
Abstract
The neuropeptide corticotropin releasing hormone (CRH) is the central nervous system (CNS) transducer of stressful stimuli. Endogenous CRH is released from neuronal terminals in several central nervous system regions—for example, amygdala and hypothalamus—during stress, and exogenous CRH administration mimics stress-related behaviors and hormonal patterns. However, whereas the role of endogenous CRH as a stress neuromodulator has been established, recent findings suggest that the peptide also functions to influence cognitive, emotional, and neuroimmune functions by modulating neuronal communication in a number of circuits. Although anatomic and pharmacologic approaches have provided evidence for this wider spectrum of CRH actions, the discrete regions and specific circuits activated by CRH have not been fully elucidated. In this article, the authors report on the use of two complementary methods to discern specific regions and cell groups activated by the administration of CRH. Glucose metabolism analysis provided quantitative measures of CRH-induced activation, but at a regional resolution; expression of the immediate early gene c-fos permitted a single cell resolution, but underestimated the neuroanatomic extent of CRH-induced activation. Overlapping regions activated using both methods delineated discrete cortical, limbic, and motor pathways. Importantly, cell groups activated by CRH included those possessing either or both members of the CRH receptor family, suggesting that both receptors may mediate the effects of the endogenous ligand. In summary, CRH activates a broad but selective array of neuronal structures belonging to cortical, limbic, and motor circuits. These findings indicate that stress-related release of this peptide may contribute to a spectrum of important modulations of CNS function.
The neuropeptide corticotropin releasing hormone (CRH) is a key central nervous system (CNS) transducer of stressful stimuli (Vale et al., 1981). Stress-induced release of CRH from hypothalamic nerve terminals augments the release of adenocorticotropic hormone to stimulate glucocorticoid secretion from the adrenal cortex (Vale et al., 1981; Baram et al., 1997a). Outside the hypothalamus, CRH is expressed in neurons and terminals throughout the neocortex and in selected limbic and brainstem regions (Swanson et al., 1983; Sakanaka et al., 1986). This anatomic distribution has suggested that CRH may have effects in addition to its well-established neuroendocrine function (Koob et al., 1993; Behan et al., 1995; Baram and Hatalski, 1998). Thus, CRH administration into the cerebral ventricles (icv) reproduces stress-induced behaviors (Koob et al., 1993), CRH antagonists block behaviors elicited by stressful stimuli (Swiergiel et al., 1993) and release of the peptide in amygdala during stress has been established (Merali et al., 1998). These observations indicate a key role for CRH in the behavioral and neuroendocrine responses to stress (Koob et al., 1993; Swiergiel et al., 1993; Baram et al., 1997a).
Evidence for additional roles for CRH in the CNS has been mounting, implicating the peptide in the mechanisms of learning and memory (Behan et al., 1995), food intake homeostasis (Gardner et al., 1998), febrile responses (Cartwell et al., 1999), and neuronal injury (Baram and Hatalski, 1998). The cellular mechanisms of these diverse effects of CRH have remained a topic of active investigation: electrophysiologic studies have shown that CRH selectively activates neuronal populations in regions including amygdala, hippocampus, locus coeruleus, and cerebellum (Baram and Hatalski, 1998). Indeed, neurotransmitter function of endogenous CRH has been demonstrated in locus coeruleus (Valentino et al., 1983) and inferior olive. Although physiologic release of CRH in cortical and limbic synapses may promote processes of learning and memory (Lee et al., 1992; Behan et al., 1995), larger amounts of exogenously administered CRH increase limbic neuronal excitation leading to seizures (Ehlers et al., 1983; Baram et al., 1992, 1997b), and may participate in mechanisms of neuronal injury (Baram and Hatalski 1998). The excitant actions of CRH are particularly rapid and prolonged in developing rat (Baram et al., 1992), where the peptide has been suggested to mediate certain triggered seizures (Baram and Hatalski, 1998).
The postsynaptic mechanisms by which CRH influences neuronal function involve activation of specific receptors. Two general CRH receptor types, CRF1 and CRF2, have been characterized (Potter et al., 1994; Chalmers et al., 1995). These are found in largely nonoverlapping distributions, suggesting that they may mediate discrete functions of CRH (Potter et al., 1994; Chalmers et al., 1995; Avishai-Eliner et al., 1996; Eghbal-Ahmadi et al., 1998). For example, neuroendocrine and proexcitatory effects of CRH may be mediated by the CRF1 receptor type (Baram et al., 1997b). The function(s) of the second receptor, CRF2, has not been fully characterized; recent evidence for its roles in stress-related and homeostatic effects of CRH has been emerging (Eghbal-Ahmadi et al., 1999). In addition, whether CRH constitutes the native endogenous ligand activating CRF2 has remained a focus of debate (Vaughan et al., 1995).
Although the postsynaptic targets of CRH have been a topic of intense study, the specific neuronal pathways mediating CRH effects have not been clarified. For example, although in vivo electrophysiologic approaches using electrodes implanted in limbic structures have demonstrated that CRH-induced neuronal activation originates in amygdala and spreads to hippocampus and neocortex (Ehlers et al., 1983; Baram et al., 1992), no CRH-expressing projection neurons have been found that can convey direct CRH-mediated neurotransmission from amygdala to hippocampus (Swanson et al., 1983). In addition, the extent of the neuronal circuitry activated by CRH cannot be determined from EEG recordings limited to the regions of electrode placement, or by selective injections of the peptide to specific sites (Cartwell et al., 1999).
The goal of the current study was to use two independent and complementary measures of neuronal activation to answer the following questions: what neuronal pathways and structures are activated by intra-CNS CRH administration? and what are the relations between CRH-activated neuronal circuits and the distribution of each of the CRH receptors? These data should permit, and may be a prerequisite for, full understanding of the mechanisms by which CRH exerts the spectrum of its effects on the CNS.
MATERIALS AND METHODS
Experimental design
The overall strategy of these studies was to compare glucose utilization rates ([14C]2-deoxyglucose quantitative autoradiographic method) and the expression of the immediate early gene (IEG) c-fos between control animals and those subjected to CRH infusion. Potential diurnal variability in CRH receptor occupancy or neuronal activation was addressed by infusing CRH between 8 am and 10 am. Rats were subjected to surgery the day before infusion of the neuropeptide and were returned to the animal facility overnight. All experimental procedures were approved by the Institutional Animal Care Committee and conformed to the rules of the European Committee Council Direction of November 24, 1986 (86/69/EEC), the French Department of Agriculture (License No. 00733) for the [14C]2-deoxyglucose (2DG) study, and the National Institutes of Health guidelines for c-fos experiments.
Animals
These experiments were performed on developing rats (postnatal, 10 to 12 days) for several reasons: (1) previous work from the laboratory has characterized the physiologic and pathologic actions of CRH during this age (Baram et al., 1992, 1997b; Baram and Hatalski, 1998); (2) the quantitative methods for circuit activation, that is, regional glucose use and the time-course of c-fos expression, have been previously defined for this age (Nehlig et al., 1988; Fernandes et al., 1999); (3) some actions of CRH, particularly excitatory modulation of amygdala and hippocampal neurons, are particularly prominent during the second postnatal week in the rat (Baram and Hatalski, 1998), potentially partially because of the ontogenic profile of the CRH receptors (Pihoker et al., 1992; Avishai-Eliner et al., 1996; Eghbal-Ahmadi et al., 1998). Overall, 33 immature rats participated in these experiments.
For metabolic studies, 5 rats were subjected to CRH infusion and 4 served as controls. For c-fos mRNA in situ hybridization, stress-free controls (n = 6) and those subjected to vehicle (n = 7) were compared with animals infused with CRH (n = 11). For the 2DG experiments, animals were offspring of Sprague Dawley rats purchased from Janvier Breeding Center, Le Genest-St-Isle, France. For the c-fos study, rats were offspring of the same strain, purchased from Zivic-Miller, Zelienople, PA, U.S.A. Dams were housed in approved, uncrowded animals facilities under controlled temperature and a 12-hour light–dark cycle with unlimited access to lab-chow and water. Cages were inspected for presence of pups at 12-hour intervals, and date of birth was considered day 0.
Surgical procedures and CRH administration
For CRH infusion, pups were implanted with stainless steel cannulae into the left lateral cerebral ventricle on the daypreceding the experiments, as described in detail elsewhere (Baram et al., 1992; stereotaxic coordinates, using Bregma as landmark: AP −0.7, L 2.0, V 3.3). Cannula position was verified in all cases. For the 2DG study, animals underwent placement of catheters in addition to cannulae implantation as described in detail previously (Nehlig et al., 1988). Briefly, a femoral artery and vein were catheterized with polyethylene catheters under light halothane anesthesia. Both catheters were threaded under the skin up to the back of the hind paw to allow free access to the catheters without disturbing rat movements (Nehlig et al., 1988). Peptide infusion, or the infusion of an equal volume of vehicle, was performed on postnatal days 10 to 12 to freely moving pups using a micropump (Baram et al., 1992, 1997b).
The dose of CRH (0.75 nmol in 1.3 μL) was selected based on previous dose response studies (Baram et al., 1992, 1997b). The dose used in the current study has been shown to lead to activation of select and specific neuronal populations (measured behaviorally and by EEG using depth electrodes in multiple brain regions). In addition, all of the effects of this dose of CRH have been entirely prevented using nonselective antagonists of CRH that block CRF1 and CRF2 receptors. The activatory effects of this CRH dose in limbic regions have also been selectively blocked by nonpeptide, selective antagonists of the CRF1 receptor subtype (Baram et al., 1997b).
Determination of cerebral glucose utilization rates
CMRglcs were measured by the [14C]2-deoxyglucose technique (Sokoloff et al., 1977) adapted to immature rats (Nehlig et al., 1988). The [14C]2DG (4.625 Mbq/kg, specific activity 1.65 to 2.04 GBq/mmol; Isotopchim, Ganagobie-Perhuis, France) was injected as an intravenous bolus to freely moving rats 30 minutes after the onset of CRH-induced behavioral activation (Ehlers et al., 1983; Baram et al., 1992, 1997b) and 40 minutes after the infusion of vehicle for the control rats. Timed arterial blood samples were drawn over the next 45 minutes for the measurement of plasma glucose and 2DG concentrations. Animals were killed by decapitation 45 minutes after tracer injection, brains were rapidly removed, frozen using isopentane, and cut into 20-μm coronal sections. Sections were apposed to Amersham Biomax MR films for 10 days, along with [14C]-methylmethacrylate standards calibrated for their 14C concentration in brain tissue. The autoradiograms were digitized and analyzed by densitometry using an image-processing system (Biocom 500, Les Ulis, France). The localization of specific nuclei was assessed on adjacent sections stained with cresyl violet according to the developing rat brain atlas of Sherwood and Timiras, 1970. CMRglcs were calculated according to the operational equation of the 2DG method using the usual rate and lumped constants (Sokoloff et al., 1977; Nehlig et al., 1988). Controlling for the site of cannula implantation, CMRglcs were determined in 47 cerebral structures of each animal, and values of CMRglcs were compared separately in sides ipsilateral and contralateral to the cannula between vehicle and CRH-infused groups using a Dunnett's t-test for multiple comparisons.
Mean arterial blood pressure was regularly recorded throughout the experimental procedure. Hematocrit was determined from blood samples collected into capillary tubes just before the isotope injection and 30 minutes later. Immediately before 2DG injection and animal death, blood samples were obtained for measurement of pH, pO2, and pCO2 using a blood gas analyzer (model 158; Corning Medical and Scientific, Halstead, U.K.).
Determination of neuronal activation using in situ hybridization of c-fos
One hour after infusion of CRH, animals were decapitated and brains were rapidly dissected onto powdered dry ice and stored at −80°C. Sections were cut coronally at 20 μm using a cryostat, mounted on gelatin-coated slides, and stored at −80°C. For the synthesis and preparation of the riboprobe, a plasmid containing a 667 base-pair fragment of c-fos cDNA (kindly provided by Dr. C.M. Gall) was linearized with Eco RI. Radioactive antisense cRNA was synthesized by incubating T7 RNA polymerase (30 U; Promega, Madison, WI, U.S.A.) with 1 μg linearized plasmid in 2.5 mmol/L ATP/GTP/UTP, 6 μmol/L [α-36S]-CTP (New England Nuclear, Boston, MA, U.S.A.), 10 mmol/L DTT, 40 mmol/L Tris-HCl (pH 7.5), 6 mmol/L MgCl2, 2 mmol/L spermidine, 10 mmol/L NaCl, and 40 U RNase inhibitor (RNASIN, Promega). After 1 hour at 37°C, RNase-free DNase (RQ1-DNase, Promega) was added for 15 minutes at 37°C. The integrity of each transcript was determined by acrylamide gel electrophoresis and the probe was subjected to alkaline hydrolysis. The probe was purified by column chromatography (Select-D[RF], 5prime-3prime, Boulder, CO, U.S.A.). The specific activity of each probe was 3 to 4 × 106 cpm/μg.
In situ hybridization procedure followed previously published protocols (Avishai-Eliner et al., 1996; Eghbal-Ahmadi et al., 1998, 1999). Briefly, sections were brought to room temperature, air-dried, and fixed in fresh 4% buffered paraformaldehyde for 20 minutes, followed by dehydration and rehydration through graded ethanols. Subsequently, sections were exposed to 0.25% acetic anhydride in 0.1 mol/L triethanolamine (pH 8) for 8 minutes and were dehydrated through graded ethanols. Sections were air-dried and prehybridized for an hour at 55°C in a humidity chamber. Sections were then hybridized for approximately 20 hours with the 35S-labeled ribonucleotide probe at 55°C. Posthybridization, sections were washed in 2 × SSC (1 × SSC denotes 0.15 mol/L NaCl, 15 mmol/L trisodium citrate buffer, pH 7) for 5 minutes at room temperature, and were digested using RNase-A (200 μg/mL; Calbiochem, La Jolla, CA, U.S.A.) for 30 minutes at 37°C. Sections were washed successively (at 55°C) in 2 × SSC and 1 × SSC for 5 minutes, 0.25 × SSC for 30 minutes, and 0.1 × and 0.03 × SSC for 1 hour each. Finally, sections were dehydrated through 100% ethanol and apposed to film (Hyperfilm β-Max, Amersham, IL, U.S.A.) for 15 days.
Semiquantitative analysis of c-fos mRNA was accomplished as described previously (Eghbal-Ahmadi et al., 1998, 1999). Films were scanned using a StudioStar scanner (AGFA; resolution 1200 × 1200 dots per inch) equipped with a transparency bed and the Fotolook 32 software. Digitized images were acquired onto a Dell computer and analyzed using the ImageTool software program (University of Texas Health Science Center, San Antonio, TX, U.S.A.). For each group 6 to 8 brains were used, 2 to 8 sections for each region per brain. Each side was analyzed separately (when possible the CRH-injected side was marked by a needle hole: values for the ipsilateral side were invariably higher than the contralateral side). For each animal, values generated for each structure were averaged. Structures analyzed for optical densities of c-fos mRNA were defined at several coronal levels (Sherwood and Timiras, 1970): A−0.4 to −0.1 mm for entorhinal cortex; A−2.0 mm for parietal cortex, posterior thalamus, and hypothalamus; A−2.3 mm for piriform cortex, medial hippocampus, medial nuclei of thalamus, and some amygdala nuclei; A−3.6 to −3.8 mm for anterior thalamus and paraventricular thalamic nuclei; A−4.4 for globus pallidus and striatum; and A−5.5 to −5.6 mm for medial septum. Densities were calibrated using 14C standards and are expressed in μCi/g (Eghbal-Ahmadi et al., 1998, 1999). The significance of measured quantitative differences between groups was evaluated using Student's t-test, with levels set at P < 0.05.
RESULTS
Effects of corticotropin releasing hormone on LCMRglc
In 10-day-old control rats, CMRglcs were in the same range as those previously reported (Nehlig et al., 1988). In rats subjected to CRH administration, CMRglcs increased significantly over control levels in widespread yet selective patterns. For the majority of structures, activation after CRH infusion was bilateral, in some ipsilateral, and in only three contralateral (Fig. 1).
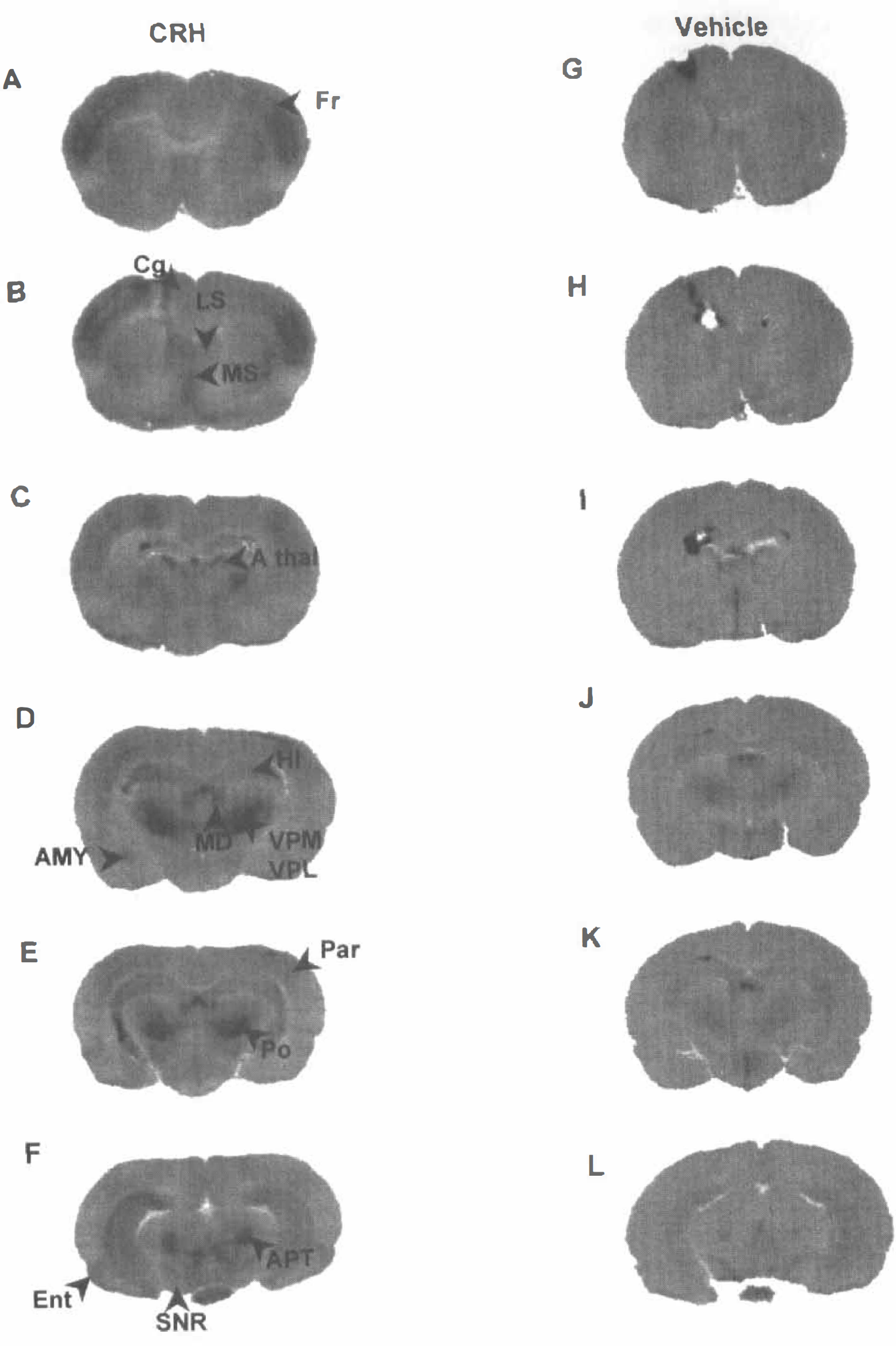
[14C]2DG autoradiographs of coronal rat brain sections at multiple levels, from animals given corticotropin releasing hormone (CRH) or vehicle into the cerebral ventricle. Note the low homogenous level of grain density in control (vehicle) sections. Coronal levels progress antero-posteriorly.
CMRglcs increased bilaterally in several cerebral cortex regions. As shown in Table 1, robust (170% to 450%) bilateral increases in glucose use were observed in frontal, cingulate, parietal, and entorhinal cortices. More modest metabolic activation was noted in primary olfactory and visual cortices. Piriform cortex was the only cortical region activated ipsilaterally only, whereas no metabolic activation was found in the auditory cortex.
Effects of CRH on LCMRglcs in the brain of P10 rats
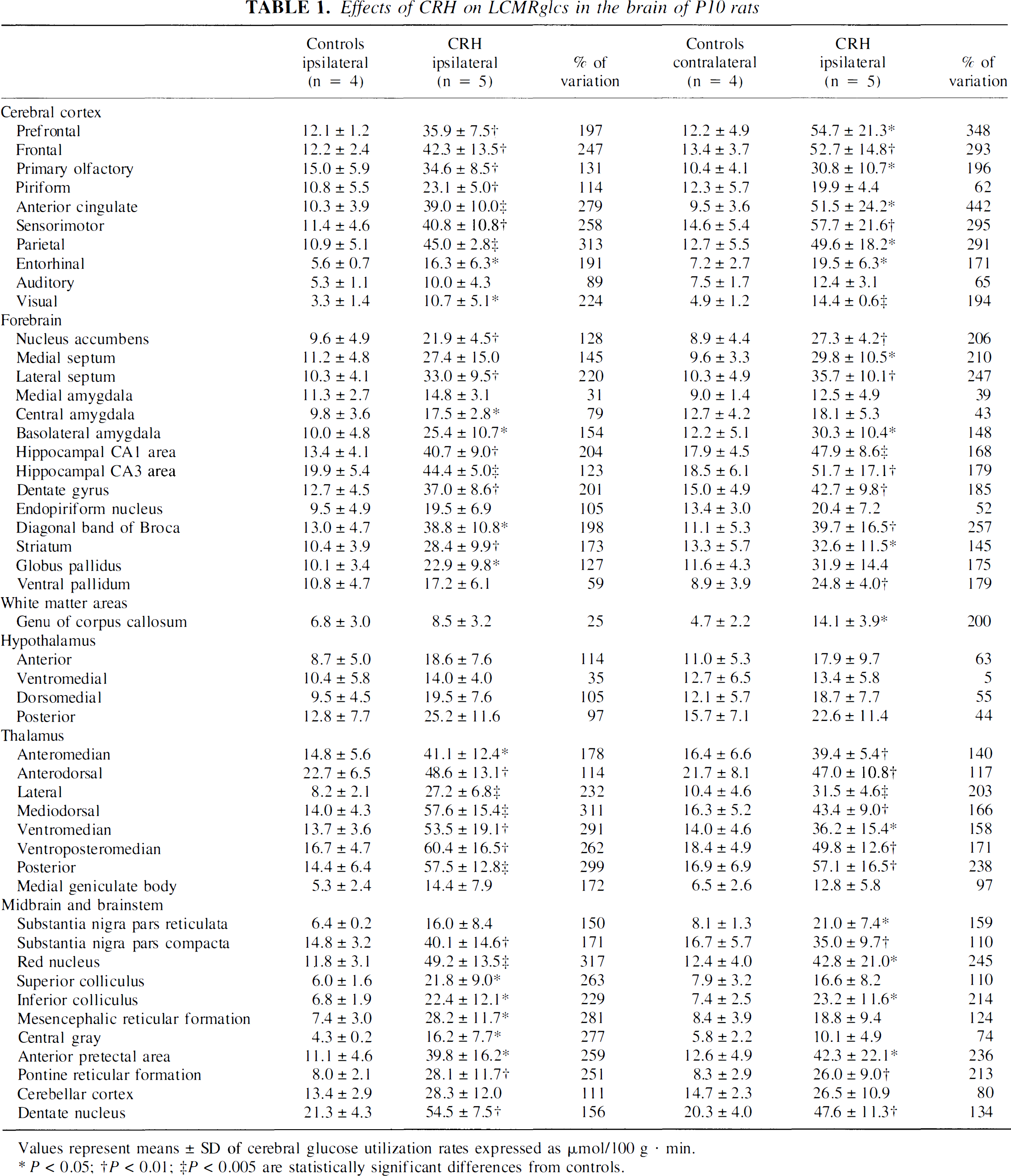
Values represent means ± SD of cerebral glucose utilization rates expressed as μmol/100 g · min.
P < 0.05;
P < 0.01;
P < 0.005 are statistically significant differences from controls.
In limbic regions, CMRglc was significantly increased bilaterally in the hippocampal formation (Table 1). In the amygdala, the basolateral nucleus was activated bilaterally, whereas the central nucleus (ACe) was activated ipsilaterally only and there was no metabolic change in the medial nucleus. Whereas the nucleus accumbens and lateral septum were activated bilaterally, the medial septum demonstrated significantly enhanced CMRglcs only on the side contralateral to CRH infusion. The most robust enhancement of cerebral metabolism within limbic structures (150% to 250%) occurred in dentate gyrus, CA1, septum, and nucleus accumbens.
In motor regions, CMRglcs increased significantly in the striatum and substantia nigra pars compacta bilaterally, whereas similar increases in substantia nigra pars reticulata reached statistical significance in the contralateral side only. Increased metabolic rates were found also in the ipsilateral globus pallidus and contralateral ventral pallidum. Most pronounced metabolic rate increases (130% to 170%) were noted in substantia nigra pars reticulata, globus pallidus, and striatum. In thalamus, significant bilateral increases of CMRglcs were found in all analyzed nuclei. Maximal increase (>300%) was observed in the ipsilateral mediodorsal nucleus. CMRglc was not increased in any hypothalamic nucleus after CRH-infusion.
In mesencephalic nuclei, large increases in CMRglcs (240% to 320%) were observed bilaterally in the red nucleus and pretectal area, and ipsilaterally in the superior colliculus, mesencephalic reticular formation, and central gray. Metabolic activity was not affected by CRH in the endopiriform nucleus, medial geniculate body, and cerebellar cortex.
Effects of corticotropin releasing hormone on c-fos expression
C-fos mRNA expression was evaluated 1 hour after CRH administration to provide a comparable time-point to that used for the LCMRglc analysis. Both significant overlap and discrepancies were noted, with several brain structures found to be activated after CRH administration using one of the two methods only. No c-fos mRNA signal was detectable in any brain region of stress-free animals that were killed within 45 seconds of the investigators' entry to the animal facility (data not shown). The specific effects of CRH on c-fos mRNA expression were distinguished from those of handling and infusion procedures by choosing vehicle-treated animals sacrificed 1 hour after infusion as controls. Fig. 2 illustrates the pattern of c-fos mRNA expression induced by CRH at single cell and regional levels, focusing on structures analyzed for CMRglc.
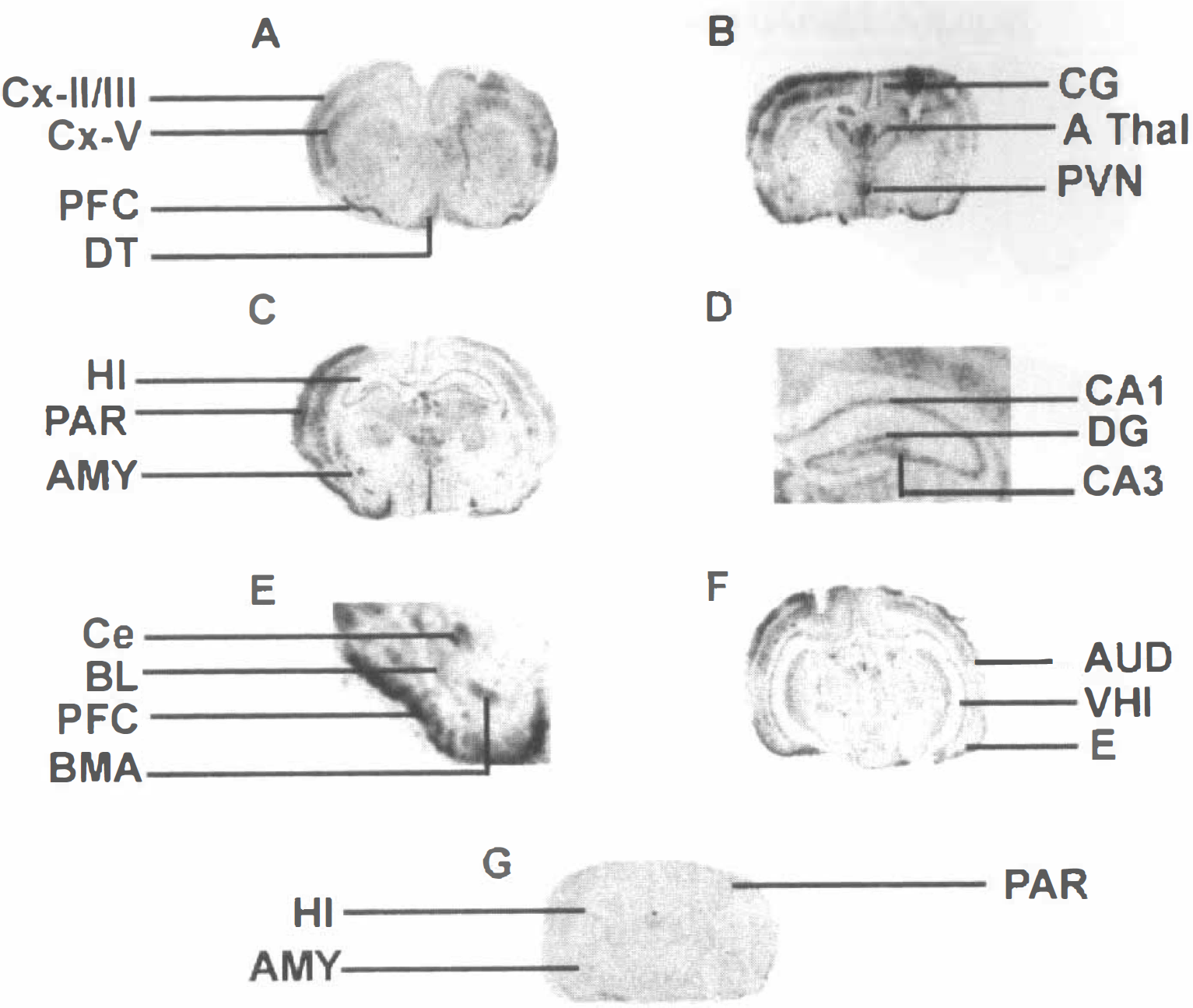
Expression of the immediate early gene c-fos mRNA after CRH administration. Coronal sections from immature rats subjected to in situ hybridization and semiquantitative analysis, as described in Materials and Methods.
Predominantly ipsilateral expression of c-fos mRNA was induced in most cortical regions (Table 2). C-fos expression was particularly prominent over the cingulate, piriform, parietal, and entorhinal cortices; whereas in frontal and sensorimotor cortices, induction of c-fos mRNA did not reach statistical significance as opposed to CMRglcs. The most striking increase of CRH-induced c-fos expression (P < 0.01) was observed in the ipsilateral piriform cortex.
Effects of CRH injection on the expression of c-fos mRNA in the brain of P10 rats
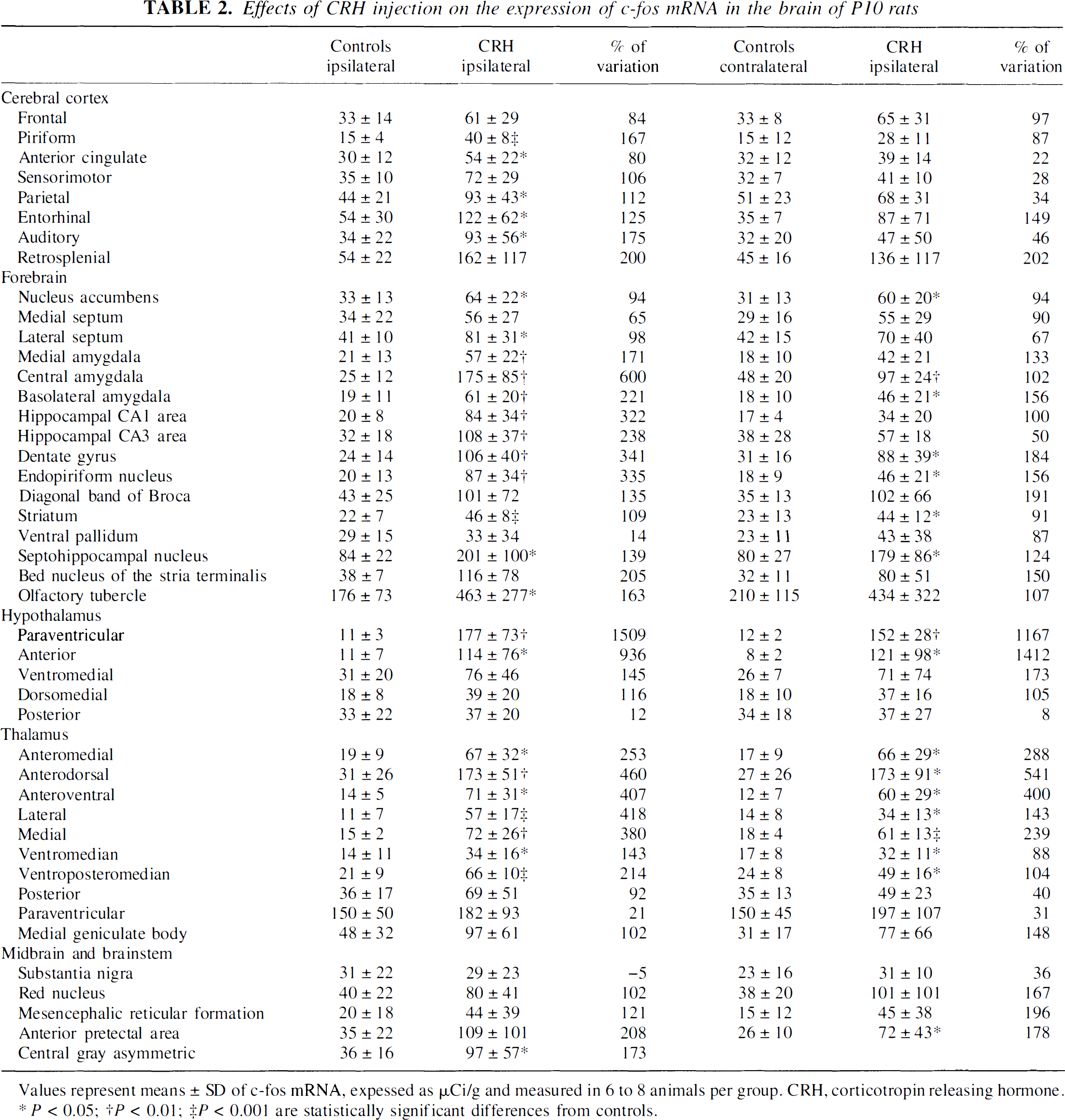
Values represent means ± SD of c-fos mRNA, expessed as μCi/g and measured in 6 to 8 animals per group. CRH, corticotropin releasing hormone.
P< 0.05;
P< 0.01;
P < 0.001 are statistically significant differences from controls.
In limbic regions, c-fos expression was enhanced bilaterally over the dentate gyrus granule cell layer (Table 2). In hippocampal CA3 and CA1 pyramidal cell layers, c-fos induction was selectively ipsilateral. In the amygdaloid complex, whereas robust induction of c-fos expression was found in the ACe and basolateral nuclei bilaterally, only ipsilateral medial nucleus was activated. In addition, robust c-fos expression was induced by CRH bilaterally in the endopiriform, septo-hippocampal, and accumbens nuclei; ipsilaterally in the lateral septum; and not at all in the medial septum.
In motor regions, c-fos increased significantly and bilaterally in the striatum. In contrast with metabolic activation, c-fos mRNA expression was not induced by CRH in substantia nigra or ventral pallidum. In thalamus, bilateral c-fos expression was significantly increased in most nuclei. In hypothalamus, in contrast to the lack of metabolic activation, significant c-fos expression was induced bilaterally in paraventricular nucleus and anterior hypothalamic area. Ipsilateral c-fos induction in the ventromedial hypothalamus, at levels close to statistical significance (P = 0.08), was also observed.
In mesencephalic structures, c-fos expression was significantly induced in the contralateral pretectal area, and in the midline central gray. In contrast to metabolic activation, CRH did not induce c-fos expression in the reticular formation or red nucleus.
Effects of corticotropin releasing hormone on activity and behavior
Administration of CRH resulted in a consistent, stereotyped spectrum of behaviors, as described in detail previously (Baram et al., 1992, 1997b). After a latency of 7.1 ± 1.1 minutes, rats developed repetitive and continuous oral automatisms, consisting of chewing and licking motions. These were associated with excessive grooming and were then interspersed with wet-dog shakes and semi-rhythmic forepaw clonus. These behaviors, aside from obvious involvement of motor circuits, have been shown to correlate with electrographic discharges in limbic circuits (Baram et al., 1992, 1997b).
DISCUSSION
The principal findings of the current study were as follows: (1) CRH administration resulted in activation of widespread yet selective brain regions comprising cortical, limbic, and motor circuits; (2) the [14C]2DG quantitative autoradiographic method and the expression of the immediate early gene c-fos yielded only partially overlapping, complementary information about the full extent of neuronal circuits activated by CRH; (3) CNS structures activated by CRH included regions rich in CRF and those expressing primarily CRF2 receptors, suggesting that both receptors may be targets for the peptide and involved in mediating its physiologic actions.
Corticotropin releasing hormone activated neuronal structures involved in a number of discrete circuits. For example, cortical regions including the entorhinal, parietal, and olfactory cortices were activated using both c-fos and metabolic criteria. A limbic circuit consisting of the hippocampus, certain amygdala nuclei, the septal region and nucleus accumbens was activated by CRH, as was a motor circuit consisting of basal ganglia and ventral pallidum. Additional CRH-activated regions consisted of select dorsal thalamic nuclei involved in transmission of specific input to limbic, prefrontal, and temporal association areas.
The majority of structures activated by CRH using c-fos induction (Marrosu et al., 1996), glucose utilization, or both criteria, have been shown to express mRNA for one or both CRH receptor types (Potter et al., 1994; Chalmers et al., 1995; Avishai-Eliner et al., 1996; Eghbal-Ahmadi et al., 1998). For example, in the majority of cortical regions, CRF1-mRNA is most abundant in layers II and V-VI (Potter et al., 1994; Chalmers et al., 1995; Avishai-Eliner et al., 1996). Whereas cortical c-fos expression closely overlapped the laminar distribution of CRF1 (Fig. 2), the [14C]2DG method did not permit differentiation of a cortical laminar pattern. However, the quantitative 2DG method demonstrated a correlation between areas with the most prominent metabolic activation (>300%) induced by CRH administration and the densest expression of CRF1 receptors—that is, in frontal, cingulate, and parietal cortices (De Souza et al., 1984; Chalmers et al., 1995; Chen et al., 2000). This correlation was also supported by the pattern of c-fos expression. In addition, while CRF2-receptor distribution has generally been confined to subcortical regions, CRF2-mRNA has been reported in piriform and frontal cortices (Chalmers et al., 1995; Eghbal-Ahmadi et al., 1998) so that CRF2 may participate in CRH-induced cellular activation in these two cortical regions.
In thalamus, widespread CRH-induced neuronal activation was demonstrated using both methods, with a maximal metabolic increase (200%) in the mediodorsal nucleus. Activation patterns correlated well with the distribution of CRF1-immunoreactive neurons, particularly prominent in the rodent mediodorsal nucleus (Chen et al., 2000). Conversely, CRF2 expression in thalamus of the immature rat was essentially confined to the ventroposterior medial and lateral nuclei (Eghbal-Ahmadi et al., 1998), suggesting that in thalamus, as in cortical areas, the neuronal actions of CRH are likely to primarily involve activation of CRF1.
In addition, the current study suggests that, at least in certain CRH-modulated circuits, the peptide's action might require activation of CRF1 and CRF2. For example, the hippocampal formation, highly activated by CRH using glucose metabolism and c-fos expression, has been shown to express CRF1 (Avishai-Eliner et al., 1996) and CRF2 (Eghbal-Ahmadi et al., 1998). Thus, in the dentate gyrus (bilaterally activated using both methods), and in CA3 and CA1 fields (bilaterally activated using 2DG, but expressing c-fos unilaterally), both receptors may participate in transducing the cellular actions of the neuropeptide. Other components of the limbic circuit, for example, the lateral septum and medial nucleus of the amygdala, are rich in CRF2-mRNA and virtually devoid of CRF1-mRNA. Because both of these regions expressed c-fos on the side of CRH infusion, and metabolic activity of lateral septum was enhanced bilaterally, it is likely that their activation requires binding of CRH to the CRF2 receptor. This finding also supports the notion that CRH may function as an endogenous ligand for this latter receptor. Conversely, in the basolateral amygdala nuclei, expressing almost exclusively CRF1 receptors, the striking CRH-induced activation may require binding to this receptor type, found on target neurons for the actions of CRH.
In the current study, robust activation was noted also in structures relatively devoid of either CRF1- or CRF2-mRNAs, yet strongly implicated in functions attributable to CRH. An eloquent example involves the central nucleus of the amygdala (ACe), where CRH is released during stress (Merali et al., 1998), and direct CRH administration leads to behaviors found during stress (Swiergiel et al., 1993). Pronounced activation of ACe was evident in this study, whereas CRF1-mRNA expression in this nucleus has been found to be modest (Potter et al., 1994; Chalmers et al., 1995; Avishai-Eliner et al., 1996). However, recent immunocytochemical analyses (Chen et al., 2000) indicate intense and abundant CRF1-like immunoreactivity in ACe, suggesting that CRF1 in ACe may be transported from cell bodies residing in other nuclei (for example, lateral amygdala;Pitkanen et al., 1997). Thus, the current study—demonstrating activation of ACe by CRH—implicates ACe-CRF1 receptors in the mechanisms by which endogenous CRH may exert physiologic and behavioral responses involving this nucleus.
Indeed, several regions shown in this article to be activated by CRH receive CRH-containing projections, such as those from the amygdaloid ACe to the brainstem and hypothalamus (Swanson et al., 1983; Sakanaka et al., 1986; Pitkanen et al., 1997). In the current study, CRH induced c-fos expression in hypothalamic nuclei—specifically the paraventricular nucleus, and anterior and ventromedial nuclei—without increasing glucose utilization. The receptors underlying these effects have not been fully resolved. Most studies have not revealed expression of CRF1 in the paraventricular nucleus of the stress-free rat (Potter et al., 1994; Chalmers et al., 1995), whereas recent immunocytochemistry has shown presence of CRF1 in rodent paraventricular nucleus (Chen et al., 2000). CRF2 expression has been demonstrated in the paraventricular nucleus of the adult, but not the 10-day-old rat (Eghbal-Ahmadi et al., 1998). In the ventromedial nucleus, abundant CRF2 has been shown in immature and adult rat, whereas CRF1 is not largely expressed (Chalmers et al., 1995), indicating that CRH may well activate hypothalamic ventromedial neurons through binding to CRF2. In addition, it appears that, at least in hypothalamic neurons, an increase in CMRglc is not a prerequisite for immediate early gene induction. A mismatch between c-fos induction and metabolic activation, similar to the one reported in this article, has been shown previously after water deprivation. In that paradigm, expression of c-fos protein was markedly increased in hypothalamic magnocellular nuclei without metabolic activation, despite known increases in neuronal firing rates (Sagar et al., 1988).
A mismatch of c-fos expression and metabolic activation was evident also in an additional important projection area of CRH-containing neurons from Ace, the bed nucleus of stria terminalis (BNST). In this region, rich in CRF1-immunoreactive neurons (Chen et al., 2000), and in the dorsomedial hypothalamic and septo-hippocampal nuclei, small cell groups expressing c-fos were readily discernible. However, because they comprised a minor proportion of the cells of the respective structure, the activation of these neuronal populations did not enhance overall regional metabolism significantly.
In the motor circuit, CRH-induced activation was evident in the basal ganglia and ventral pallidum. Earlier binding studies (De Souza et al., 1984), not distinguishing between CRF1 and CRF2, have demonstrated CRH receptors in globus pallidus and striatum. Recently, CRF1-like immunoreactivity has been localized to rodent basal ganglia including striatum, globus pallidus, and ventral pallidum (Chen et al., 2000). The presence of CRF2-immunoreactivity in these regions has not been determined.
Within midbrain, whereas metabolism increased in the central gray, substantia nigra, red nucleus, inferior and superior colliculus, pretectal area, and mesencephalic reticular formation, c-fos induction was evident solely in the central gray and pretectal area. Thus, in midbrain, as in cortical areas, the 2DG method seems a more sensitive measure of neuronal activation than c-fos expression. This is not surprising because c-fos expression in response to CRH requires not only the presence of the cellular machinery to activate transcription of this immediate early gene, but also the presence of a mature signal transduction cascade downstream of the CRH receptor that is capable of inducing c-fos. Indeed the immaturity of the cyclic adenosine monophosphate second messengers' system implicated in c-fos induction in the immature rat has been documented. In addition, metabolic activation may propagate more readily through relay neurons than signals required for c-fos expression. This phenomenon has been demonstrated both after cutaneous stimulation and focal onset seizures (Gass et al., 1997). In both settings, activation of c-fos was restricted to the site of stimulation and second-order relay nuclei, whereas enhanced metabolism involved third-or fourth-order relay neurons.
It should be noted that the studies described in this article were conducted on immature 10-to 12-day-old rats, in which CRH has been shown to result in intense excitation of limbic neurons leading to electrographic and limbic seizures (Baram et al., 1992, 1997b; Baram and Hatalski, 1998). The mechanisms for this enhanced potency of CRH are not fully understood, but may derive at least in part from the increased expression of CRF1, the receptor mediating CRH-induced neuronal activation, in hippocampal and amygdala neurons during this developmental phase. Emerging evidence supports the notion that endogenous CRH is also more abundant in amygdala and hippocampus early in postnatal life (Yan et al., 1998; and unpublished observations), supporting additional roles for the peptide in neuronal and circuit plasticity during development. These age-specific actions of CRH probably underlie some of the differences between an earlier study (Sharkey et al., 1989) and the current study. Using adult animals, Sharkey et al. (1989) found CRH-induced increases in cerebral metabolism primarily in neuronal circuits involved in coordination of the stress response, whereas the current study implicated a more wide-ranging involvement of limbic and cortical circuits.
The use of immature rats also determined the magnitude of CRH-induced metabolic increases recorded in the current study, which ranged between 150% and 400% (compared with generally <100% increases found by Sharkey et al., 1989). Activation in this range has been previously documented in immature rats of the same age after administration of other neuronal excitants, that is, pentylenetetrazol (Pereira de Vasconcelos et al., 1992) and lithium-pilocarpine (Fernandes et al., 1999). In fact, the latter regimen led to metabolic increases of over 600% above control values in cerebral cortex and limbic forebrain regions. It should be emphasized that neuronal death has not been observed in any of these three paradigms. This ability to tolerate massive enhancement of metabolic activity without neuronal death seems unique to the immature brain.
In summary, the extent of neuronal activation by CRH was demonstrated using the independent methods of c-fos expression and glucose utilization. While excellent general congruence of activation was observed in the circuits noted above, discrepancies were observed, likely related to enhanced sensitivity and superior quantification provided by metabolic activation, contrasted with the ability to detect small cell groups expressing c-fos without alteration of regional metabolism. Corticotropin releasing hormone-activated structures generally express mRNA for one or both CRH receptors, or receive these receptors through axonal transport. As these structures participate in important motor and limbic circuits, their activation by CRH provides evidence for a modulatory role of CRH on functions subserved by these circuits.
In conclusion, glucose metabolism complemented by induction of immediate-early gene expression yielded quantitative and detailed analysis of the neuroanatomical matrix of CRH-induced neuronal activation. Overlapping regions activated using both methods delineated discrete cortical, limbic and motor pathways. Importantly, cell groups activated by CRH included those possessing either or both members of the CRH receptor family, suggesting that both receptors may mediate the effects of the endogenous ligand. These findings indicate that stress-related release of CRH may contribute to a spectrum of important modulations of CNS function.
Footnotes
Abbreviations used
Acknowledgements
The authors thank M. Hinojosa for editorial support and W. Cariaga for excellent technical assistance.