
Abstract
Although neurons and glia inevitably undergo degeneration in the core of ischemic lesions, many cells, particularly immune cells, infiltrate the core and survive in it. Such infiltrating cells may play certain roles in the regeneration and repair of damaged brain tissues. In this study, we characterized macrophage-like cells that accumulated in the ischemic core of a rat brain whose right middle cerebral artery was transiently occluded for 90 mins. Many of the accumulated macrophage-like cells expressed Iba1, a marker of macrophages/microglia, as well as NG2 chondroitin sulfate proteoglycan (NG2), which has been recognized as a marker of oligodendrocyte progenitor cells. Such macrophage-like cells were termed BINCs (brain Iba1+/NG2+ cells) to distinguish them from NG2/Iba1+ or NG2+/Iba1− cells that were also present in the perilesion and the contralateral hemisphere. Electron microscopy showed the localization of NG2 along the plasma membrane of cells that had many phagosomes and irregular-shaped or reniform heterochromatin-rich nuclei, which are characteristics of monocytes/macrophages. Brain Iba1+/NG2+ cells were highly proliferative and their number peaked at 7 days post-reperfusion. An immunoblot analysis of NG2 revealed the presence of two NG2s: one expressed by BINCs with a molecular weight of 300 kDa, and the other found in the contralateral hemisphere with a molecular weight of 290 kDa. Taken the various functions of NG2, BINCs may be involved in not only phagocytosis of degenerated cells but also the healing and regeneration of lesion cores.
Introduction
NG2 chondroitin sulfate proteoglycan (NG2) has been considered a marker of progenitor cells in the brain. In fact, the term ‘NG2 glia’ almost corresponds to oligodendrocyte progenitors (OPCs) and many NG2 glia in the normal brain express platelet-derived growth factor receptor α (PDGFRα), another marker of OPCs (Nishiyama et al, 2002; Stallcup, 2002). The term ‘polydendrocytes’ has been proposed for NG2 glia, on the basis of their morphology showing elaborate multiple-branched processes and a profound relationship in lineage with oligodendrocytes (Nishiyama et al, 2002). NG2 glia can generate not only oligodendrocytes but also stellate or type 2 astrocytes in vitro. Therefore, the term ‘oligodendrocyte-type 2 astrocyte progenitors’ has been used for NG2 glia (Stallcup, 2002). Furthermore, NG2 glia or oligodendrocyte-type 2 astrocyte progenitors have been shown to give rise to neurons (Belachew et al, 2003; Kondo and Raff, 2000). Thus, NG2 glia are thought to be in an undifferentiated state in a normal brain. Therefore, NG2 has attracted attention as a candidate molecular marker of multipotent stem cells in the brain.
In addition to NG2 glia, many macrophage-like cells that accumulate in central nervous system (CNS) lesions have been shown to be NG2+ (Bu et al, 2001; Hampton et al, 2004; Jones et al, 2002; McTigue et al, 2001; Yokoyama et al, 2006). Such NG2+ cells have been classified into macrophage/monocyte lineage by positive immunohistochemical staining with an antibody to CD11b or Iba1 (Imai et al, 1996), both of which are microglial/macrophage markers. These cells are highly proliferative and accumulate in the core of CNS lesions (Hampton et al, 2004; Zai and Wrathall, 2005). Although there is little knowledge of their nature and roles, NG2+ macrophage-like cells have been recognized as blood—borne cells distinct from neuroectodermal NG2 glia (Bu et al, 2001). However, NG2+ macrophage-like cells isolated from stab wounds or NG2+ primary-cultured microglia give rise to cells with neuroectodermal phenotypes as NG2 glia do (Yokoyama et al, 2006). Thus, the accumulation of NG2+ macrophage-like cells has been reported in several types of lesion of the CNS, whereas the presence of such NG2+ cells in ischemic lesions of the brain has not been reported.
In this study, we aim to determine whether NG2+ macrophage-like cells are present in ischemic lesions using a rat model whose middle cerebral artery (MCA) has been transiently occluded. We find that NG2+ macrophage-like cells markedly accumulate during a subacute phase of ischemic events. Because electron microscopy reveals that NG2+ cells bear many phagosomes within their cytoplasm, they may be involved at least in the removal of degenerated tissue. The NG2 expressed by the macrophage-like cells has a distinct molecular weight from that of NG2 expressed in the normal mature brain. Finally, we used the term BINCs (brain Iba1+/NG2+ cells) for NG2+ macrophage-like cells that accumulate in the ischemic core to distinguish them from NG2 glia, reactive resident microglia, and NG2− monocytes.
Materials and methods
Animals and Surgical Procedures
The experiments were conducted in accordance with the Guide for Animal Experimentation of Ehime University School of Medicine. Adult male Wistar rats (8 to 10 weeks old; body weight, 230 to 280 g) were used for all experiments. They were anesthetized with a gas mixture of halothane (1.0% to 1.5%), oxygen (30%), and nitrogen (70%) during operation. A temperature probe was inserted into the rectum of the rats, and a heating lamp was used to maintain rectal temperature at 37.0°C to 37.5°C. The right MCA was occluded by an intraluminal filament technique, as described previously (Hashimoto et al, 1999). To obstruct the origin of the right MCA, a 4-0 nylon monofilament suture was inserted into the internal carotid artery through the right external carotid artery. After the intraluminal filament was placed in the correct position, the neck incision was sutured. The rats were evaluated for neurologic deficits 1 h after operation by the method defined by Bederson et al (1986) using the following scale: grade 3, consistently circling toward the left (paretic) side; grade 2, consistently showing a reduced resistance to lateral push toward the left side without circling movement; grade 1, showing consistent forelimb flexion but no other abnormalities; grade 0, showing no neurological deficits. Only the rats classified as grade 2 or 3 were used for further study.
The rats were reanesthetized and their intraluminal suture was carefully removed 90 mins after MCA occlusion. They were then reared for an additional 1 to 14 days with free access to food and water. Rats in the sham control group were treated in the same way but without MCA occlusion.
In Vivo Immunohistochemistry
The rats (n = 5) were killed at 7 days post-reperfusion under ether anesthesia and transcardially perfused with a fixative (4% paraformaldehyde in phosphate-buffered saline (PBS) containing 2 mmol/L MgCl2) for 10 mins at a flow rate of 80 mL/min. The dissected brains were immersed in 15% sucrose in PBS at 4°C overnight, rapidly frozen in dry ice powder and sliced at the caudoputamen level into 4-μm thick coronal sections using a cryostat.
Two immunohistochemical techniques were employed: the streptoavidin—peroxidase method for double staining, and the indirect immunofluorescence method for triple staining. Unless otherwise stated, incubation was performed at room temperature.
Streptoavidin—Peroxidase Method: After the brain sections were rinsed in Tris-buffered saline containing 0.1% Tween 20 (TBSt) for 20 mins, endogenous peroxidase activity was blocked by incubating the sections in methanol containing 0.6% H2O2 for 30 mins. Then, the sections were incubated in TBSt containing 1% bovine serum albumin (BSA; Sigma Chemical Co., St Louis, MO) for 20 mins, followed by incubation with an anti-Ki67 rabbit polyclonal antibody (Table 1) at 4°C overnight. The sections were further incubated with a biotinylated goat anti-rabbit IgG antibody (Labvision, Fremont, CA) for 60 mins, and then with streptavidin—peroxidase (Labvision) for 60 mins. Immunoreaction was visualized with 3-amino-9-ethylcarbazole as chromogen (Labvision). For double staining to detect Iba1 in the same sections, the sections were incubated in 0.1mol/L glycine-HCl buffer (pH 2.5) for 1 h to remove the antibodies bound to the sections. After incubation in TBSt containing BSA, the sections were incubated with a polyclonal antibody to Iba1. Immunoreaction for Iba1 was visualized by peroxidase reaction with True Blue (KPL, Gaithersburg, MD) as a substrate of peroxidase.
Primary antibodies used in this study
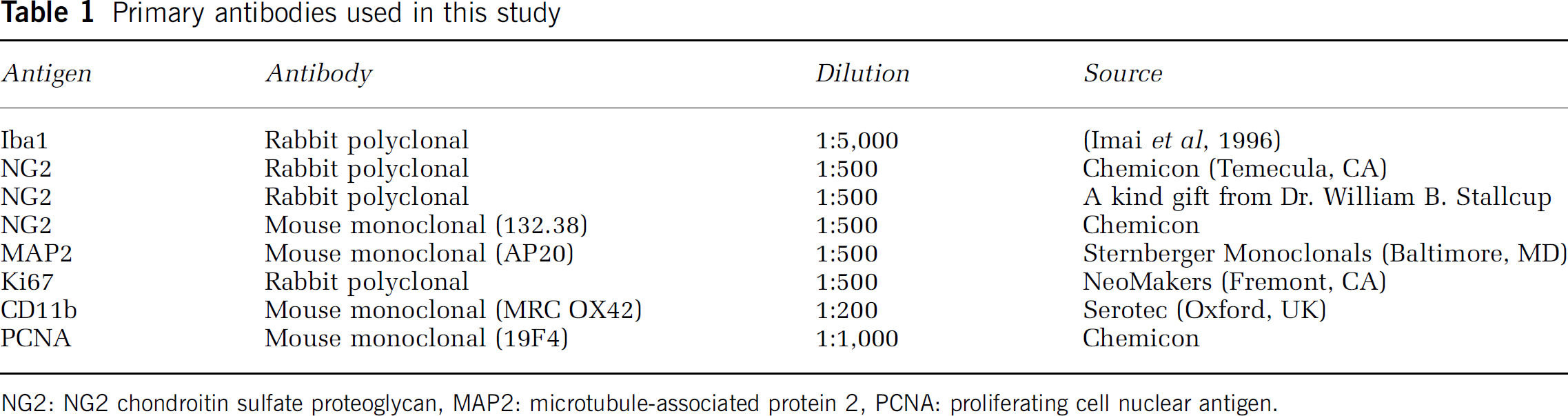
NG2: NG2 chondroitin sulfate proteoglycan, MAP2: microtubule-associated protein 2, PCNA: proliferating cell nuclear antigen.
Indirect Immunofluorescence Method: After the brain sections were sequentially rinsed in TBSt and incubated in TBSt containing BSA, they were incubated with the primary antibodies listed in Table 1. Immunoreaction was visualized with fluorescein isothiocyanate- and Cy3-labeled secondary antibodies. Hoechst 33258 was used for nuclear staining.
The immunostained specimens were observed with a BX-52 Olympus (Tokyo, Japan) conventional microscope equipped with a CCD camera and differential interference contrast optics.
Immunoelectron Microscopy
Immunoelectron microscopic observation was performed as described previously (Matsumoto et al, 2007a). In brief, the rats subjected to transient MCA occlusion were fixed at 7 days post-reperfusion by a 10-min transcardial perfusion of a fixative containing 4% paraformaldehyde and 0.2% glutaraldehyde (Wako, Osaka, Japan) in PBS. After blocking endogenous peroxidase activity, the sliced brain was incubated in TBS containing 0.02% Tween 20 and 0.1% BSA for 20 mins, followed by incubation with polyclonal antibodies to Iba1 or NG2, or a monoclonal antibody to NG2 diluted in TBS containing 0.1% BSA at 4°C overnight. The slices were incubated with a biotinylated goat anti-mouse IgG antibody (Chemicon) for 60 mins, and then with streptavidin—peroxidase (Labvision) for 60 mins. Immunoreaction was visualized with diaminobenzidine (Nichirei, Tokyo, Japan) as a chromogen. After postfixation, dehydration, and embedding in Epon, ultrathin sections were cut, stained with lead citrate and uranyl acetate solutions, and observed using an electron microscope operated at 100 kV (JEM 1230; JEOL, Tokyo, Japan).
Counting Cells in Ischemic Core
Brain sections fixed at 1, 3, 7, or 14 days post-reperfusion were double-immunostained with anti-Iba1 and monoclonal anti-NG2 antibodies in combination with nuclear staining (n = 3, each time point). Micrographs were obtained with a CCD camera using a × 20 objective lens. Four or five × 20 optic fields (0.14 mm3/field) of the ischemic core in the temporal cortex (somatosensory and visceral areas; denoted as C in Figure 1) were printed. The total number of cells in the five fields was obtained by counting Hoechst 33258-stained nuclei. Iba1+/NG2+, Iba1−/NG2+, and Iba1+/NG2− cells were also counted.
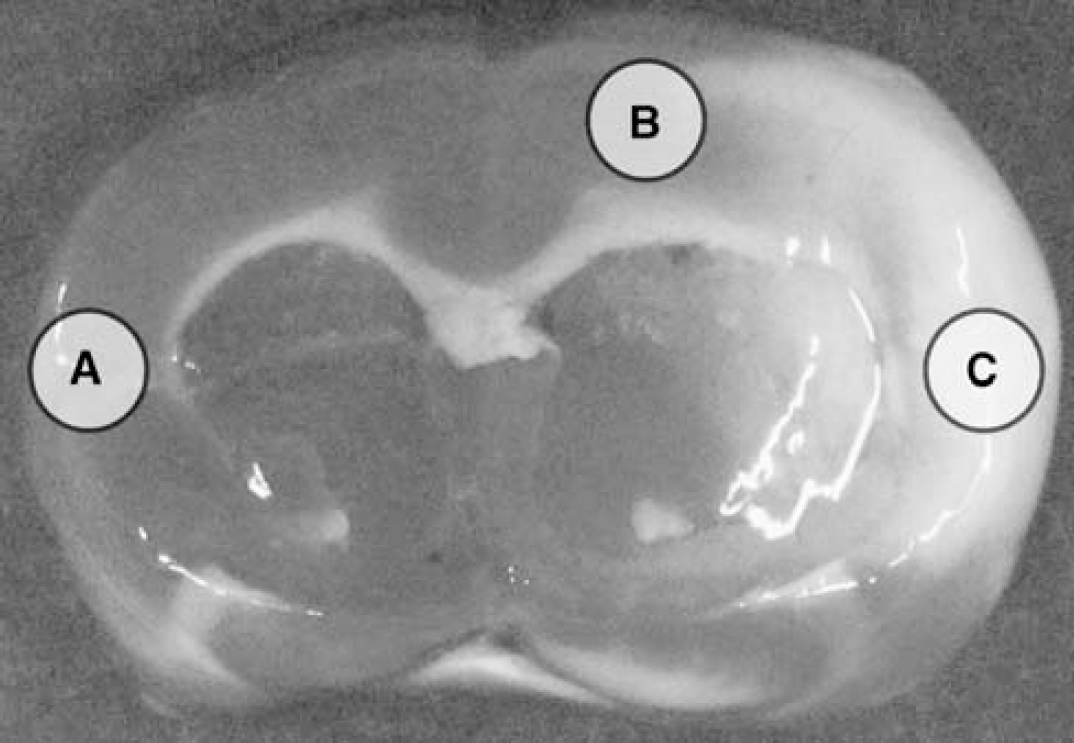
Definitions of (
Immunoblotting of Brain Tissue
The rats deeply anesthetized with ethyl ether were transcardially perfused with chilled PBS for 3 mins at a flow rate of 80 mL/min at 7 days post-reperfusion. The brains were dissected, and coronal sections of 3 mm thickness were prepared at the caudoputamen level. Some sections obtained at 3 days post-reperfusion were stained with 2% 2,3,5-triphenyltetrazolium hydrochloride (Wako) for 3 mins at room temperature. On the basis of the results of the 2,3,5-triphenyltetrazolium hydrochloride staining, the contralateral cortex, the remote region of the ipsilateral cortex and the ischemic core of the cerebral cortex were defined (Figure 1). The tissue samples from these three regions at 1, 3, 5, 7, and 14 days post-reperfusion were immediately homogenized with Polytron in 10 volumes of PBS, and then mixed with Laemmli's sample solution. The lysates were electrophoresed, transferred to nitrocellulose membranes, and immunoblotted with antibodies to β-actin, Iba1, NG2, proliferating cell nuclear antigen (PCNA), and MAP2 (Table 1). Immunoreaction was visualized using nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate, as described previously (Tanaka et al, 1998).
Immunoreactive bands were analyzed by densitometry using NIH image software. The densitometry data were classified in terms of β-actin density. Data from three independent experiments were subjected to statistical analysis using analysis of variance with Schéffe's post hoc test (Tanaka et al, 1999).
Isolation of Brain Iba1+/NG2+ Cells Accumulated in Ischemic Core
To characterize NG2 expressed by BINCs, BINCs were isolated from the ischemic hemisphere and briefly cultured as follows. At 7 days post-reperfusion, the deeply anesthetized rats were transcardially perfused with chilled PBS containing an antibiotic-antimicotic mixture (Invitrogen, Carlsbad, CA) for 3 mins at a flow rate of 80 mL/min. The brains were dissected, and the ischemic hemisphere was minced with scissors and triturated using a Pasteur pipette in 10 mL of PBS containing 0.25% ethylene-diamine tetraacetic acid (EDTA; Sigma) for 5 mins. The mixture of minced tissues was filtered through a nylon mesh of 160 μm pore size, and then the filtrate was centrifuged at l,000r.p.m. for 5 mins at 4°C. The pellet was resuspended in 6 mL of Dulbecco's modified Eagle's medium (Sigma) containing 3% fetal bovine serum (Wako). Isolated cells were seeded on polystyrene dishes for suspension culture (Sumitomo, Akita, Japan) and incubated for 30 mins in a CO2 incubator. Macrophage/microglia-lineage cells but not neuroectodermal cells preferentially attach to the dishes. The dishes were rinsed with warm Dulbecco's modified Eagle's medium containing 3% fetal bovine serum to remove unattached cells that are not of the macrophage/microglial lineage (Tanaka et al, 1998). For immunoblot analysis, the cells were lysed with Laemmli's sample solution containing 3% sodium dodecyl sulfate. The lysate was electrophoresed, transferred onto nitrocellulose membranes, blotted with antibodies to NG2, and visualized with 5-bromo-4-chloro-3-indolyl phosphate and nitro blue tetrazolium. For immunocytochemistry, purified Iba1+ cells were scraped off with a rubber scraper, and reseeded onto poly-
Chondroitinase ABC Treatment
Brain tissues from the contralateral, remote, and core regions were homogenized with Polytron in 10 volumes of 40 mM Tris—HCl (pH 8.0) containing 40 mol/L sodium acetate at 7 days post-reperfusion (n = 2). Then, they were incubated with chondroitinase ABC (0.05 U/mL; Sigma) at 37°C for 1 h (Dou and Levine, 1994). After the incubation, the tissues were mixed with Laemmli's sample solution, boiled, electrophoresed, and immunoblotted with monoclonal and polyclonal anti-NG2 antibodies.
N-glycanase Treatment
Purified BINCs isolated from the ischemic hemisphere at 7 days post-reperfusion seeded on polystyrene dishes for suspension culture were scraped off with a rubber scraper in 1 mL of PBS containing EDTA (Sigma) and 1 μL of protease inhibitor mixture (Wako). For immunoblotting study, part of the cell suspension was lysed with Laemmli's sample solution as a control. Deglycosylation was performed using a Glyco N-Glycanase kit (PROzyme, San Leandro, CA). After centrifugation at 15,000 r.p.m. for 20 mins at 4°C, the pellet of the suspension was resuspended in 100 μL of the buffer provided by the manufacturer with the protease inhibitor mixture, and boiled for 5 mins. The suspended pellet was mixed with a detergent solution provided by the manufacturer, and centrifuged at 15,000 r.p.m. for 20 mins at 4°C. The supernatant was divided into two parts: one was mixed with N-Glycanase in the supplied buffer, and the other with the buffer alone. The two solutions were incubated for 18 h at 37°C. The solutions were mixed with Laemmli's sample solution, electrophoresed, transferred onto nitrocellulose membranes, blotted with monoclonal and polyclonal antibodies to NG2, and visualized with 5-bromo-4-chloro-3-indolyl phosphate and nitro blue tetrazolium.
Reverse Transcription-Polymerase Chain Reaction
Total RNA was extracted from tissue homogenates of the ischemic core at 7 days post-reperfusion (n = 2) and the remote and contralateral regions, and reverse-transcribed to obtain single-strand cDNA, as described elsewhere (Mori et al, 2002). For kinetic analysis, total RNA was also extracted from homogenates of ischemic tissues at 1, 3, 5, 7, and 14 days post-reperfusion (n = 2) and the control rat tissue. The sequences of the primers used are given in Table 2. Polymerase chain reaction was conducted using Taq polymerase (TaKaRa EX Taq, TaKaRa) to amplify cDNAs using the primers listed in Table 2 except primers B, F, and G for the NG2 core protein. Polymerase chain reaction amplification was performed as follows: cDNA products of the reverse transcription (RT) were denatured for 3 mins at 94°C. The amplification involved denaturation at 94°C for 1 min, annealing at 60°C for 1 min, and extension at 74°C 1 min. To obtain large cDNAs using primers B, F, and G for the NG2 core protein, polymerase chain reaction (PCR) amplification was performed as follows: 94°C for 3 mins, followed by 35 cycles of 94°C for 1 min, 60°C for 1 min, and 72°C for 3 mins using a mixture of Taq and Tgo polymerase (Expand High-Fidelity PCR system, Roche, Mannheim, Germany).
Oligonucleotide primers for RT-PCR
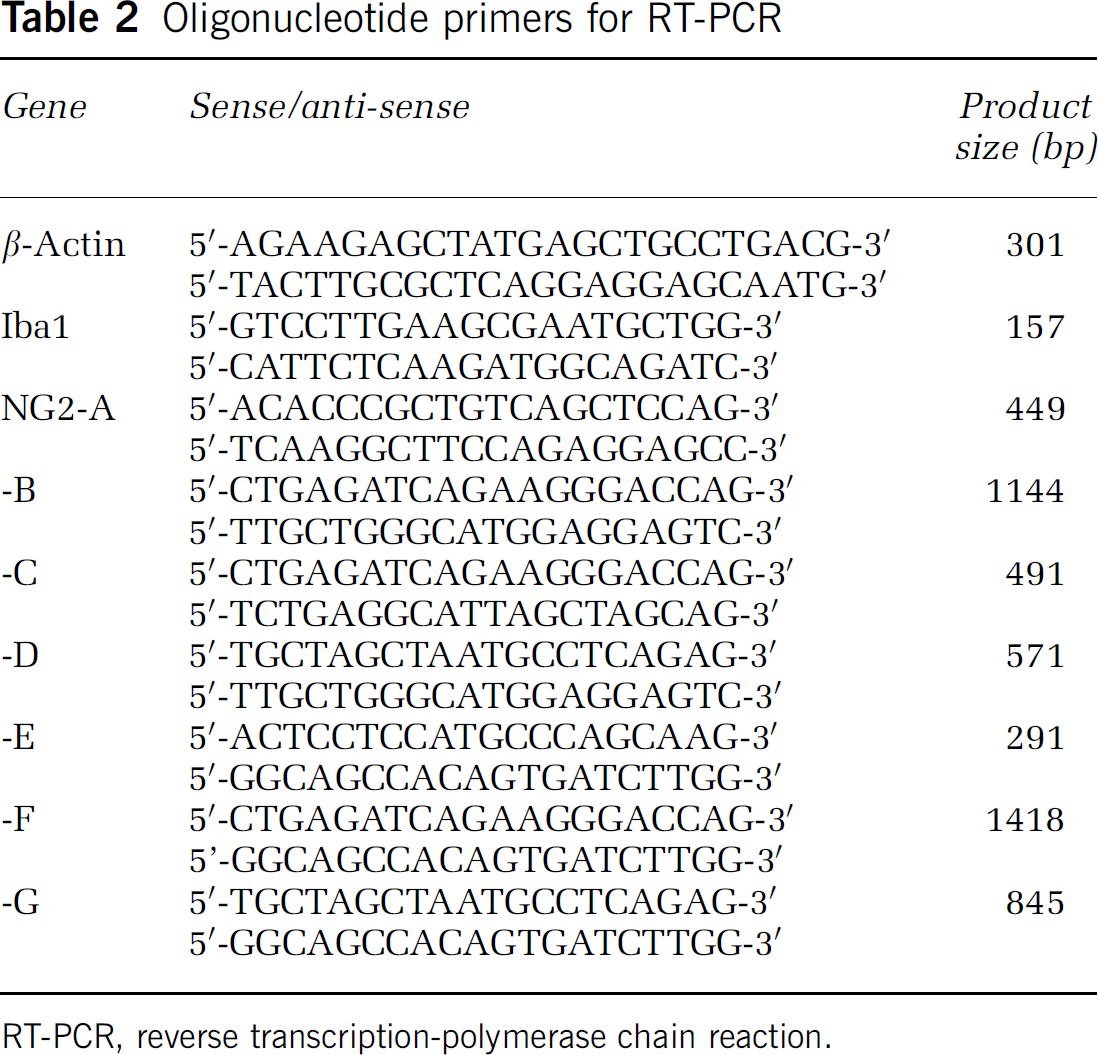
RT-PCR, reverse transcription-polymerase chain reaction.
Results
Immunohistochemical Localization of Macrophage Markers and NG2 in Ischemic Lesion
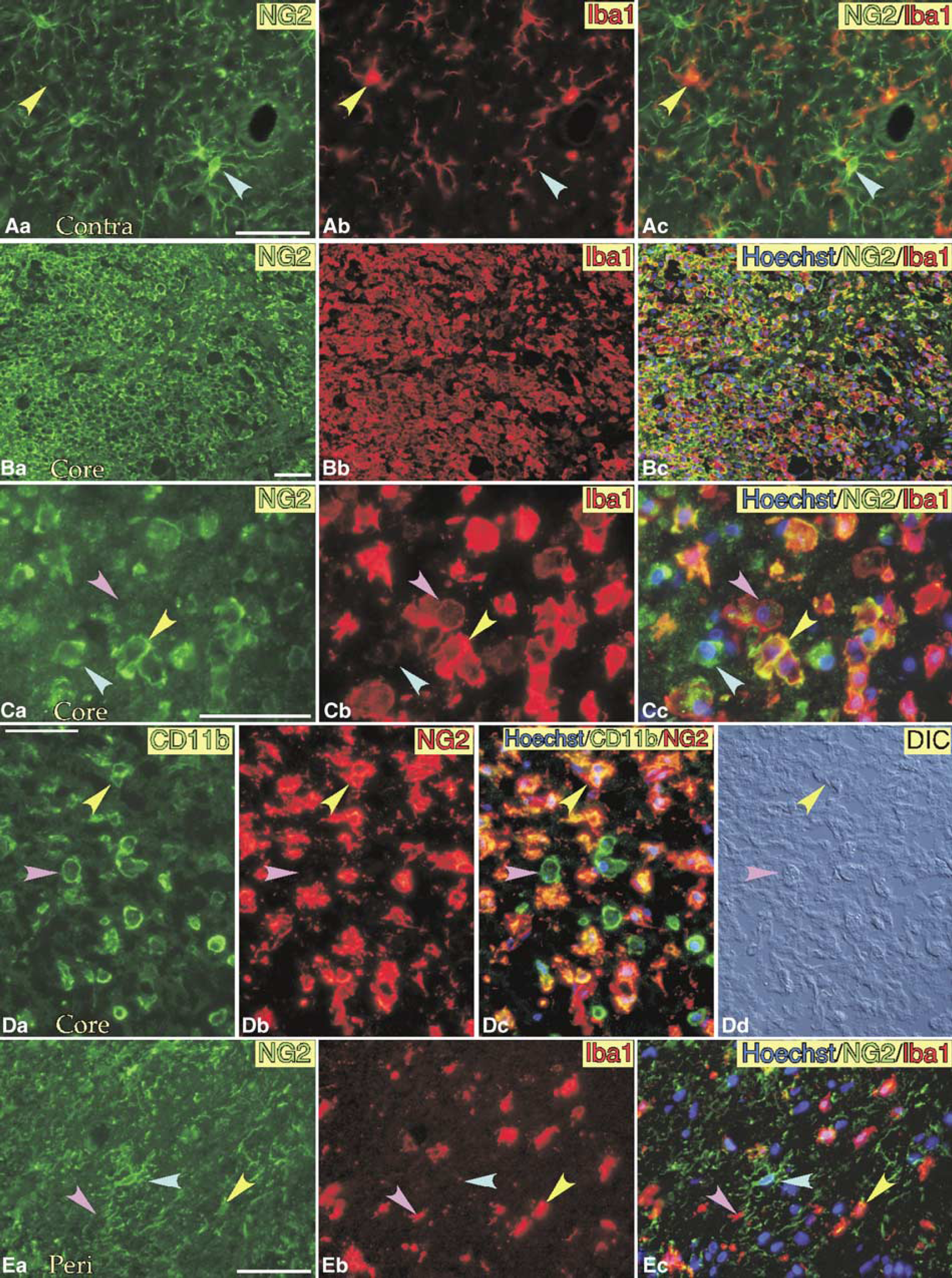
We used antibodies to Iba1 and CD11b as markers of microglia or macrophage-lineage cells. To localize NG2, both polyclonal and monoclonal antibodies to NG2 were used. Ramified NG2 glia were diffusely distributed in the normal brain (Figure 2A, blue arrowheads). They were distinct from Iba1+/NG2−microglia indicated by yellow arrowheads, and located close to each other, as described elsewhere (Nishiyama et al, 1997). In the ischemic core in the cerebral cortex of the right hemisphere at 7 days post-reperfusion (see Figure 1), many Iba1+ amoeboid cells were observed, most of which were NG2+ (Figure 2B). Closer observation revealed the presence of three types of cell depending on the expressions of Iba1 and NG2 in the ischemic core: Iba1+/NG2+ cells (yellow arrowheads in Figure 2C), Iba1+/NG2− cells (pink), and Iba1−/NG2+ cells (blue). Because the three types of cell were all amoeboid, they were indistinguishable from each other unless immunostained. From the data using a combination of nuclear staining with Hoechst 33258 for determination of total cell number in a optic field of × 20 lens and double-immunofluorescence staining with antibodies to Iba1 and NG2, nearly 80% of total cells in the ischemic core in the temporal cortex were Iba1+, and two-thirds of total Iba1+ cells exhibited NG2 immunoreactivity at 7 days post-reperfusion as summarized in Table 3. Therefore, about half of cells in the ischemic core were Iba1+/NG2+, which were termed BINCs. Iba1−/NG2+ cells formed nearly 5% of the total number of cells in the ischemic core at 7 days post-reperfusion, which may be residual NG2 glia or OPCs (Ohta et al, 2003).
The proportion of Iba1+ or NG2+ cells in ischemic core

Localization of Iba1 and NG2 observed by immunoelectron microscopy. The ischemic brain was fixed at 7 days post-reperfusion. (
The accumulation of BINCs in the ischemic core can also be identified by immunostaining with an antibody (OX42) to another microglial marker, CD11b (yellow arrowheads in Figure 2D). NG2−/CD11b+ cells indicated by pink arrowheads in Figure 2C were indistinguishable from BINCs based on observation with differential interference contrast optics (Figure 2Dd). In the peri-infarct region (Figure 2E), in addition to BINCs (yellow arrowheads) and amoeboid Iba1+/NG2− cells (pink), ramified Iba1−/NG2+ cells (blue) were present. Iba1+/NG2− cells with blunted processes may be reactive resident microglia.
Electron microscopy showed that many NG2+ cells in the ischemic core displayed monocytes/macrophage-like morphology (Figure 3). Macrophage-like cells with many phagosomes had an Iba1+ cytoplasm beneath the plasma membrane (Figure 3A). Anti-NG2 antibodies immunostained the plasma membrane of cells, whose nuclei were usually eccentric and often reniform, characteristic of monocyte/macrophage (Figure 3B to 3D). NG2+ cells often bore phagocytosed materials within their cytoplasm (Figure 3B and 3C). However, a significant number of NG2+ macrophage-like cells had no phagosomes (Figure 3D). In addition to such macrophage-like cells, a different type of NG2+ cell was found in small number as denoted by the asterisk in Figure 3D. Because this type of cell had no cytoplasmic phagosomes and had less chromatin in the nucleus, it may be an NG2 glial cell.
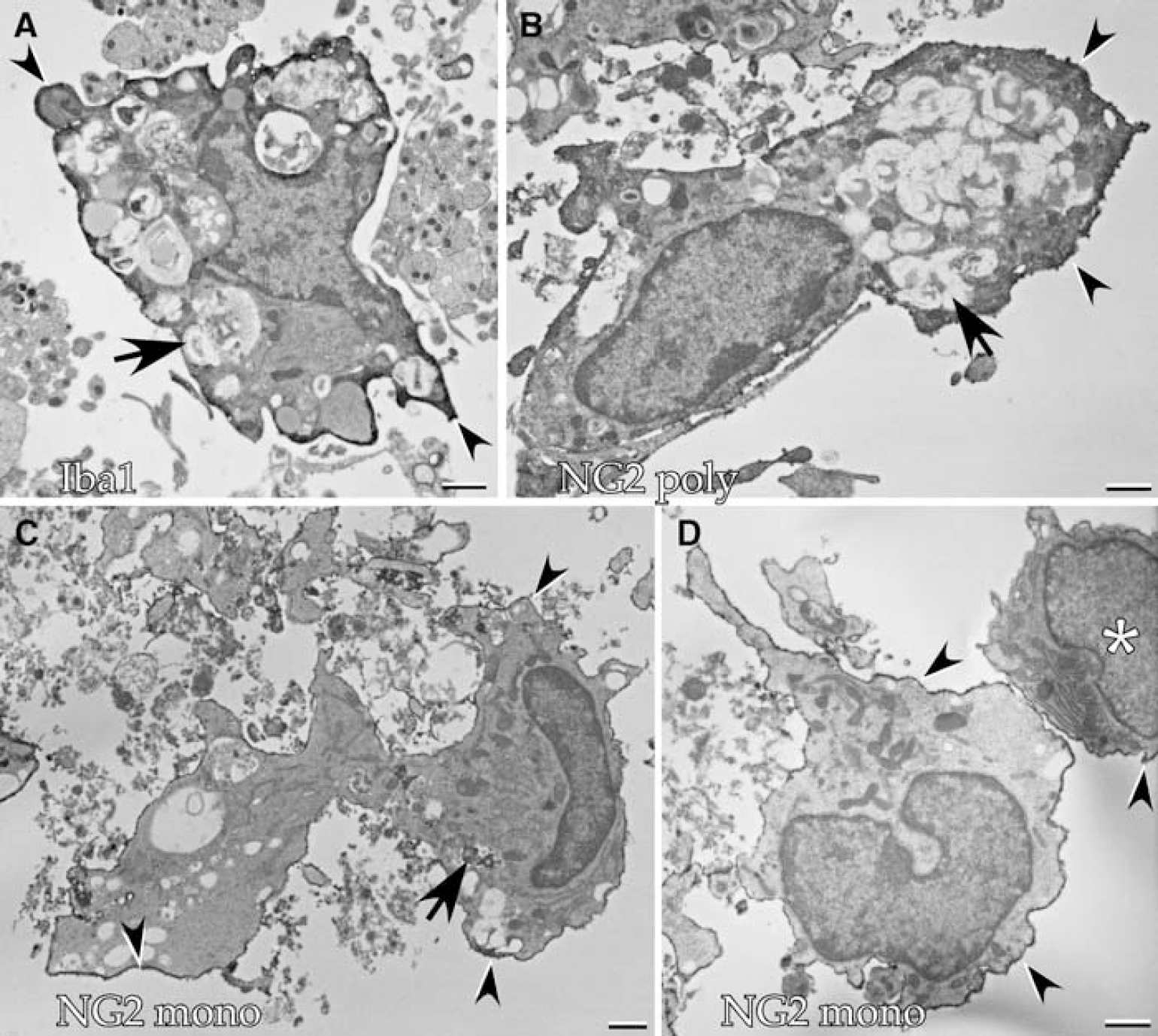
Localizations of NG2 and MG markers in ischemic brain. (
Immunoblot Analysis of Spatial Protein Expression in Ischemic Brain at 7 Days Post-Reperfusion
The presence of Iba1, NG2, PCNA, and MAP2 in ischemic brains was examined by immunoblotting (Figure 4A and 4B). Protein samples were prepared from the contralateral, remote, and core regions at 7 days post-reperfusion. Only a trace amount of MAP2 in the ischemic core and an intermediate amount in the remote region were detected, indicating that the sample preparation was performed correctly. Iba1 expression level increased markedly in ischemic lesions, particularly in the ischemic core. NG2 expression levels were almost the same among the contralateral, remote, and core regions. Note, however, that NG2s from the different regions had different molecular weights. The estimated molecular weights of the two NG2s were 290 and 300 kDa; the 290-kDa NG2 was prominent in the contralateral region, whereas the 300-kDa NG2 in the core region. Proliferating cell nuclear antigen was hardly detected in the contralateral region, and its expression level markedly increased in the core region.
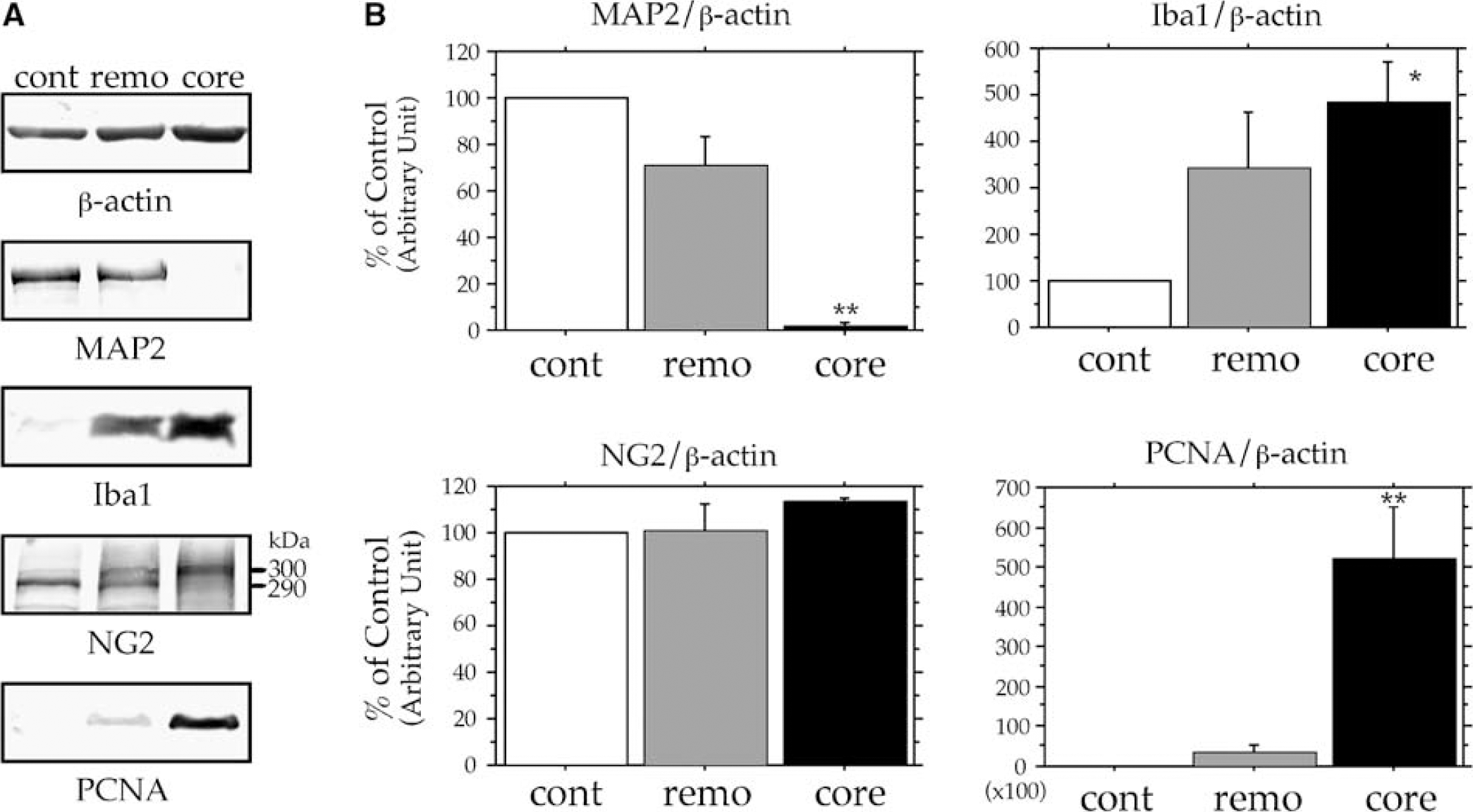
Expressions MAP2, Iba1, NG2, and PCNA in contralateral (cont), remote (remo), and ischemic core (core) regions. The three regions are defined in Figure 1. (
Proliferation of Brain Iba1+/NG2+ Cells in Ischemic Core
The above immunoblotting experiments on PCNA expression showed the presence of highly proliferativecells in the ischemic core. To determine whether such proliferating cells are BINCs, immunohistochemical staining using an anti-Ki67 antibody was performed. Most Iba1+ amoeboid cells were Ki67+ in the ischemic core at 7 days post-reperfusion (Figure 5Aa), whereas Iba1+ cells in the peri-infarct region were Ki67− (Figure 5Ab), indicating that reactive microglia in the peri-infarct region were not proliferative. Most NG2+ amoeboid cells in the core were also Ki67+ (Figure 5B). Therefore, most BINCs in the ischemic core were cycling cells at 7 days post-reperfusion.
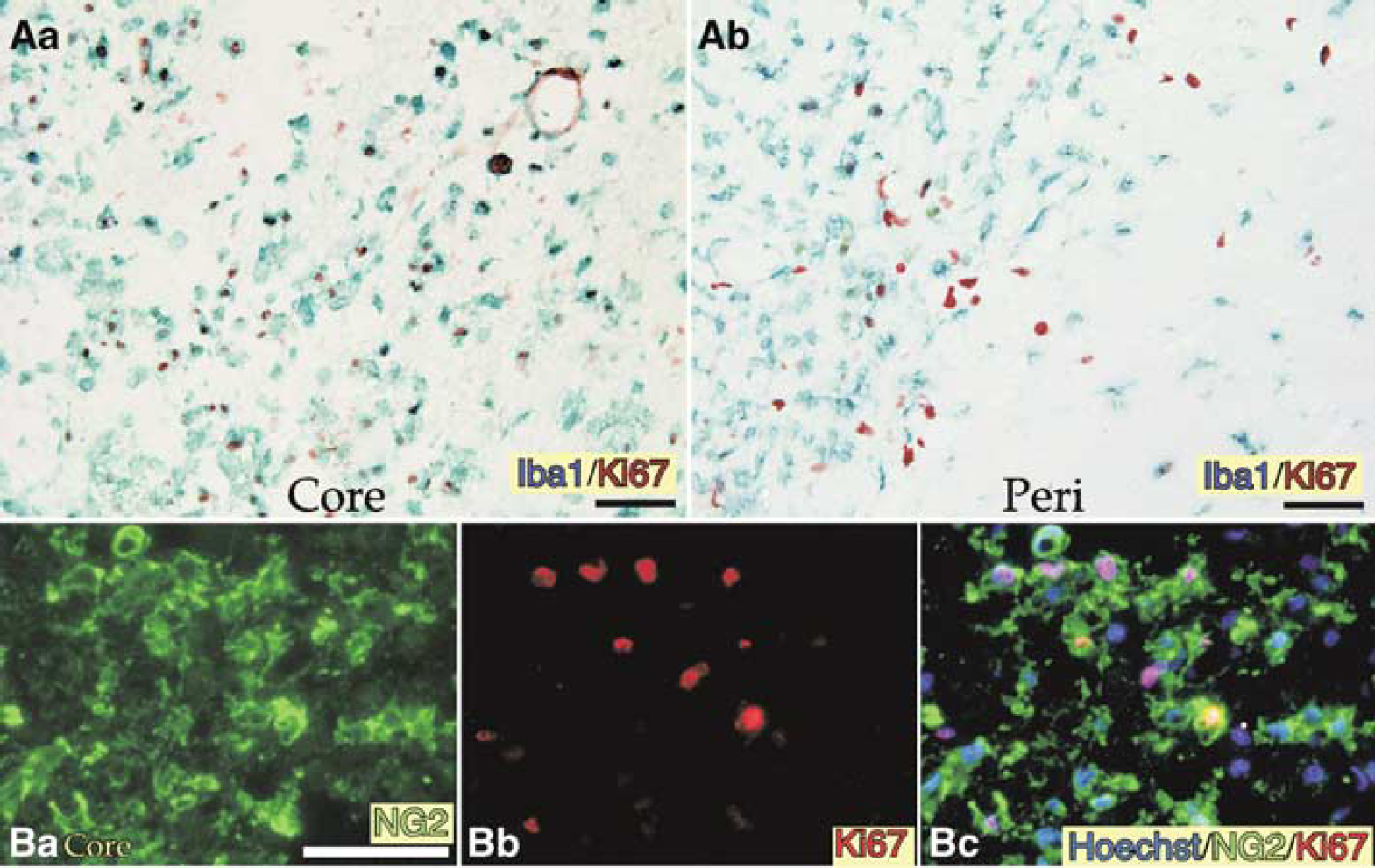
Expressions of Ki67 in Iba1+ or NG2+ cells in the ischemic brain. (
Kinetic Changes in Number of Brain Iba1+/NG2+ Cells in Ischemic Core
Figure 6A shows the kinetic changes in the number of cells in the ischemic core in the cerebral cortex. Although the total number of cells in the ischemic core decreased during the acute phase of ischemic events, it significantly increased at 7 days post-reperfusion and remained constant until 14 days post-reperfusion. Beyond 14 days post-reperfusion, the ischemic tissue in the cortex gradually degenerated and disappeared (data not shown). In the cerebral cortex of the normal brain, both Iba1+ microglia and NG2 glia, which are distinct from each other as shown in Figure 2, account for 5% of all the cells in the cortex (Figure 6B). However, NG2 glia almost completely disappeared at 1 day post-reperfusion in the ischemic core, and about half of the Iba1+ cells also disappeared. Brain Iba1+/NG2+ cells were already present in the ischemic core at 3 days post-reperfusion, as has been described for traumatic brain lesions (Yokoyama et al, 2006). The number of Iba1+ or NG2+ cells gradually increased and reached its maximum at 7 days post-reperfusion. As described above, about half of the cells in the ischemic core at 7 days post-reperfusion were BINCs. The number of BINCs decreased at 14 days post-reperfusion, whereas the total number of cells in the ischemic core remained the same as that at 7 days post-reperfusion.
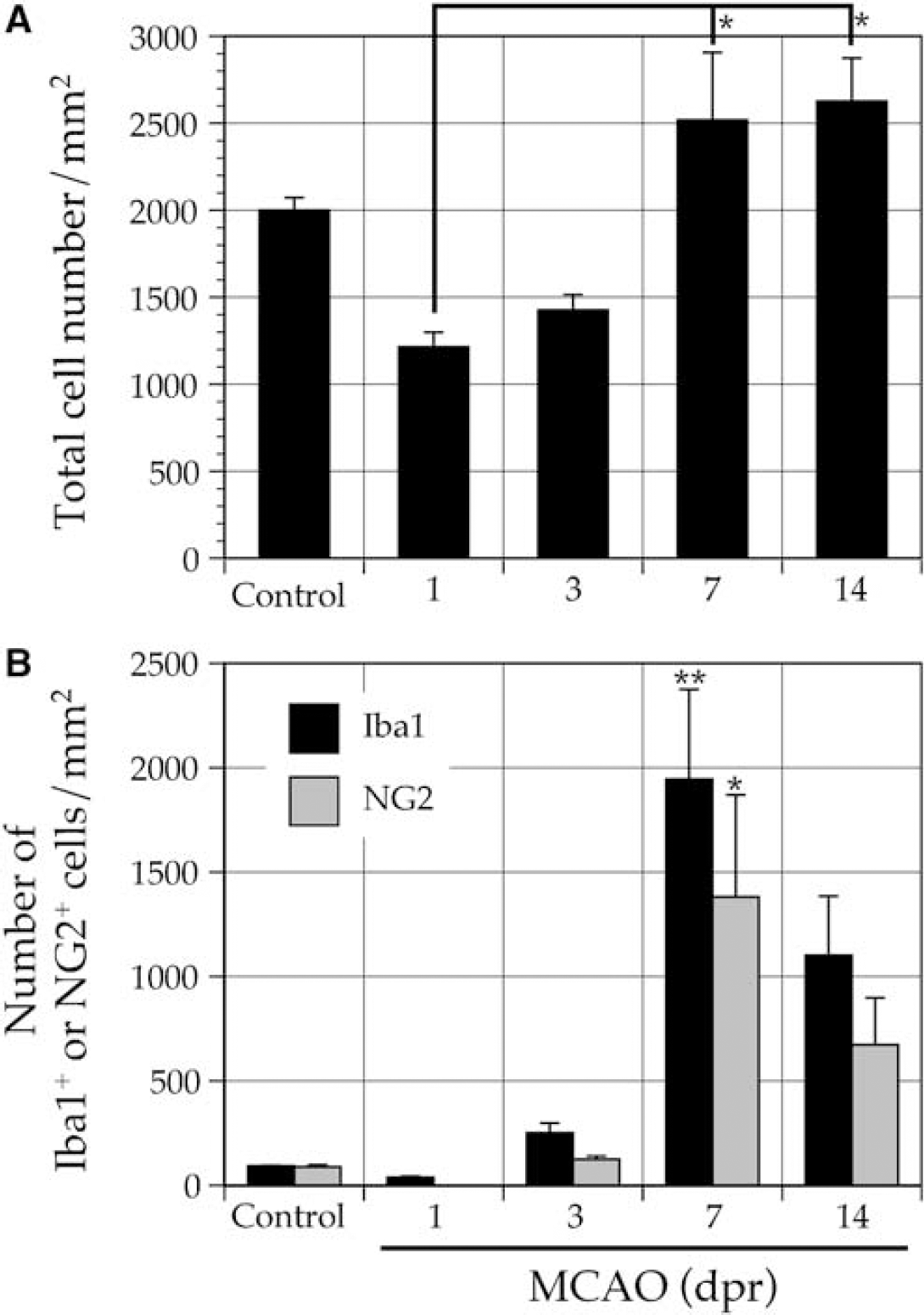
Total number of cells/mm3 percentage of Iba1+ or NG2+ cells in ischemic core (cerebral cortex). Sections from normal and ischemic brains were immunostained with Hoechst 33258 and antibodies to Iba1 and NG2. (A) Total number of cells/mm3 was determined by counting nuclei stained with Hoechst 33258. The data are expressed as mean cell number/mm3. (
Kinetic Changes in Expression Levels of mRNAs and Proteins in Ischemic Brain
The kinetic changes in the expression levels of mRNAs and proteins were investigated in the ischemic core after the onset of ischemic events (Figure 7). The expression levels of mRNAs encoding Iba1 and NG2 increased from 3 to 14 days post-reperfusion, after they decreased at 1 day post-reperfusion (Figure 7A). Immunoblot analysis showed similar results for these molecules including PCNA (Figure 7B and 7C). The expression level of PCNA peaked at 7 days post-reperfusion and was downregulated at 14 days post-reperfusion (Figure 7C), suggesting that the maximum proliferation of BINCs occurs at approximately 7 days post-reperfusion. The expressions of the two NG2s were separately analyzed. As shown in Figure 4, the total NG2 levels in the three regions of the ischemic brains were almost constant. However, the expression level of the 290-kDa NG2 decreased after the onset of the ischemic event, whereas that of the 300-kDa NG2 increased markedly peaking at 7 days post-reperfusion.
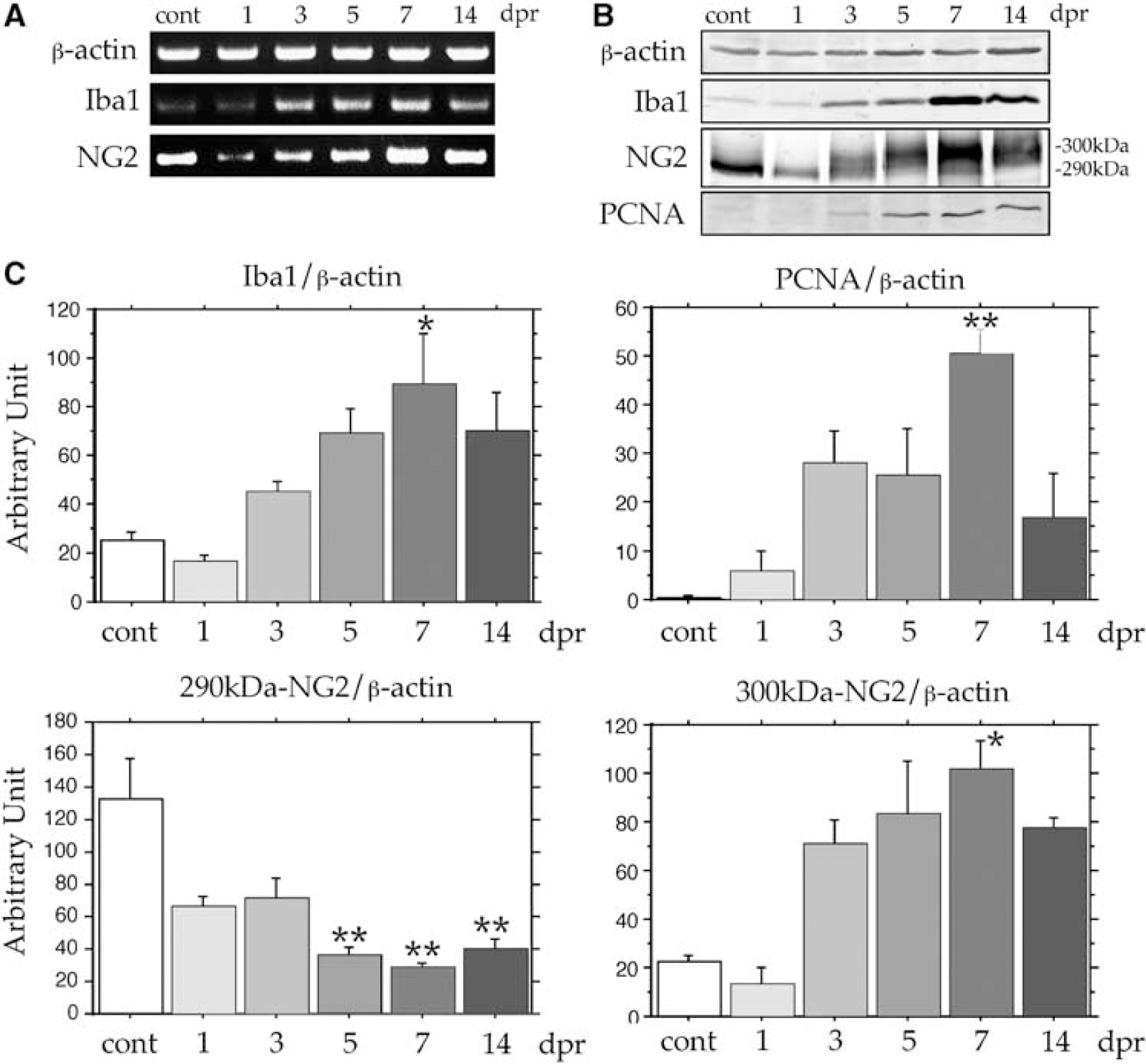
Kinetic study of expressions of Iba1, NG2, and PCNA in the ischemic core from 1 to 14 days post-reperfusion using RT-PCR analysis and immunoblotting. (
Characterization of NG2 Expressed in Core Region
Finally, we investigated molecular basis of the difference in molecular weight between the two NG2s. To clarify the nature of NG2 expressed by BINCs, BINCs were isolated from ischemic lesions of the rat brain at 7 days post-reperfusion. Most of the isolated cells were Iba1+ NG2+, or BINCs (Figure 8). As revealed by immunoblotting experiments using anti-NG2 antibodies, the isolated BINCs expressed only the 300-kDa NG2 (Figure 9A). The use of polyclonal and monoclonal anti-NG2 antibodies produced the same results. To determine whether the difference in molecular weight is due to the difference in glycosylation level, tissue lysates from the contralateral, remote, and core regions were incubated with the chondroitinase ABC and then subjected to immunoblotting with antibodies to NG2 (Figure 9B). The chondroitinase treatment did not apparently change the molecular weights of the major immunoreactive bands. NG2 is also shown to be N-glycosylated (Wen et al, 2006). After treatment with N-glycanase, the molecular weight of NG2 expressed by isolated BINCs decreased by approximately 20 kDa and produced a band of approximately 280-kDa as revealed by immunoblotting using the polyclonal antibody (Figure 9Ca), as has been described elsewhere (Wen et al, 2006). The 280-kDa band was apparently smaller than the 290- and 300-kDa bands. By contrast, when using the monoclonal antibody, the NG2 of BINCs treated with N-glycanase was not observed, suggesting that the monoclonal antibody recognizes the portion including the N-glycosylation site of NG2. Thus, splicing or novel genes might have generated the two core proteins. A gene encoding the NG2 core protein consists of 10 exons (Gibbs et al, 2004). Reverse transcription-polymerase chain reaction experiments using seven sets of primers revealed that all mRNAs derived from the three brain regions contained transcripts from all 10 exons, indicating that the difference in molecular weight between the two NG2s is not due to distinct alternative splicing or novel genes.
Discussion
Expression of NG2 by Brain Iba1+/NG2+ Cells in Ischemic Core
We demonstrated a pronounced accumulation of Iba1+/NG2+ microglia/macrophage-like cells in the ischemic core. Although the close association between the spatial localizations of NG2 glia (or Iba1−/NG2+ cells) and microglia in the brain has been indicated (Nishiyama et al, 1997), Iba1+ cells in the ischemic core expressed NG2 as demonstrated by immunohistochemical analysis using thin sections (4 μm) and by the in vitro immunostaining of isolated Iba1+ cells. Immunoblot analysis showed the presence of the NG2 core protein in the ischemic core and in cells isolated from lesions. The expression of mRNA for the NG2 core protein in the ischemic core was demonstrated by RT-PCR analysis. Resident microglia in the normal mature brain and reactive microglia such as Iba1+ cells with blunted processes in the peri-infarct region were NG2−. Accordingly, cells expressing both Iba1 and NG2 in the ischemic core are distinct from any well-characterized brain cells; therefore, the term ‘BINCs’ was used in this study.
To the best of our knowledge, this is the first report demonstrating the accumulation of NG2+ macrophage-like cells in ischemic brain lesions. Although NG2+ macrophage-like cells or BINCs accumulated in the CNS lesions have been observed for many years, they seem not to have attracted as much attention as NG2 glia. Neurons and glia are believed to inevitably undergo degeneration in the core of ischemic lesions, but they may survive in the region surrounding the core often referred to as the ischemic penumbra. This may be why the study of BINCs in the ischemic core has been unattractive to many researchers. Moreover, BINCs may have simply been recognized as blood—borne cells scavenging dead cells. In fact, BINCs in traumatic brain lesions have phagocytic activity for internalizing infiltrating erythrocytes (Yokoyama et al, 2006). The present immunoelectron microscopy demonstrated that BINCs phagocytosed degenerated materials. However, BINCs and many other types of immune cell including monocytes, neutrophils, and lymphocytes infiltrate the core and survive in it (Kato et al, 1996; Matsumoto et al, 2007a). The accumulation of various immune cells suggests that they play some active roles rather than scavenge cell debris.
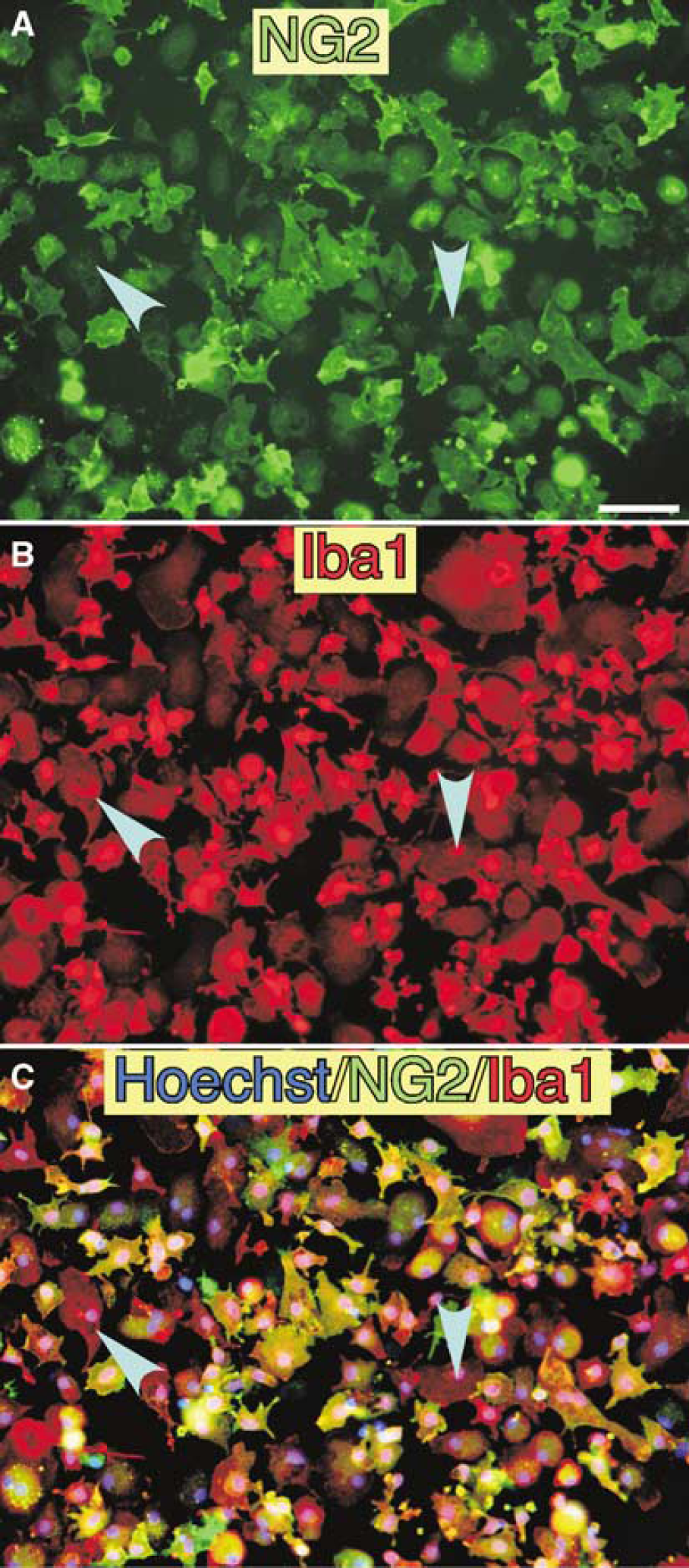
Isolated BINCs from an ischemic hemisphere at 7 days post-reperfusion were double-immunostained with antibodies to (
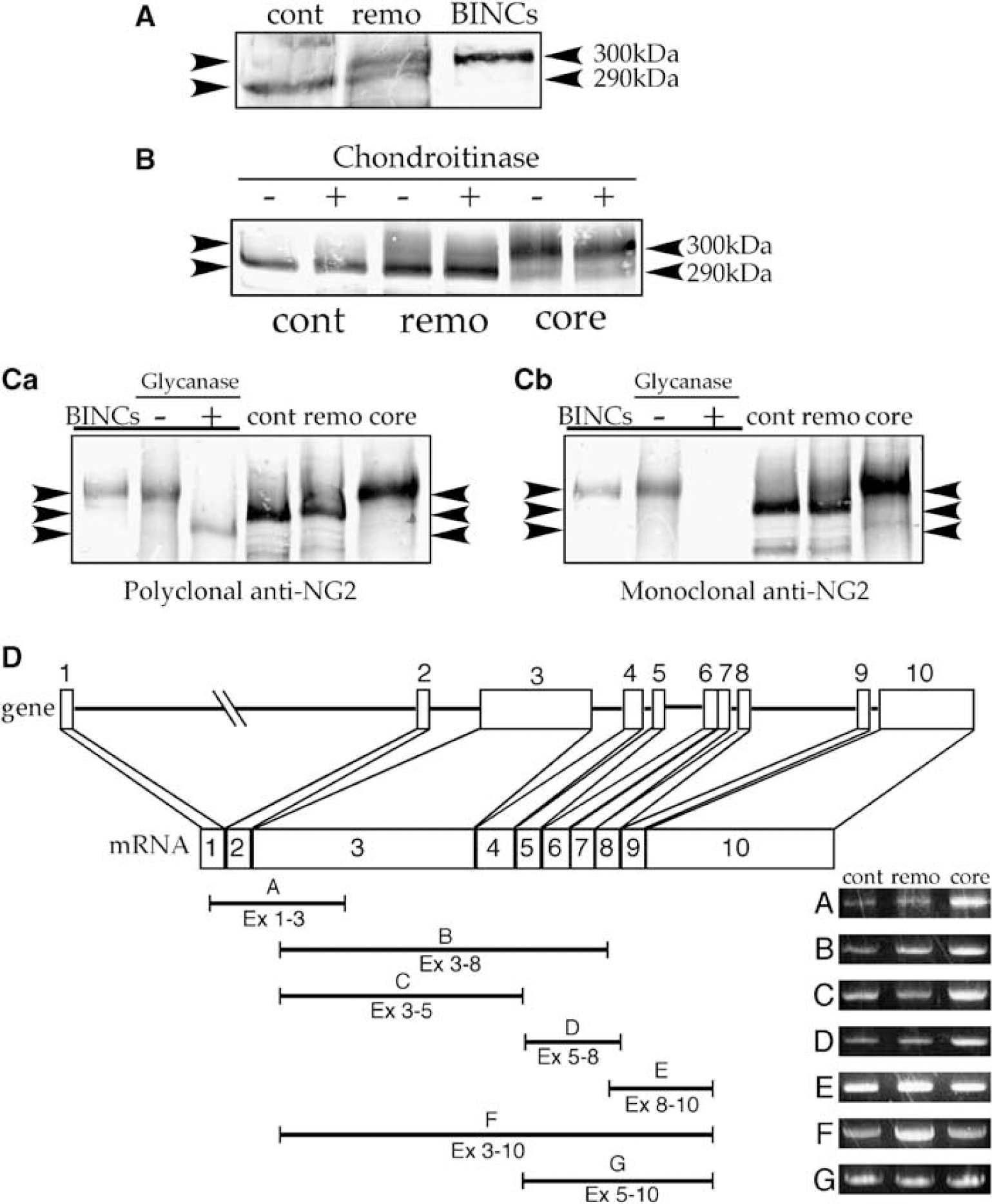
Difference between molecular species of NG2 core protein expressed in ischemic and nonischemic regions. (
Origin of Brain Iba1+/NG2+ Cells in Ischemic Core
Shortly after the reperfusion after MCA occlusion, many resident microglia in the ischemic core undergo apoptotic changes but not activation (Matsumoto et al, 2007a). This is also shown in the present cell counting experiments (Figure 6). At the same time, resident NG2 glia also disappear and neutrophils markedly infiltrate the ischemic core within 24 h post-reperfusion. The invasion of monocytes also becomes observable in the ischemic core at approximately 24 h post-reperfusion. However, the detection of BINCs was rather late, starting at 3 days post-reperfusion. Such a time course of changes in the major cell type present in the ischemic core suggests that BINCs are derivatives of neither resident microglia nor NG2 glia. Therefore, BINCs in the ischemic core may be blood-borne cells. This is also based on the results of studies employing the transplantation of the bone marrow of green fluorescent protein-transgenic mice (Tanaka et al, 2003). Bu et al (2001) have shown that NG2+ macrophage-like cells are absent in damaged brain slices in culture but are present in brain lesions with similar damage, suggesting that the cells are blood-borne.
However, no NG2+ cells exist in normal blood or bone marrow; they have been found only in blood of patients with special types of acute leukemia (Neudenberger et al, 2006). Therefore, bone marrow-derived precursors may first express NG2 after their invasion of brain lesions. In addition, Iba1+/NG2+ cells appear in traumatically damaged liver in the acute phase, although the accumulation of cells is much less pronounced than that in the ischemic core of the brain (data not shown). Accordingly, damaged tissues may produce some signal molecules that induce NG2 expression in Iba1+ monocyte-like cells.
Roles of Brain Iba1+/NG2+ Cells in Ischemic Core
As described above, the most prevalent view on the function of microglia or macrophages that accumulate in ischemic lesions may be in terms of scavenging cell debris (Kreutzberg, 1996). Accumulation of macrophage-like cells expressing a phagocytic cell marker CD68 is also reported in cases of human beings (Postler et al, 2000). The continuous proliferation of BINCs in the ischemic core, however, suggests their novel roles in lesions beyond their classic roles as macrophages. In fact, there were many BINCs that contained no phagosomes in their cytoplasm (Figure 3D). As one of such possible novel roles of BINCs in the pathologic brain, involvement in regeneration of damaged brain may be proposed, in light of recent findings suggesting that some populations of monocytes or macrophages have a multipotent ability to generate various cells, including endothelial cells, hepatocytes, and neurons (Takahashi et al, 1999; Tonchev et al, 2003; Urbich and Dimmeler, 2004; Zhao and Mazzone, 2005). Furthermore, NG2+ primary microglia and BINCs isolated from traumatic brain lesions give rise to cells with neuroectodermal phenotypes after the formation of cell aggregates in a culture medium containing 70% serum (Yokoyama et al, 2004, 2006). The number of BINCs peaked at 7 days post-reperfusion and decreased at 14 days post-reperfusion Figure 6), whereas the total number of cells was constant from 7 to 14 days post-reperfusion. Besides, no TUNEL+ BINCs were observed from 3 to 28 days post-reperfusion (data not shown). The cells that increased in number in the ischemic core appeared to be GFAP+. These observations indicate that BINCs serve as progenitors giving rise to GFAP+ cells. In fact, a decrease in the number of BINCs and a simultaneous increase in the number of GFAP+ cells were observed in the core of stab wounds (Yokoyama et al, 2006), although few GFAP+ cells were marked with bromodeoxyuridine in CNS lesions (Okada et al, 2006). BINCs likely express nestin, an intermediate filament protein that is expressed in multipotent cells including neural stem cells, based on a report demonstrating that proliferative cells in ischemic lesions express nestin and NG2 (Kronenberg et al, 2005).
Collectively, BINCs might give rise to both neuroectodermal and mesodermal cells leading to regeneration of damaged brain tissues. BINCs accumulate in lesion core, where they should eventually disappear within weeks or months after the onset of brain damage. An appropriate control of proliferation and differentiation of BINCs in the lesion core might prevent the loss of brain tissues.
Our recent study revealed that macrophage-like cells expressing CD200 but not NG2 accumulate simultaneously with BINCs in ischemic core (Matsumoto et al, 2007b). Iba1+/NG2− cells that formed one-third of macrophage-like cells or Iba1+ cells in ischemic core (Table 3) were CD200+. CD200, a protein belonging to immunoglobulin superfamily, is expressed mainly in lymphoid cells and involved in suppression of myeloid cells including microglia that express CD200 receptor (CD200R) (Hoek et al, 2000). In ischemic core, CD200R is expressed in BINCs (unpublished results). Therefore, the interaction between CD200+, Iba1+/NG2− cells, and CD200R+ BINCs may play some roles in regulation of proliferation and differentiation of these macrophage-like cells through secretion of cytokines and growth factors.
Molecular Bases Underlying Two NG2s
The apparent molecular weight of the dominant NG2 species expressed in BINCs in the ischemic core was 300 kDa, whereas that of the NG2 found in the nonischemic region was approximately 290 kDa. The difference in molecular weight may not be due to the glycosaminoglycan side chain based on the results of the present experiments using chondroitinase ABC and N-glycanase. Furthermore, the monoclonal antibody that may recognize N-glycosylation site of NG2 immunostained both NG2 glia and BINCs, indicating that both NG2s are N-glycosylated. To determine whether alternative splicing or novel genes are responsible for this difference, detailed RT-PCR analysis was performed. Consequently, no evidence was found showing the involvement of any specific alternative splicing or novel genes in the generation of the two NG2s. Collectively, the difference in molecular weight may have arisen from the post-translational partial degradation of the NG2 core protein, but not the glycosylation of the core protein. It is likely that such post-translational modification of the core protein is simply the shedding of the extracellular domain of the NG2 core protein (Asher et al, 2005; Stallcup, 2002).
The NG2 core protein has three domains (Nishiyama et al, 1991): a large extracellular domain, a transmembrane domain, and a short cytoplasmic tail. The NG2 core protein has a molecular weight of 300 kDa, and the smaller 290-kDa species may be secreted by cells by shedding the extracellular domain (Asher et al, 2005). Thus, BINCs in ischemic core likely have NG2 as a transmembrane protein, whereas NG2 glia as OPCs in the nonischemic region of mature rats release most NG2 by shedding the extracellular domain. Thus, this study showed that NG2 glia and BINCs could be distinguished by their NG2 species. The 300-kDa NG2 may be a good marker of BINCs.
NG2 is a unique transmembrane protein capable of binding to growth factors and extracellular matrix proteins. The extracellular domain of NG2 can bind to PDGF-AA or basic fibroblast growth factor (Goretzki et al, 1999) and sequester them at the cell surface. Besides, NG2 can form a complex with PDGFRa. Cultured OPCs or NG2 glia expressing a receptor of PDGF-AA (or PDGFRa) cannot proliferate in response to PDGF-AA when treated with a blocking antibody to NG2. Thus, NG2 functions in signaling via growth factors. Considering the marked proliferation of BINCs in the ischemic core, NG2 might be responsible for their active proliferation through the sequestration of growth factors. In fact, mRNA encoding PDGF-A was expressed at high levels in ischemic core when the active proliferation of BINCs was observed (data not shown). However, it was not clear whether BINCs expressed PDGFRa. Even if BINCs expressed the receptor, the expression level was rather low (data not shown). Thus, the role of the 300-kDa NG2 in BINCs remains to be elucidated.
Footnotes
Acknowledgements
We thank Professor William B Stallcup for his critical suggestions and providing antibodies to NG2.