-2 (COX-2) expression in the primate brain.
Cyclooxygenase-2 localization
Immunoblots revealed a 70- to 72-kd COX-2–immunoreactive band (Gidlund et al., 1981) in tissue derived from the SD group, which was barely detectable in group C (Fig. 1C). Immunoreactive neurons were observed in animals that received six SD episodes rostral to the chemical stimulation site (Fig. 1D). Immunoreactive MAP-2 was demonstrable in the same neurons that displayed COX-2 immunoreactivity.
Gene-expression patterns
Increases in normalized gene-expression signals above 1.5-fold were observed for two genes among a total 9,182 elements: the COX-2 (1.6-fold) and basic transcription element–binding protein 1 (1.6-fold) genes. Twenty-one elements were found to be different by at least 1.4-fold. Figure 2 shows representative images of scanned arrays hybridized with left cortical samples.
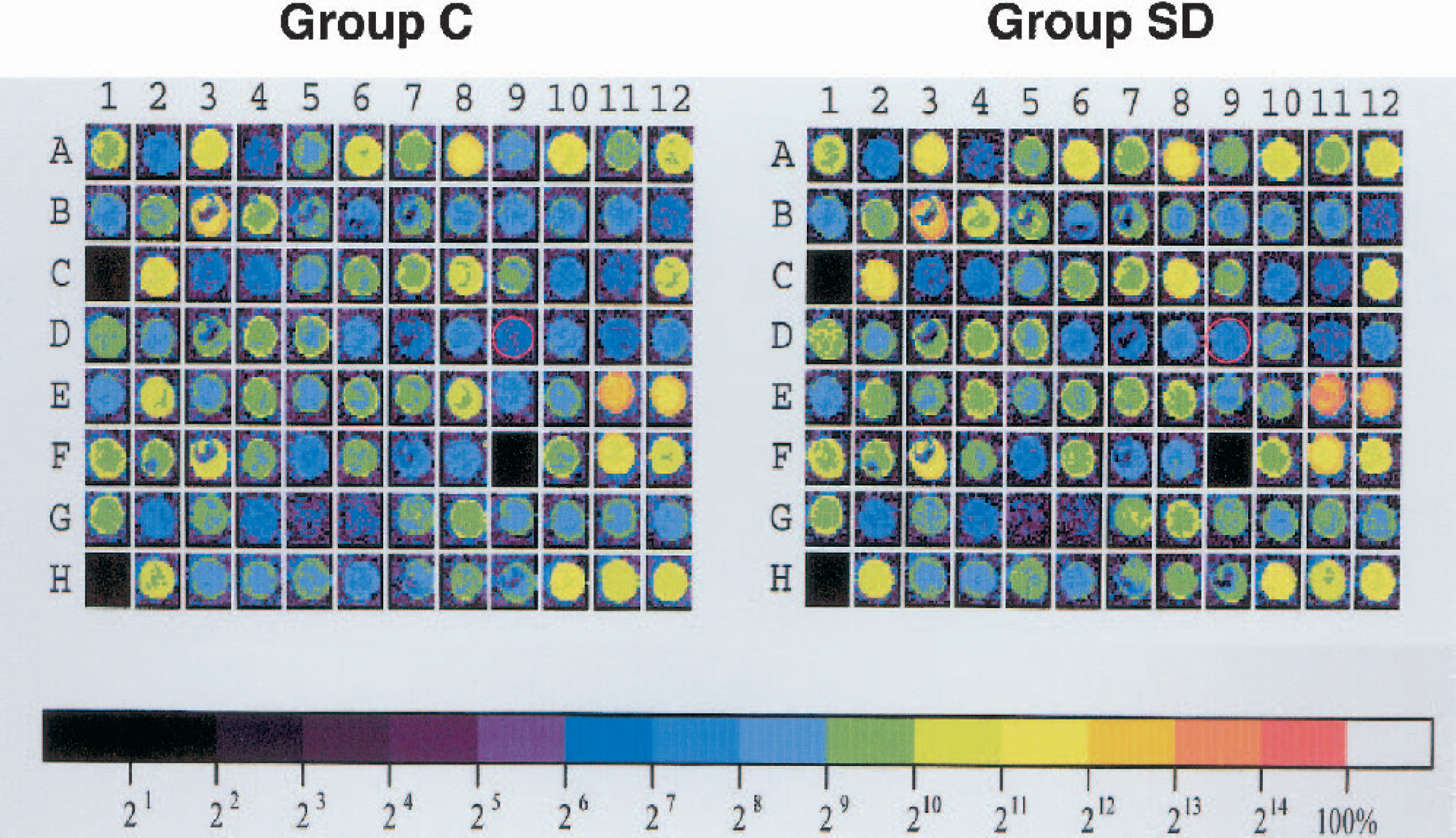
Gene-expression patterns in the brains of animals in the spreading depression (SD) and control (C) groups. Representative images of scanned arrays hybridized with cortical samples from C and SD animals are shown. Of the 9,182 elements that were examined, expression of the cyclooxygenase-2 gene (red circles) was found to be increased by 1.6-fold in the SD group.
DISCUSSION
Previous studies have suggested that COX-2 plays a role in the development of ischemic injury during focal brain ischemia in rodents (Collaco-Moraes et al., 1996; Nogawa et al., 1997) and that prevention of COX-2 expression might be beneficial in treating human ischemic stroke. Ischemia-related SDs have been regarded as good targets for pharmacologic intervention in stroke because they exacerbate the preexisting energy depletion in the periinfarct zone (Hossmann, 1994; Takano et al., 1996). Cyclooxygenase-2 was induced after SD and as a result of focal brain ischemia in rat models (Koistinaho et al., 1999; Miettinen et al., 1997). Koistinaho and Chan (2000) reported that SD directly induced COX-2 expression in focal brain ischemia by stimulating the NMDA receptor and activating phospholipase A2. Only a few postmortem studies, however, reported COX-2 expression in human brain ischemia (Iadecola et al., 1999; Sairanen et al., 1998).
In our study, COX-2 was induced in the cortices that experienced SD, supporting previous observations in rodents (Koistinaho et al., 1999; Miettinen et al., 1997). Cyclooxygenase-2 gene expression increased by 1.6-fold in the SD group, as detected using a DNA microarray. The gene transcribing basic transcription element–binding protein 1, a thyroid hormone–regulated gene found in the developing rat brain, was also upregulated (Cayrou et al., 2002); the relation between this protein and SD remains to be clarified. Enard et al. (2002) showed numerous quantitative differences in gene expression between closely related mammalian species. Because they showed that such differences were particularly pronounced in the human brain, primate cDNA arrays analysis will need to be improved before we can fully identify candidate genes that may be involved in SD. In summary, as in rodents, neuronal COX-2 is induced in the primate cortex in response to SD.
Footnotes
Acknowledgments:
The authors thank Dr. Toshiho Ohtsuki for his scientific input.