
Abstract
Functional neuroimaging studies have suggested a specific role of the extrastriate cortex in letter string and visual word form processing. However, this region has been shown to be involved in object recognition and its specificity for the processing of linguistic stimuli may be questioned. The authors used an event-related functional magnetic resonance imaging design with category priming to record the response elicited by the passive viewing of single letters, geometric figures, and of the categorically ambiguous stimulus “O” that pertains to both sets of familiar symbols. Bilateral activations in the extrastriate cortex were found, with a left predominance particularly pronounced for the ambiguous stimulus. Individual analysis of spatial extent and signal intensity showed a priming × stimulus × hemisphere interaction. When primed by the congruous categoric set, a bilateral decrease in activation was observed for letters and geometric figures. The ambiguous stimulus behaved as a letter for the left hemisphere, with decreased activation when primed by letters, whereas in the right hemisphere, an adaptation effect occurred when primed by geometric figures. These priming effects suggest that, for the ambiguous stimulus, letter processing was systematically involved in the left extrastriate cortex. The current results support the existence of a neural substrate for the abstract category of letters.
Keywords
Following the early work of Petersen et al. (1990), a number of functional neuroimaging studies have investigated the role of the extrastriate cortex in the processing of visually presented linguistic stimuli. Although considerable evidence for the involvement of these regions in orthographic and visual word form processing has been obtained, particularly for the left fusiform gyrus and lateral occipitotemporal cortex (Petersen et al., 1990; Price et al., 1996; Puce et al., 1996; Pugh et al., 1996; Kuriki et al., 1998; Tarkiainen et al., 1999; Uchida et al., 1999), the specific nature of the processes supported may be questioned because of possible confounding effects related to the stimuli and tasks used in the experimental design. These effects include the level of familiarity (Price et al., 1996; Tagamets et al., 2000), the meaningfulness (Moore and Price, 1999), the string length (Indefrey et al., 1997; Fujimaki et al., 1999; Tarkiainen et al., 1999), and the structural characteristics of the stimuli. The latter point in particular may be of crucial importance as the lateral occipital complex (LOC) (Malach et al., 1995) has been shown to respond more strongly to structured objects (Kourtzi and Kanwisher, 2000).
Further data on the involvement of the extrastriate cortex in word processing have been obtained from word priming studies coupled with functional cerebral imaging (Buckner et al., 1995, 2000; Badgaiyan and Posner, 1997; Doyle and Rugg, 1998; Badgaiyan et al., 1999; Backman et al., 2000; Yasuno et al., 2000), but without clear evidence for hemisphere predominance. Again, as facilitation effects in the same regions also have been observed with object priming (Beason-Held et al., 1998; Buckner et al., 1998; Buckner and Koutstaal, 1998; Grill-Spector et al., 1999; Henson et al., 2000; Kourtzi and Kanwisher, 2000), the specific role of the inferior occipitotemporal cortex in perceptual and even conceptual priming effects with linguistic stimuli is unclear.
One question that subsists is whether the extrastriate cortex would show any difference in activation for single letters compared with familiar, structured, and meaningful stimuli, a difference that would indicate some degree of specificity for alphabetic processing, notably in the left hemisphere. Indeed, the existence of letter representations and the occurrence of automatic letter processing in the left inferior occipitotemporal cortex has been suggested by some recent studies (Polk and Farah, 1998; Garrett et al., 2000). Because the neural networks that subserve reading are multicomponential and likely include elementary visual analysis, Tagamets et al. (2000) stressed the importance of no change between conditions other than that in the stimuli presented so that all other aspects of the task be identical. An alternate approach would make use of a strictly identical stimulus presented under conditions that vary in one aspect. With respect to the question raised above, a convenient experimental design would be to check whether differences appear for a physically identical, but categorically ambiguous, stimulus when opposing the categoric context of presentation. Using such an experimental design, Celsis et al. (1999) found a difference in the left occipitotemporal region for the perception of an ambiguous stimulus presented in speech and nonspeech contexts. This finding suggests that a phonetic representation of the ambiguous stimulus was maintained and accessed in the nonspeech context as the result of automatic, autonomous processes.
In the current study, the authors used event-related functional magnetic resonance imaging (fMRI) to assess the activations elicited by the passive viewing of letters, familiar geometric figures, and of an ambiguous stimulus that pertains to both categoric sets. To elicit context effects and to attempt to differentiate specific visual form effects from abstract representation and more conceptual processes, the authors used a priming design. The main objective of the study was to evidence differences in the extrastriate response to the ambiguous stimulus, in particular a priming context × hemisphere interaction that would support, particularly in a passive viewing experiment, the existence of abstract, automatic processes in letter perception and identification.
MATERIALS AND METHODS
Subjects
Six right-handed, healthy women (aged 23 to 30 years) were tested during a passive visualization task. All six subjects were university graduates. After explanation of purpose and procedure, written informed consent was obtained before scanning, and the Toulouse Human Subjects Protection Committee approved the study.
Image acquisition
Images were acquired on a 1.5 Tesla Siemens Magnetom Vision (Siemens Medical Systems, Erlangen, Germany) equipped with echo planar imaging hardware. For functional MR images, 10 contiguous slices (6-mm-thick) parallel to the intercommissural plane (from z = −18 mm to z = 36 mm) were collected using a T2 *-weighted gradient echoplanar imaging sequence (flip angle = 90°, TE = 66 milliseconds, TR = 2.5 seconds, field of view = 20 cm, 64 × 64 interpolated to 128 × 128 matrix).
To perform projections of the activations on anatomic images, a three-dimensional, high-resolution T1 -weighted data set of the whole brain was acquired for each volunteer (flip angle = 12°, echo time = 7 milliseconds, repetition time = 15 milliseconds, field of view = 30 cm, 256 × 256 matrix, 128 slices 1 mm in thickness).
Stimuli
Two sets of 10 stimuli were used (Fig. 1). The letter set (L) consisted of nine uppercase letters made of two, three, or four straight lines plus the letter “O”. The figure set (F) consisted of nine familiar geometric figures—that is, polygons with three or four sides—plus the circle. The letter “O” and the circle were physically identical. Both categoric sets (L and F) thus included one ambiguous item that can be categorized either as a letter or as a familiar geometric figure. However, one salient visual feature distinguished the ambiguous stimulus from all other stimuli, as the “O” was the only curved form. In addition, a second feature distinguished the ambiguous stimulus from the letters, but not from the geometric figures. Indeed, the ambiguous stimulus appeared as a closed contour, alike the polygons of the figure set, whereas the stimuli of the letter set were made of open sets of lines, with the only exception of the A that includes a closed part.

The two sets of stimuli. The letter set consisted of 10 uppercase letters, including the letter “O”, and the figure set consisted of 10 simple geometric figures, including the circle. The letter “O” and the circle were physically identical.
All stimuli were presented foveally using a system of binocular glasses (Resonance Technology Incorporation, Northridge, CA, U.S.A.), subtending an angle 0.75° in width and 1° in height, and appeared white on a black screen.
Experimental protocol
A run was divided into two steps (Fig. 2). The first step was a priming period followed by a rest period, and the second step was the imaging period during which each presentation of one of the stimuli was followed by six acquisitions that sampled the fMRI response.
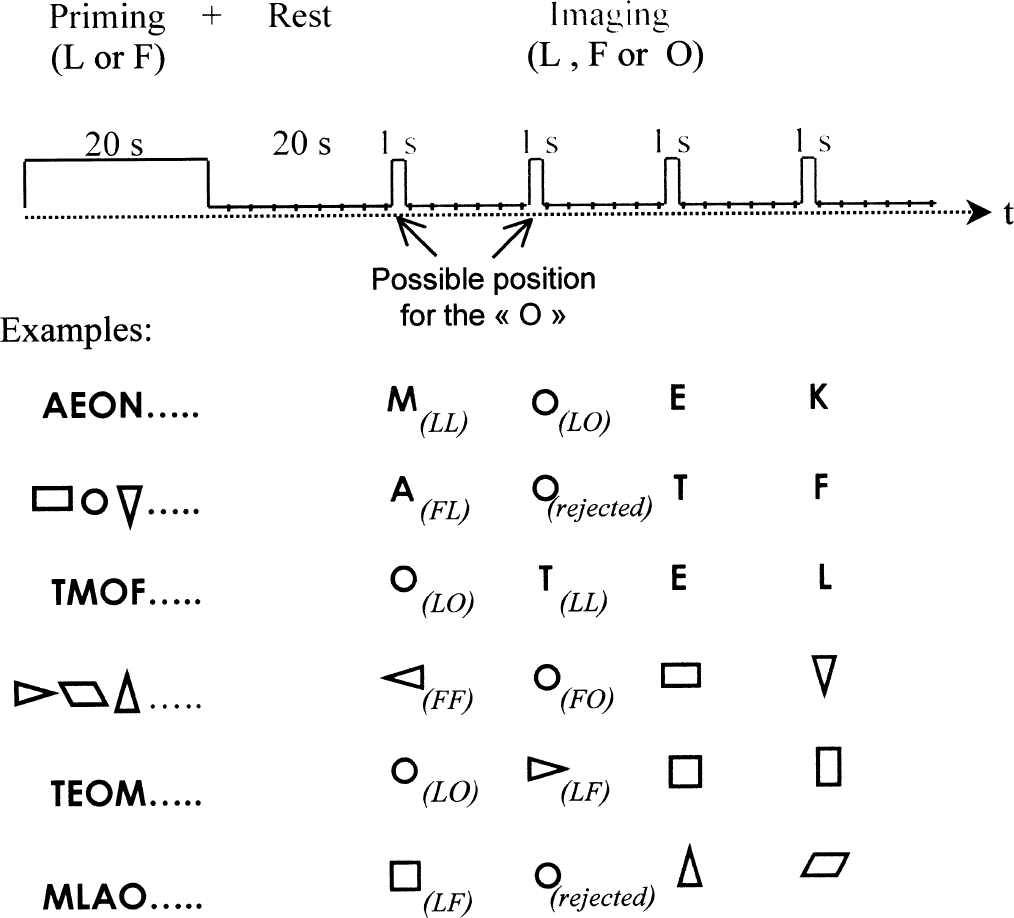
The experimental protocol. (top) The sequence of periods and events in one run of the experimental protocol. Examples of runs are showed to explain how each stimulus, processed individually in the event-related functional magnetic resonance imaging design, was allocated to one of the six experimental conditions. Note that the ambiguous stimulus “O” was not retained when in second position in the incongruous conditions (LF or FL, see text).
During the priming period, each stimuli of a given categoric set (L or F), including the “O”, was randomly displayed twice, for 1 second and with no blank between presentations. The duration of the priming period thus equaled 20 seconds. No acquisition was performed during priming, but subjects were hearing, through special earphones (Resonance Technology), the recorded fMRI noise to keep constant the sound environment during the entire run.
After the priming period, a 20-second black screen was presented to allow the hemodynamic response to return to baseline. During this rest period, 8 acquisitions were performed; the first 7 were discarded to allow for T1 equilibration.
During the imaging period, four single stimuli were presented, either from the letter set (L) or from the figure set (F). Each stimulus was presented for 1 second, and 15 seconds separated 2 stimuli. During these 15 seconds, 6 volumes (10 slices, repetition time = 2.5 seconds) were acquired to sample the fMRI response. Such an event-related fMRI design with low stimulus-presentation rate and optimal contrast interstimulus interval (Bandettini and Cox, 2000) minimizes the direct effect of the preceding stimulus on the stimulus-specific response (Friston et al., 1998). Among the four stimuli, one and only one ambiguous symbol “O” was presented, and always in the first or second position (to minimize the lag between the priming context and the ambiguous stimulus).
Combining priming and imaging periods, four types of run were produced: letter priming/letter imaging, figure priming/letter imaging, figure priming/figure imaging, and letter priming/figure imaging. Each combination was repeated 6 times, giving a total of 24 runs per subject. The 24 runs were randomly presented with regard to the different combinations, and the presentation order of the runs differed from one subject to each other.
The event-related fMRI paradigm allowed the separation of each stimulus and particularly the separation, within a run, of the ambiguous stimulus “O” from the other stimuli (letters and figures). Finally, the authors' experimental design allowed them to process separately the “O”, the letters, and the figures. The ambiguous stimulus was presented 24 times across all the imaging periods, whereas each of the remaining stimuli was presented only 4 times. Including the presentations of the priming periods, the repetition rate of the ambiguous stimulus thus was approximately three times that of any other stimulus.
The three types of stimuli combined with the two priming categories produced six experimental conditions as follows (see examples in Fig. 2):
LL: letters, excluding the ambiguous stimulus, presented after letter priming
FL: letters, excluding the ambiguous stimulus, presented after figure priming
LF: figures, excluding the ambiguous stimulus, presented after letter priming
FF: figures, excluding the ambiguous stimulus, presented after figure priming
LO: the ambiguous stimulus “O” presented after letter priming
FO: the ambiguous stimulus “O” presented after figure priming
The LL and FF conditions are referred to as congruous conditions, because the stimuli used for priming and imaging pertained to the same categoric set, whereas the LF and FL conditions are referred to as incongruous conditions because the stimuli pertained to different categoric sets. The LO and FO conditions are referred to as ambiguous conditions as only the ambiguous stimulus “O” was used to generate the images.
When extracting the ambiguous conditions from the figure priming/letter imaging and letter priming/figure imaging combinations, the ambiguous stimulus was retained when it was in first position but was never retained when in second position to avoid the category priming effect to be broken. However, the ambiguous stimulus was retained regardless of its position when presented in the homogeneous priming–imaging combinations. This led to 8 presentations of the ambiguous stimulus per condition and per subject. The other stimuli retained for the statistical analysis were the event in first position when the ambiguous stimulus retained was in second position, and conversely, the event in second position when the ambiguous stimulus retained was in first position. This selection was performed to prevent a possible position effect on the ambiguous character that was always presented in first or second position, because the other events were also in first or second position. This led to six presentations of unambiguous stimuli per condition and per subject.
Group analysis
Images were processed with SPM96 software (Welcome Department of Cognitive Neurology, London, U.K.). Echoplanar images were resized in the standard anatomic space defined by the atlas of Talairach and Tournoux (Talairach and Tournoux, 1988). Images were next realigned to the first functional image of each subject to correct residual head movement. The functional images of each run and each subject then were coregistered with the first functional image of one subject taken as reference subject. After coregistration, functional images were smoothed with an 8-mm full width at half maximum Gaussian kernel to reduce the residual anatomic variability between subjects.
The hemodynamic response for the group analysis using the general linear model was supplied as a convolution of the input function with a Poisson function (λ = 6). The input function used for the convolution was a boxcar signal with 1 for the first image and 0 for the remaining 5 images of the sampling. The statistical analysis was applied on the images corresponding to the 6 conditions described above (LL, FL, LF, FF, LO, and FO). The thresholds used for the 6 conditions were P = 0.001 for the voxel height (Z > 3.09, uncorrected) and n > 30 voxels for spatial extent.
Individual analysis
Two variables were used in the analysis of the data at the individual level: the spatial extent of the activated areas and the average MR signal intensity obtained in the region of interest (ROI) identified from localizer runs.
To analyze the spatial extent of the activated regions, the authors first constructed Z maps for each subject with SPM96. Identical thresholds were applied to each subject:P = 0.01 for the Z score value (Z > 2.33, uncorrected) and n > 10 voxels for the extent threshold. The authors then counted the number of activated voxels that pertained to the visual areas significantly activated in the group analysis in each condition for each hemisphere and for each subject. The number of activated voxels was introduced as the dependent variable in a three-factor, repeated measure multivariate analysis of variance (MANOVA) (priming × stimulus × hemisphere). The multivariate analysis of variance bypasses the assumption of sphericity for factors with more than two levels. Comparisons involving the three-level stimulus factor are thus reported using Rao's R values for significance testing.
To estimate the MR signal intensity, a ROI was first defined for each subject from an independent data set. This set consisted of all the event-related scans corresponding to the third and fourth stimuli of each run that were not otherwise used in the study. The use of independently defined ROIs by means of localizer runs has been recently introduced in the design of fMRI studies (Gauthier et al., 2000; Kourtzi and Kanwisher, 2000). It allows for an individual definition of the area of interest in which the signal intensity in the images to be analyzed will not depend on the threshold used for voxel selection. Using the same hemodynamic response model as in the group analysis, the ROI was obtained by thresholding the Z maps at z > 2.33 (P < 0.01) in the slices with z ranging from 0 to −18 mm. Then, for each event-related fMR response after the presentation of the first and second stimuli of each run (original data set), the time course of MR signal intensity in each voxel of the ROI was plotted from time point 0 (image just preceding the stimulus) to time point 6 (last image sampling the fMR response) and was normalized to the minimum. The authors then calculated the average value for time point 2 (the image with a 4-second lag after the beginning of the stimulus that was expected to correspond to the peak response) for each hemisphere and according to each experimental condition. These values were submitted to a three-factor, repeated measure MANOVA (priming × stimulus × hemisphere).
RESULTS
Group analysis
Figure 3 shows the activations observed in the six conditions for the group of six subjects, projected on the anatomic images of the reference subject. Table 1 indicates, for each condition and in each hemisphere, the Talairach coordinates of the voxel with the maximum Z score, the corresponding Brodmann area, the number of voxels in the activated areas, and the value of the maximum Z score.
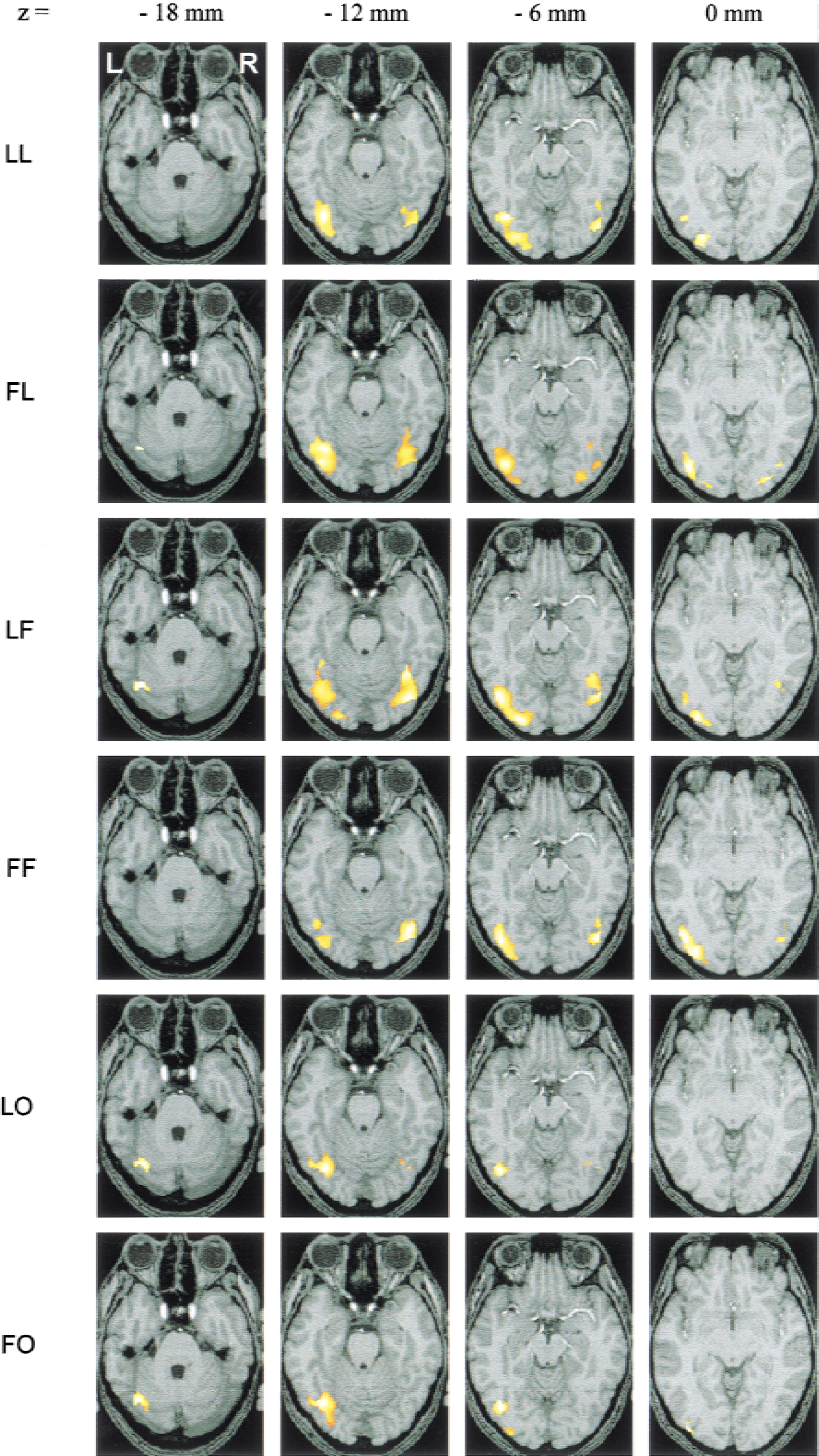
Activated areas in group analysis. Activations are projected on the images of the subject taken as reference for coregistration. Activations are thresholded at P = 0.001 for height threshold (Z > 3.09) and at n > 30 voxels for extent threshold for the 6 conditions. See text for explanation of abbreviations.
Loci of activations obtained from the group analysis for each condition
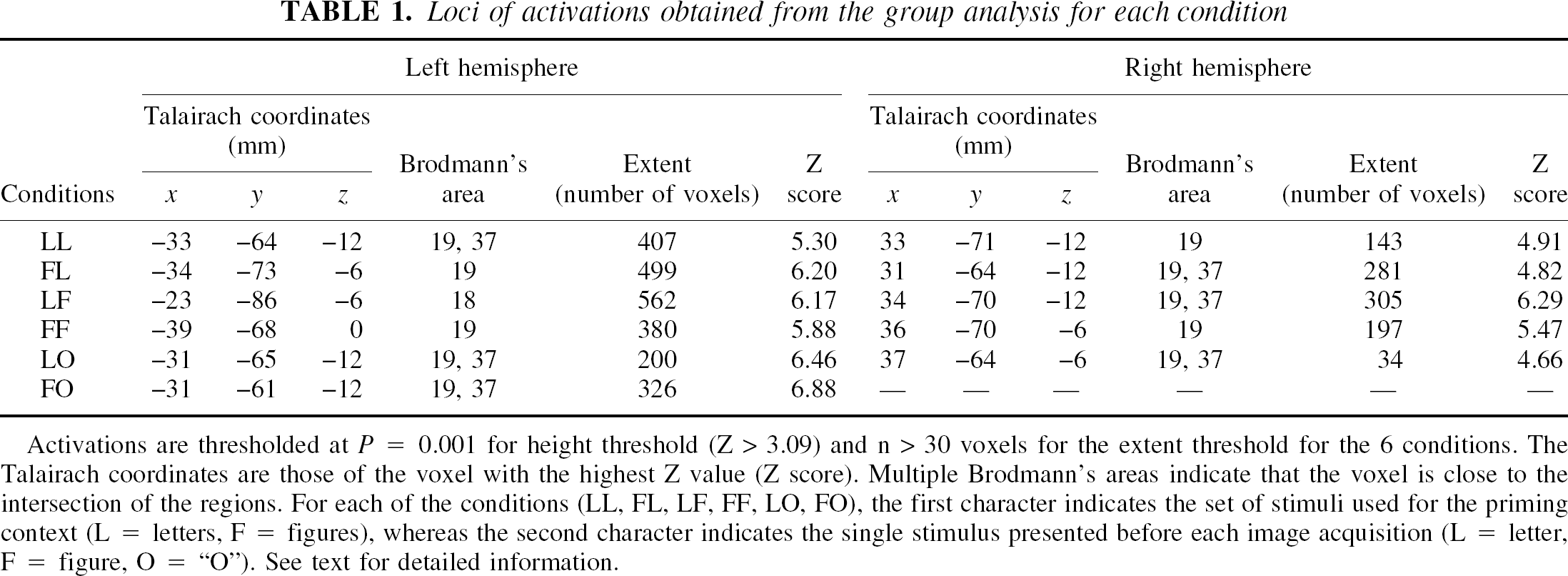
Activations are thresholded at P = 0.001 for height threshold (Z > 3.09) and n > 30 voxels for the extent threshold for the 6 conditions. The Talairach coordinates are those of the voxel with the highest Z value (Z score). Multiple Brodmann's areas indicate that the voxel is close to the intersection of the regions. For each of the conditions (LL, FL, LF, FF, LO, FO), the first character indicates the set of stimuli used for the priming context (L = letters, F = figures), whereas the second character indicates the single stimulus presented before each image acquisition (L = letter, F = figure, O = “O”). See text for detailed information.
The activations are distributed between z = −18 mm and z = 0 mm and are located in the Brodmann areas 19, 37, and 18 for the 6 conditions, that is, in the posterior fusiform gyrus, in the inferior occipital gyrus, and in the lingual gyrus. They pertained to the lateral occipital complex (LOC) defined by some authors (Malach et al., 1995; Kanwisher et al., 1997; Grill-Spector et al., 1999, 2000; Kourtzi and Kanwisher, 2000). Activations were exclusively localized in these regions, and the authors verified that lowering the Z threshold to 0.01 did not reveal any new distant cluster.
For letters and figures, bilateral activations in these regions are observed regardless of condition. However, left hemisphere dominance appears in the four conditions (LL, FL, LF, and FF). The ambiguous stimulus elicited less extensive activations, with a pronounced asymmetry in favor of the left hemisphere for both the letter and the figure priming contexts. In particular, the FO condition shows activation in the left hemisphere only (Table 1 and Fig. 3).
When the stimulus pertained to the set used for priming (congruous conditions LL or FF), the extent of activation was clearly reduced compared with that observed in the incongruous conditions (FL or LF) in both hemispheres. A reverse priming effect across hemispheres can be suspected for the ambiguous stimulus, because in the left hemisphere the extent of activation decreased with the letter priming, as observed with the letters, whereas it slightly increased in the right hemisphere, as observed with the figures. As a whole, data presented in Fig. 3 and Table 1 seem to indicate a priming × stimulus × hemisphere interaction.
Individual analysis
Figure 4 shows the activations obtained in the subject taken as reference for the coregistration. Significant activations are found in the areas revealed by the group analysis. Outside areas in the occipital pole, the only consistent activation was located, for the LO condition and for 3 of the subjects, in areas typically activated by phonologic and lexical-semantic tasks (Brodmann areas 22, 42, and 6/44 in the left hemisphere).
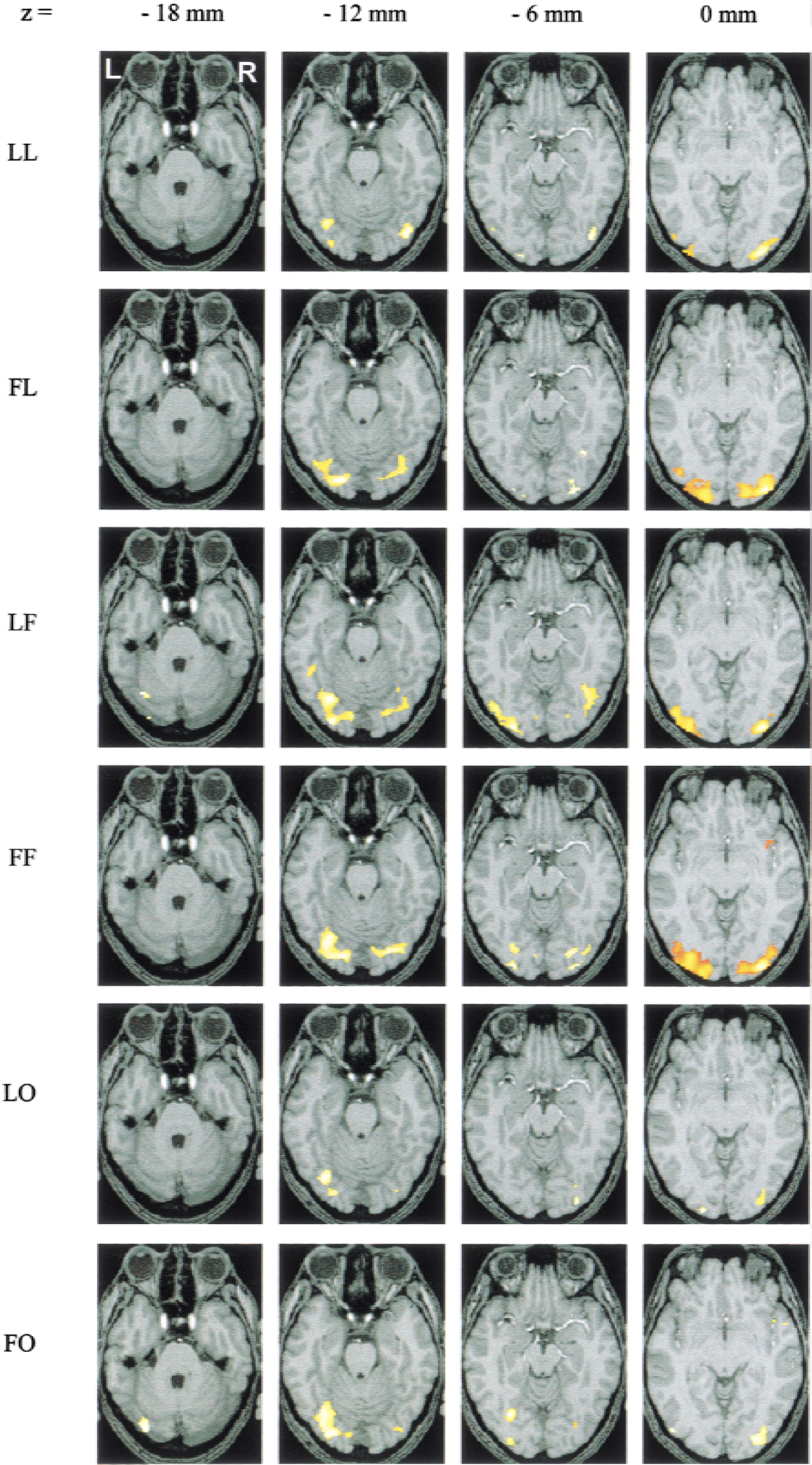
Individual activations of the subject taken as reference for coregistration. Activations are thresholded at P = 0.01 for height threshold (Z > 2.33) and at n > 10 voxels for extent threshold for the 6 conditions. See text for explanation of abbreviations.
Analysis of spatial extent.
The MANOVA on the spatial extent revealed no significant main effect of priming (F(1,5) = 0.35, P = 0.58), of stimulus (R(2,4) = 0.89, P = 0.48), and of hemisphere (F(1,5) = 2.12, P = 0.21).
As suspected from the group analysis, the priming × stimulus × hemisphere interaction (Fig. 5) was significant (R(2,4) = 7.54, P = 0.044). With the letter stimuli, the extent of activation in both hemispheres was larger in the incongruous condition (FL) than in the congruous condition (LL). Similarly, with the figure stimuli, the extent was larger in the incongruous condition (LF), but in the left hemisphere only, whereas the right hemisphere seemed less sensitive to priming effects. For the ambiguous conditions, opposite effects were observed in the left and right hemisphere. Indeed, the extent of activation strongly decreased in the left hemisphere for the LO condition compared with the FO condition, whereas the reverse was true in the right hemisphere. Although the priming effect was not significant for each of the hemispheres tested separately, the priming × hemisphere interaction was significant (F(1,5) = 8.07, P = 0.036).
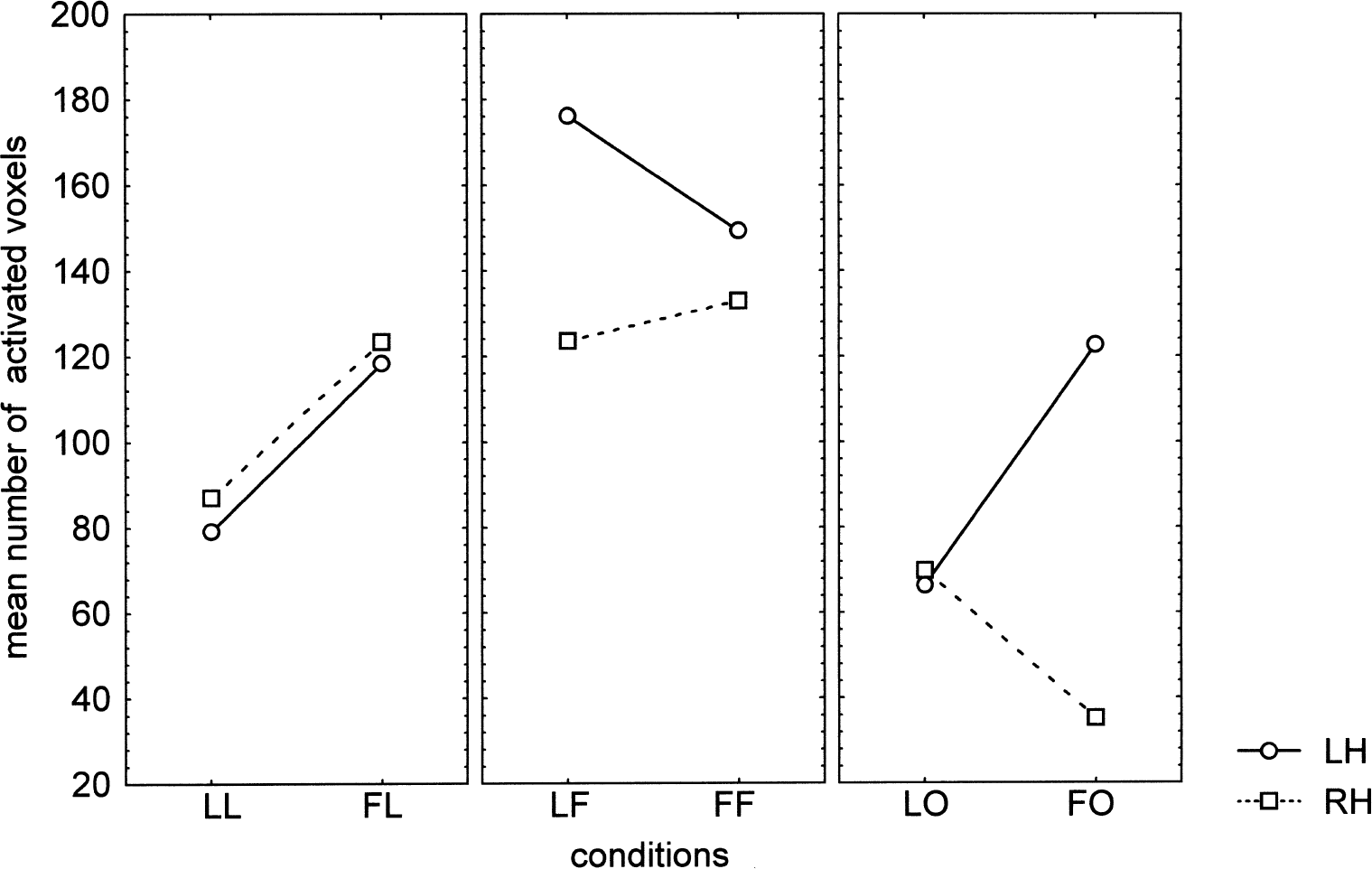
Priming × stimulus × hemisphere interaction on spatial extent (R(2,4) = 7.54, P = 0.044). The extent of activation is larger in the incongruous conditions (FL and LF) than in the congruous conditions (LL and FF), except for the right hemisphere with figures. For the ambiguous conditions, opposite effects are observed in the left and right hemisphere. See text for explanation of abbreviations.
Analysis of signal intensity.
Before processing the data on signal intensity, the authors verified for each individual that the maximum Z scores in the ROIs defined from the independent data set were again located in BA 19, 37, or 18 (Talairach coordinates: × ranging from 21 to 39, y from −51 to −87, z from 0 to −12). This demonstrates the good reproducibility of the activations obtained with the two data sets (compare with coordinates in Table 1), despite the small number of items per subject.
Figure 6 shows the time course of the percent signal change in the ROI defined from the localizer scans, averaged per hemisphere in the six subjects and for each condition. At the peak response that occurred, as expected, at time point 2—that is, 4 seconds after the beginning of the stimulus—clear differences in amplitude between conditions can be seen, showing the high sensitivity of the technique, even when using few events per subject.
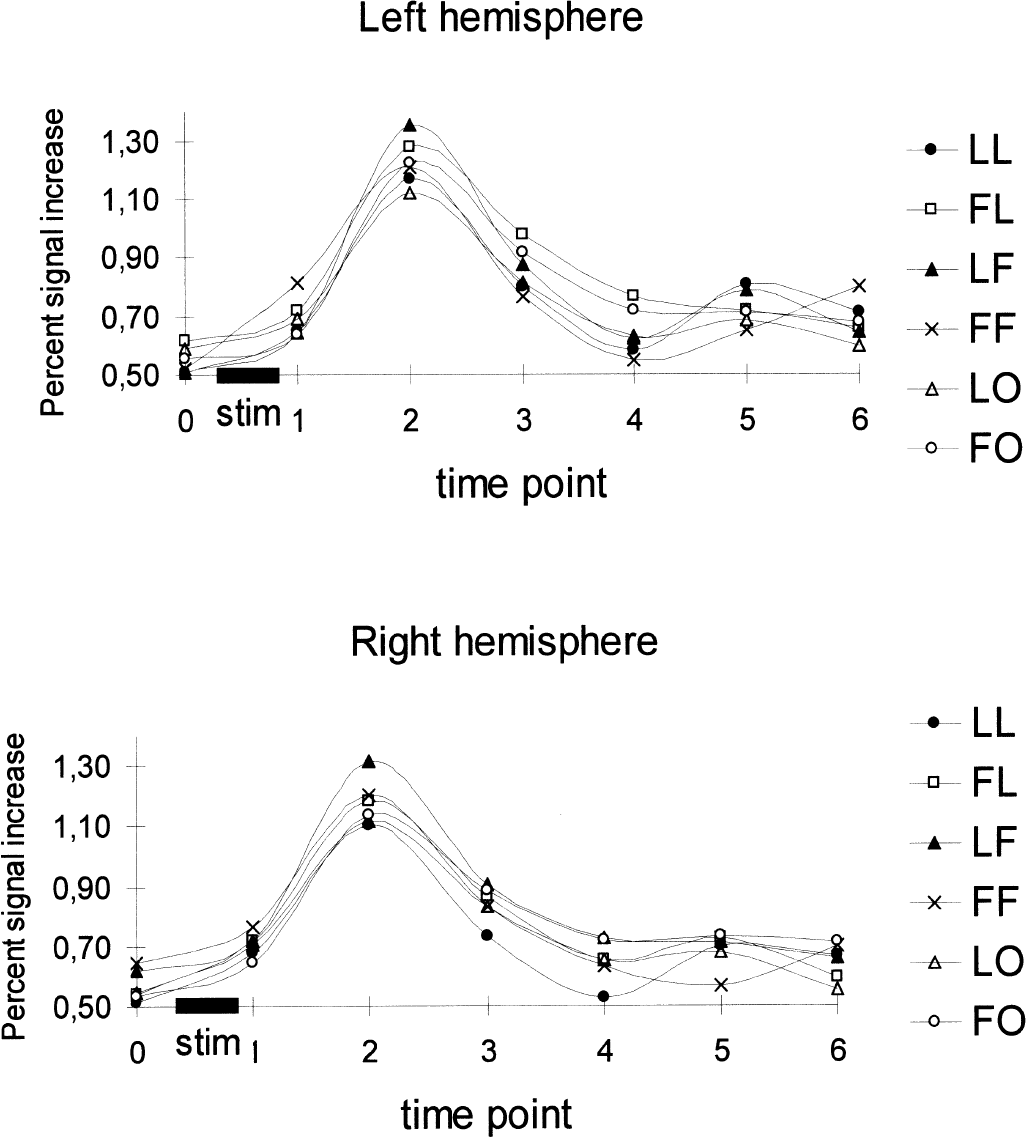
Averaged (n = 6) time course of percent signal change in the region of interest defined from the localizer scans. Individual values at time point 2 (peak response) were entered in the multivariate analysis of variance (see Fig. 7). See text for explanation of abbreviations.
The MANOVA on the peak response showed no main effect of priming context (F(1,5) = 0.07, P = 0.81) and of hemisphere (F(1,5) = 0.57, P = 0.49), but revealed a main effect of stimulus (R(2,4) = 9.18, P = 0.032). As indicated by the corresponding planned comparison, most of this main effect originated from the contrast between the ambiguous stimulus “O” and both letters and figures, whatever the prime and the hemisphere (F(1,5) = 22.95, P = 0.0049), whereas the difference between letters and figures was not significant (F(1,5) = 1.64, P = 0.26) (Fig. 7).
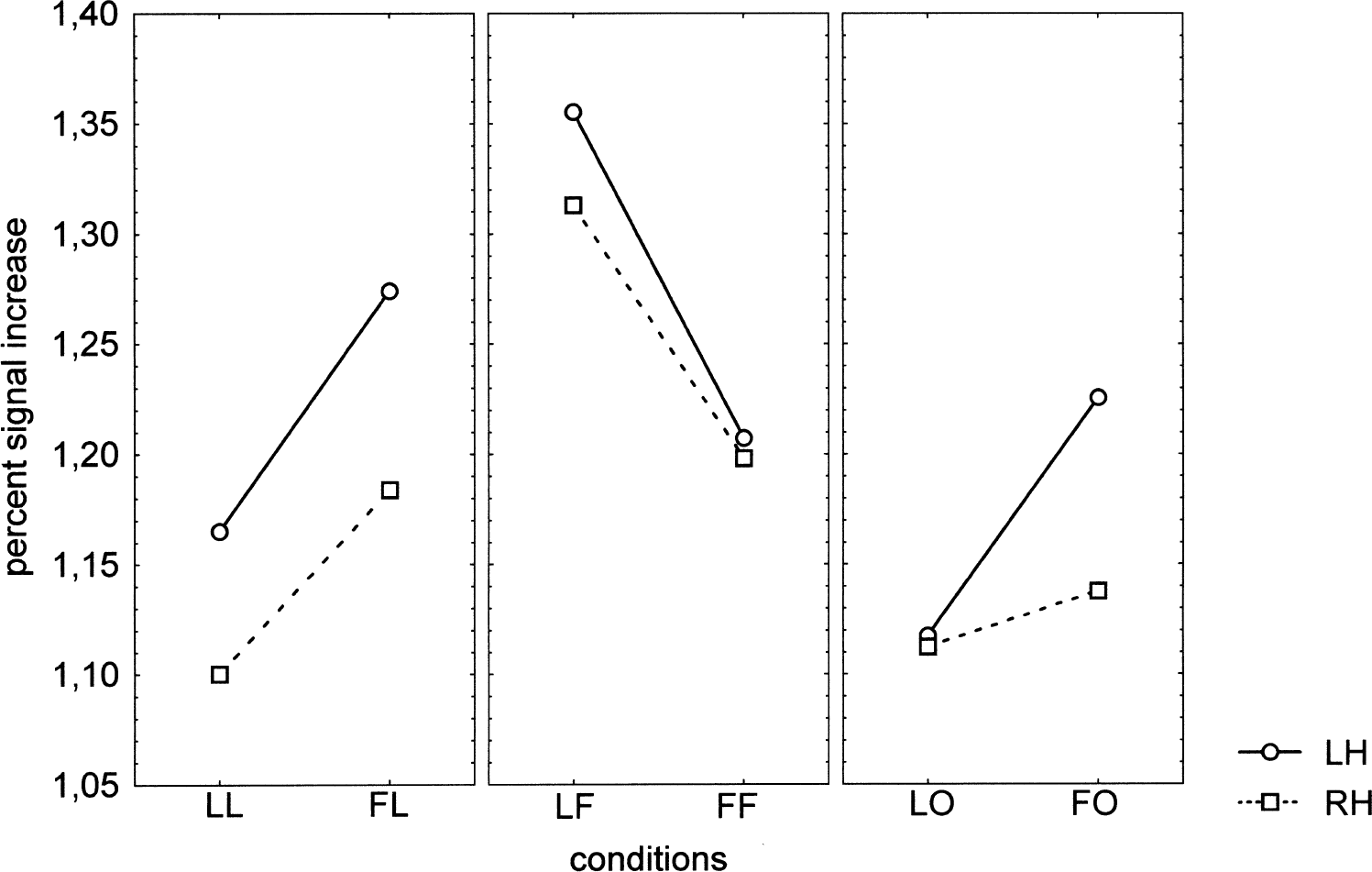
Priming × stimulus × hemisphere interaction on the peak response (R(2,4) = 14.25, P = 0.015). In comparison with congruous conditions, the incongruous conditions increased the signal intensity. The ambiguous stimulus “O” behaved as letters for the left hemisphere.
Similar to what was observed for spatial extent, the priming × stimulus × hemisphere interaction was significant (R(2,4) = 14.25, P = 0.015). As shown in Fig. 7, the incongruous conditions (FL and LF) increased the signal intensity, just as they enlarged the activated areas, for letters and familiar geometric figures. Clearly, the ambiguous stimulus “O” behaved as letters for the left hemisphere, with increased signal when primed by geometric figures (F(1,5) = 8.1, P = 0.036). In the right hemisphere, no priming effect was observed (F(1,5) = 0.31, P = 0.60), and the priming × hemisphere interaction for the ambiguous conditions (LO and FO) did not reach significance (F(1,5) = 3.81, P = 0.11).
DISCUSSION
In the current study, context effects revealed differences between the left and right extrastriate response to the passive viewing of a physically identical but categorically ambiguous stimulus that pertains to both letters and familiar geometric figures. In particular, the current results suggest that for this ambiguous stimulus, letter processing at an abstract level systematically occurred in the left LOC, whereas distinct processes were engaged in the right hemisphere. To account for the current data, involvement of the extrastriate cortex in visual shape analysis, especially orthographic processing, and in perceptual or conceptual priming should be considered.
Orthographic versus nonorthographic processing
Following Petersen et al. (1990) who showed that the left extrastriate cortex was activated by visually presented words and pseudo-words, but not by unpronounceable letter strings or false-font strings, a number of functional neuroimaging studies have tried to better delineate the role of the visual cortex in reading processes, particularly in orthographic and visual word form processing. All these studies reported activations of the extrastriate cortex consistent with the bilateral activations in the middle-posterior fusiform gyrus and lateral occipital cortex observed with single letters and familiar geometric figures in the current study. However, different activation patterns have been found when contrasting linguistic with nonlinguistic stimuli, particularly in terms of hemispheric preponderance. Greater activation in the left than in the right occipitotemporal cortex has been reported with letter strings (Price et al., 1996; Puce et al., 1996) and also with single Kanji characters (Uchida et al., 1999). Whether such a left predominance is related to orthographic processing is not clear as Pugh et al. (1996) found symmetrical activation when contrasting a purely orthographic (letter case) difference. Moreover, the Kanji morphograms used by Uchida et al. (1999) had a specific meaning and were opposed to scrambled, meaningless, and obviously less structured characters. In the study by Fujimaki et al. (1999), activations canceled when contrasting single real and pseudo-characters or strings of real and pseudo-characters, but a significant greater activation in the left occipitotemporal region was observed for strings compared with single stimuli. String length effects in visual cortex also were reported by Indefrey et al. (1997) and Tarkiainen et al. (1999), but the effects were bilateral. In the latter study, however, the high temporal resolution of magnetoencephalography allowed the authors to distinguish sources that responded in a bilateral identical manner to letter strings and symbol strings from sources that showed greater left activation with letter strings than with symbol strings of equal length. In line with the results of Tarkiainen et al. (1999), a recent study by Cohen et al. (2000), taking advantage of using both fMRI and event-related potentials to assess the response to hemifield presentation of words, showed early hemifield-dependent activations in the inferior occipitotemporal region, distinct from a strictly left lateralized and more anterior region that responded lately and independently of the hemifield of presentation.
Although the current results showed no significant hemisphere main effect, a left preponderance in extent and intensity was observed, but with both letters and familiar geometric figures. The lack of clear left advantage for letters may be explained by the use of single characters and not strings or words, and also by taking into account a familiarity effect, as suggested by Tagamets et al. (2000). The current simple geometric figures were indeed almost as familiar as letters, which is not the case of false-font stimuli. In addition, they were as spatially structured as letters and differed in this respect from textures or scrambled stimuli. If, as suggested by Garrett et al. (2000), the left temporal occipital cortex is involved in automatic letter recognition, it may be the case that such automatic processes were triggered as well by the current familiar geometric figures, giving a moderate left overall preponderance for both kinds of stimuli, possibly related to a left specialization for symbolic encoding of visual form information (Shen et al., 1999).
Early visual feature extraction versus shape processing and object recognition
Evidence on the role of the lateral occipital cortex and fusiform gyrus in shape analysis and object recognition has been obtained from recent studies. Higher bilateral activations were found with structured object than with noise, texture, or scrambled stimuli (Malach et al., 1995; Martin et al., 1996; Kanwisher et al., 1997; Grill-Spector et al., 1998; Doniger et al., 2000; Kourtzi and Kanwisher, 2000). By varying the exposure duration of masked objects, Grill-Spector et al. (2000) found a correlation between recognition performance and fMRI signal in the LOC. These activations appear largely independent of differences in visual cues (Okusa et al., 2000) and not only reflecting low-level visual processes, as shown by Op de Beeck et al. (2000) who found increased activation in the posterior fusiform gyrus when contrasting object categorization with nonsense object orientation judgement. In the current study, the bilateral activation found in this region after passive viewing of letters and simple geometric figures thus might reflect automatic recognition of familiar objects.
Although the reported activations were systematically bilateral, some results indicate that the level of familiarity and meaning may induce interhemispheric difference (Schacter et al., 1995; Kanwisher et al., 1997; Beason-Held et al., 1998; Moore and Price, 1999). A possible interpretation of these results would consider that recognition of novel object increases the demand on on-line processing for feature extraction and shape analysis predominantly in the right extrastriate cortex, whereas recognition of known objects or visual patterns would rely on more left-sided processes. This would explain why Shen et al. (1999) found a strong left hemisphere dominance in a task that involved repeated comparisons of consecutive stimuli chosen among only four visual patterns. Accordingly, the left preponderance observed in the current experiment might result from using very familiar, overlearned stimuli.
Perceptual priming: specific and abstract visual form effects
The activations obtained after presentation of the “O” were weaker, both in signal intensity and extent, than those following the other stimuli, particularly in the right hemisphere. This may result from the higher repetition rate of the ambiguous stimuli all along the experiment. Indeed, stimulus repetition has been shown to decrease the level of activation in the inferior occipitotemporal region bilaterally (Buckner et al., 1995, 1998; Buckner and Koutstaal, 1998; Grill-Spector et al., 1999), but with a right predominance (Badgaiyan and Posner, 1997; Beason-Held et al., 1998; Doyle and Rugg, 1998; Backman et al., 2000; Henson et al., 2000) that might be related to less demand on visual feature extraction and form processing, especially for familiar stimuli. A result that may appear contradictory to the decrease in activation observed in the current study and in the works quoted above is the increased activation in the inferior temporal regions reported by Dolan et al. (1997) in their PET study of rapid perceptual learning. This apparent discrepancy can be explained by the stimuli and experimental design used by the authors. They measured the response to degraded images of unknown complex stimuli (faces or objects in natural environment) before and after exposure to the undegraded version of the images (explicit learning phase). It is likely that such an experimental design induced intense on-line processing for image completion at the second presentation that resulted in enhanced activity in extrastriate regions. However, as in the current study the “O” differed from any other stimulus by one or two salient visual features (no angle, closed contour), it cannot be excluded that the overall decrease in activation results from a difference in visual processing related to increased salience and is independent of repetition priming.
In addition, the current study shows that context effects modulate the response of the extrastriate cortex to the passive viewing of visual stimuli. The decrease in activation observed for letters and familiar geometric forms after exposure to a congruent set of stimuli may be first explained, as for the “O”, by a direct repetition priming effect. Indeed, in the congruous conditions, each item was presented two times during the context phase before image acquisition. This repetition effect may however involve more abstract form representations as it has been shown to be insensitive to changes in visual cues and viewing conditions (Grill-Spector et al., 1999; Kourtzi and Kanwisher, 2000). Accordingly, Op de Beeck et al. (2000) obtained evidence that activation of the fusiform gyrus, at least in its anterior part, merely reflects proximal stimulus representation rather than part-based or global form analysis. Including such high-level description of the stimulus in the processes performed in the LOC appears necessary to account for the difference in context effects observed in the current study with the ambiguous stimulus. Indeed, the context effects cannot be explained by the repetition of the stimulus; in both letter and figure contexts, the “O” was presented two times and the adaptation caused by repetition priming should have equally decreased the response. Yet, letter context diminished the level of activation in the left hemisphere compared with figure context, in terms of both signal intensity and spatial extent. In the right hemisphere, a decrease in activation also was observed for spatial extent, but in the figure context, whereas the context effect on signal intensity was less clear. Thus, not only did the authors observe a context effect even though the rate of repetition was identical, but this context effect appeared hemisphere-dependent.
These results might reflect different processes involving two relatively independent visual form subsystems as proposed by Marsolek and colleagues: a specific visual form subsystem (SVF) representing form-specific information and mainly mediated by the right hemisphere (Marsolek et al., 1992, 1996; Burgund and Marsolek, 1997; Doyle and Rugg, 1998; Vaidya et al., 1998), and an abstract visual form subsystem that matches the stimulus to some abstract category of form and operates more effectively in the left hemisphere (Marsolek, 1995; Burgund and Marsolek, 1997). Interpreting our results within this framework would lead to the assumption that incongruous priming increased activation in the right hemisphere as the result of increased demand on the specific visual form subsystem that processed the differences in visual features between letters and geometric forms, possibly on an holistic basis (Marsolek et al., 1996). This also would account for the relative increase in spatial extent of the right activation observed with the “O” in letter context, as its global visual form is more akin to that of the geometric forms than to that of letters. In the left hemisphere, the abstract visual form would have performed easier abstract form categorization in congruous priming conditions that resulted in decreased activation under these conditions. In this respect, the ambiguous stimulus “O” would have been preferentially ascribed to the abstract form category of letters or mapped onto an abstract letter code by the left hemisphere (Bowers, 2000). Supporting the existence of a neural substrate for the abstract category of letters are the data reported by Polk and Farah (1998) who found an area in the left inferior occipitotemporal cortex that responded more to the passive viewing of letters than digits, whereas distinct areas responded more to geometric shapes than to letters or digits. Unfortunately, because of the imaging technique, only the left hemisphere was examined in this study. Polk and Farah interpreted this finding as evidence that, in the visual system, the neural substrates underlying letter recognition are segregated from those involved in the recognition of other arbitrary categories of stimuli as the result of environmental exposure.
Conceptual priming effects
Whereas the extrastriate cortex has been consistently reported to be involved in perceptual priming, conceptual priming is considered to rely mainly on frontal regions (Cabeza and Nyberg, 2000), especially the left dorsal prefrontal cortex (Buckner et al., 1998). However, recent studies on word priming reported amodal repetition effects in the extrastriate and posterior inferior temporal cortices (Badgaiyan et al., 1999; Buckner et al., 2000; Yasuno et al., 2000), thus suggesting that these regions may support higher level, more conceptual processes, or at least are modulated by top-down control (Nobre et al., 1998; Takeuchi et al., 2000).
Although the current experiment was based on passive viewing that did not require a specific task, the possibility can not be ruled out that the context modulation of the extrastriate response observed, particularly in the left hemisphere with the ambiguous stimulus, is partly related to conceptual rather than perceptual processes, such as access to lexical/semantic representations used, for example, for a “verbal tagging” of the object, as suggested by Buckner et al. (2000). Such processes may have occurred notably in the LO condition in which the individual analysis showed significant activation of the left inferior frontal and superior temporal regions in three of the subjects. A particularity of the figure set compared with the letter set should be addressed here if participants were covertly naming the stimuli. Because the authors wanted to use only familiar geometric figures physically closed to letters, some of the items of the figure set only differed in orientation and had the same name, whereas all the letters had unique names. Mutual lexical priming within the figure set might have produced less activation for these items, and comparatively greater activation with the ambiguous stimulus (FO condition), especially in the left hemisphere. However, several lines of evidence can be drawn from the current data that do not support this interpretation. It can be observed first that, for the LO condition too, there is a pronounced asymmetry in favor of the left hemisphere (Table 1). Second, compared with the FF condition, the FO condition does not elicit greater activation in the left hemisphere, neither in extent (Table 1) nor signal intensity (Fig. 7). Thus, the “O” does not appear to be less primed than the other geometric figures by the figure context. However, this finding may result from the higher repetition rate of the ambiguous stimulus. A way to eliminate pure repetition effects is to compare the letter conditions (LL and FL) with the figure conditions (LF and FF), because in these conditions that exclude the “O”, each stimulus was repeated exactly the same number of times. In these conditions, activations elicited by the geometric figures are always higher than those elicited by letters (the priming context and the hemisphere being fixed), giving little support to an increased labeling effect with the geometric figures. In addition, Fig. 7 (third panel) shows a marked decrease in the LO condition (not a marked increase in the FO condition), as FO activation is as high as FF activation in the left hemisphere (second panel), despite the higher repetition rate for the “O” than for any other figure. Finally, a naming account does not accommodate the adaptation effect in the right hemisphere for the ambiguous stimulus when it was primed by geometric figures (Fig. 5, third panel). As this adaptation cannot be explained by a pure repetition effect, an abstract category effect, as suggested above, remains likely.
At the conceptual rather than perceptual level, the priming × hemisphere interaction observed with the ambiguous stimulus may also have resulted from different, hemisphere-dependent modes of processing. In a study based on ERP recordings, Federmeier and Kutas (1999) have shown that although both hemispheres are sensitive to the context elicited by the meaning of a sentence, only the left hemisphere is sensitive to semantic similarity between expected and unexpected ending. They suggest that the left hemisphere proceeds according to a predictive mode that compares the upcoming stimulus with what is expected, whereas the right hemisphere compares the stimulus directly with context information. Recording the ERPs elicited by an ambiguous stimulus presented in either speech or nonspeech context, Celsis et al. (1999) found an N2b component in the response recorded on the left occipitotemporal region in the nonspeech context. As this component has been shown to be insensitive to acoustic differences and is considered to reflect the detection of incongruity or template mismatch, they suggested that the ambiguous phoneme was processed as an unattended stimulus when presented among environmental sounds, possibly because a phonemic representation was maintained as the result of automatic, modular perceptual mechanisms. In line with the above interpretations, it may be that the ambiguous stimulus “O” was expected and easily matched to its “alphabetic” representation in the left LOC when presented after letter context, leading to decreased activation in comparison with figure context. However, the authors must acknowledge that their main result supporting such an interpretation has been obtained with only one letter of the alphabet, the letter “O”. Caution should be exercised when interpreting the results as the authors can not exclude that their finding might be due, at least in part, to a particular perceptual status of this unique stimulus.
Conclusion
The high sensitivity of event-related fMRI coupled with repetition priming and category-cued priming of an ambiguous stimulus allowed the authors to show that the visual system processes letters differently from other familiar stimuli, and that the left lateral occipital cortex may be specifically involved in the abstract categorization of alphabetic symbols.
Footnotes
Acknowledgments:
The authors thank all of the volunteers for participating in the study, and the technicians of the MRI Unit, Department of Neuroradiology, for technical assistance.