
Abstract
Acute hypoxia directly causes cerebral arteriole vasodilation and also stimulates peripheral chemoreceptors to change autonomic neural activity. These changes may alter cerebral vascular modulation. We therefore hypothesized that dynamic cerebral autoregulation would be altered during acute exposure to hypoxia. Fifteen healthy men were examined under normoxic (21%) and hypoxic conditions. Oxygen concentrations were decreased in stepwise fashion to 19%, 17%, and 15%, for 10 mins at each level. Mean blood pressure (MBP) in the radial artery was measured via tonometry, and cerebral blood flow velocity (CBFV) in the middle cerebral artery was measured by transcranial Doppler ultrasonography. Dynamic cerebral autoregulation was assessed by spectral and transfer function analysis of beat-by-beat changes in MBP and CBFV. Arterial oxygen saturation decreased significantly during hypoxia, while end-tidal CO2 and respiratory rate were unchanged, as was steady-state CBFV. With 15% O2, very-low-frequency power of MBP and CBFV variability increased significantly by 185% and 282%, respectively. Moreover, transfer function coherence (21% O2, 0.46 ± 0.04; 15% O2, 0.64 ± 0.04; P = 0.028) and gain (21% O2, 0.61 ± 0.05 cm/secs/mm Hg; 15% O2, 0.86 ± 0.08 cm/secs/mm Hg; P = 0.035) in the very-low-frequency range increased significantly by 53% and 48% with 15% O2, respectively. However, these indices were unchanged in low- and high-frequency ranges. Acute hypoxia thus increases arterial pressure oscillations and dependence of cerebral blood flow (CBF) fluctuations on blood pressure oscillations, resulting in apparent increases in CBF fluctuations in the very-low-frequency range. Hypoxia may thus impair dynamic cerebral autoregulation in this range. However, these changes were significant only with hypoxia at 15% O2, suggesting a possible threshold for such changes.
Keywords
Introduction
The number of individuals going to high altitude or engaging in hypoxic training has been increasing (Honigman et al, 1993; Levine and Stray-Gundersen, 1997). High-altitude physiology has become more important, particularly with regard to the cerebral autoregulation related to mechanisms of acute mountain sickness (AMS) (Otis et al, 1989; Van Osta et al, 2005).
Cerebral autoregulation adjusts cerebral arteriolar caliber (or cerebrovascular resistance) to guarantee blood supply to the brain. Cerebral blood flow (CBF) is maintained relatively independent from changes in cerebral perfusion pressure or systemic arterial blood pressure (ABP). Originally, studies of cerebral autoregulation focused on ‘steady-state' relationships between mean values of ABP and CBF, establishing a plateau of CBF for ABP values between 60 and 150 mm Hg (Paulson et al, 1990). Recently, the focus of research on cerebral autoregulation has shifted to transient, or ‘dynamic‘, responses of CBF to changes in ABP, as high temporal resolution became achievable using transcranial Doppler ultrasonography in the late 1980s (Aaslid et al, 1989). Neural pacemakers in the brain, feedback loops to central regions, or local vascular controls may cause CBF oscillations or may conversely dampen CBF oscillations induced by ABP oscillations. To clarify these issues, a Fourier-based signal processing technique and transfer function analysis between spontaneous oscillations of ABP and CBF are employed. Details of this new method have been carefully explained and reviewed by Zhang et al (1998) and Panerai et al (2005), and are provided in the Materials and methods. Briefly, transfer-function gain depends on frequency alone, as does mean squared coherence, with both being computed from ABP and CBF as functions of time. Coherence indicates the strength of association between ABP and CBF velocity variabilities. A lower coherence is generally interpreted as indicating that signals are independent of each other, suggesting effective autoregulation. Conversely, higher coherence means that pressure and velocity vary together very closely and may indicate a greater dependence of velocity oscillations on pressure oscillations. Transfer-function gain expresses the relative amplitude of the relationship between these two variables. A larger gain implies that any given changes in pressure lead to larger changes in flow.
New insights into dynamic cerebral autoregulation have arisen from studies using this new approach in various clinical conditions where cerebral autoregulation could be expected to be impaired, such as autonomic failure (Blaber et al, 1997), occlusive cerebrovascular diseases (Diehl et al, 1995), carotid stenosis (Hu et al, 1999), hypertension (Lipsitz et al, 2000), brain injury (Muller et al, 2003), migraine (Muller and Marziniak, 2005), stroke (Reinhard et al, 2005), and syncope (Schondorf et al, 2001). Although complete consensus has not yet been reached regarding the notion of ‘normal’ and ‘impaired' dynamic cerebral autoregulation, numerous reports have shown common characteristics of frequency-dependent phenomena in healthy humans. For example, estimates of coherence function are reduced at lower frequency ranges, and drop to near or below 0.5 for frequencies < 0.1 Hz, signifying minimal correlations between these variables or little dependence of CBF oscillations on ABP oscillations (Eames et al, 2004; Edwards et al, 2002; Kuo et al, 1998; Muller and Marziniak, 2005; Ogawa et al, 2006; Panerai et al, 1999, 2005; Schondorf et al, 2001; Zhang et al, 1998, 2002). Moreover, several previous studies examining the effects of various interventions on dynamic cerebral autoregulation have shown that each intervention alters the dynamic relation or dependence of CBF oscillations on ABP oscillations in a specific frequency interval (Ogawa et al, 2006, Ogoh et al, 2005; Zhang et al, 2002). However, this new approach has rarely been applied to learning more about hypoxia, and the present study was therefore designed.
Acute hypoxia produces vasodilation in cerebral vascular beds (Van Mil et al, 2002; Wadsworth, 1994). Stimulation of peripheral chemoreceptors by acute hypoxia also changes autonomic nervous activity (Hughson et al, 1994; Saito et al, 1988), which is one of the cerebral vascular regulatory systems (Zhang et al, 2002). Furthermore, one preliminary report showed that chronic exposure to high altitude increases transfer function coherence and gain in the very-low-frequency range, suggesting altered dynamic autoregulation (Levine et al, 1999). We therefore hypothesized that CBF oscillations and the dynamic relationship between ABP and CBF oscillations would be altered by acute exposure to hypoxia in a dose-dependent manner. To test this hypothesis, we examined spontaneous variabilities in mean blood pressure (MBP) and CBF velocity, and analyzed dynamic relationships between these two variabilities during stepwise exposure to hypoxia.
Materials and methods
Subjects
Subjects comprised 15 healthy mean (mean (± s.d.) age, 22 ± 2 years; mean height, 172 ± 5 cm; mean weight, 62 ± 5 kg). No subject had any known medical problems. Subjects were screened carefully based on medical history and physical examination, including 12-lead echocardiography. All subjects provided written informed consent and all protocols were approved by the Ethical Committee of Nihon University School of Medicine.
Instrumentation
All experiments were performed ≥ 2 h after a meal in a quiet, environmentally controlled laboratory at an ambient temperature of 23°C to 25°C. Subjects refrained from heavy exercise and caffeinated or alcoholic beverages for ≥ 24 h before testing. Subjects were studied using a Hypoxico hypoxic tent system (Hypoxico, NY, USA).
End-tidal carbon dioxide (ETCO2) and arterial oxygen saturation (SaO2), monitored using a NPB-75 (Nellcor, CA, USA), were recorded every minute. Intermittent blood pressure was measured by oscillometric determination using a Jentow CS cuff sphygmomanometer (Colin Medical Technology, Aichi, Japan) over the brachial artery every 5 mins. Analog echocardiography was obtained from a CM5 lead. Continuous ABP was obtained at the right radial artery by tonometry using the Jentow CS at heart level on a beat-by-beat basis. Cerebral blood flow velocity in the middle cerebral artery was obtained by transcranial Doppler ultrasonography (WAKI; Atys Medical, St Genislaval, France) at a depth of 50 to 60 mm using a 2-MHz transducer. The transducer was positioned on the temporal window and fixed, at a constant angle with a probe holder that was made with a polymer mold to fit the facial bone structure and ear of each subject (Giller and Giller, 1997). The right middle cerebral artery was insonated according to the angle with the highest velocity and best-quality Doppler signal.
Protocols
Stepwise protocol (21%, 19%, 17%, and 15% O2): after ≥30 mins of supine rest, data were measured for initial room air (21% O2 baseline). O2 concentration was then decreased in a stepwise manner to 19% (effective altitude, 800 m), 17% (effective altitude, 1700 m), and 15% (effective altitude, 2600 m) using large Douglas bags (300 L) filled with 12.1% O2 gas and the hypoxic generator. Each oxygen concentration was maintained for 10 mins by the hypoxic generator and data were collected during the last 5 mins in each step. O2 levels were maintained at the target concentration ± 0.2% during hypoxic exposure, while CO2 concentration in the tent was kept < 0.1% by circulating air using a suction pump (30 L/mins) through a CO2 absorber placed in the tent. Arterial blood pressure, echocardiography and CBF velocity were recorded in 5-mins segments at a sampling rate of 1 kHz for spectral and transfer function analysis with spontaneous respiration for all data collection. O2 saturation, ETCO2, and respiratory rate were recorded every 1 min during data collection.
Time-control protocol (21% O2 10-, 20-, 30-, 40-mins): The same experiments as the stepwise protocol were performed except for oxygen concentration. Instead of hypoxic air, 21% O2 air was used for this protocol. This protocol was performed to evaluate the possible effects of time factors on these experimental conditions.
Single-dose protocol (21% O2, 15% O2): Subjects comprised nine of the 15 original subjects (mean age, 21 ± 2 years; mean height, 175 ± 4 cm; mean weight, 65 ± 7 kg). This protocol was performed to evaluate any possible influences of preceding steps in stepwise exposure to hypoxia. After ≥ 30 mins of supine rest, data were measured for initial room air (21% O2). O2 concentration was then decreased directly to 15% within 5 mins using the hypoxic generator and large Douglas bags (300 L) filled with 12.1% oxygen gas, and the 15% oxygen concentration was maintained by the hypoxic generator. Data were collected for 5 mins after oxygen concentration reached 15%.
Analysis
Beat-to-beat MBP and CBF velocity were obtained by integrating signals within each cardiac cycle using PC-based Notocord-hem 3.3 software (Notocord, Paris, France) (Figure 1). Beat-to-beat data for MBP and CBF velocity were then linearly interpolated and re-sampled at 2 Hz for spectral and transfer function analysis. Fast Fourier Transform and transfer function analysis were performed using a Hanning-window on 256-point segments with 50% overlap, to assess the dynamic pressure-flow relationship (Figure 2) (Kuo et al, 1998; Lipsitz et al, 2000; Panerai et al, 1999; Zhang et al, 1998, 2002). This data analysis was performed using DADiSP software (DSP Development, Cambridge, MA, USA).
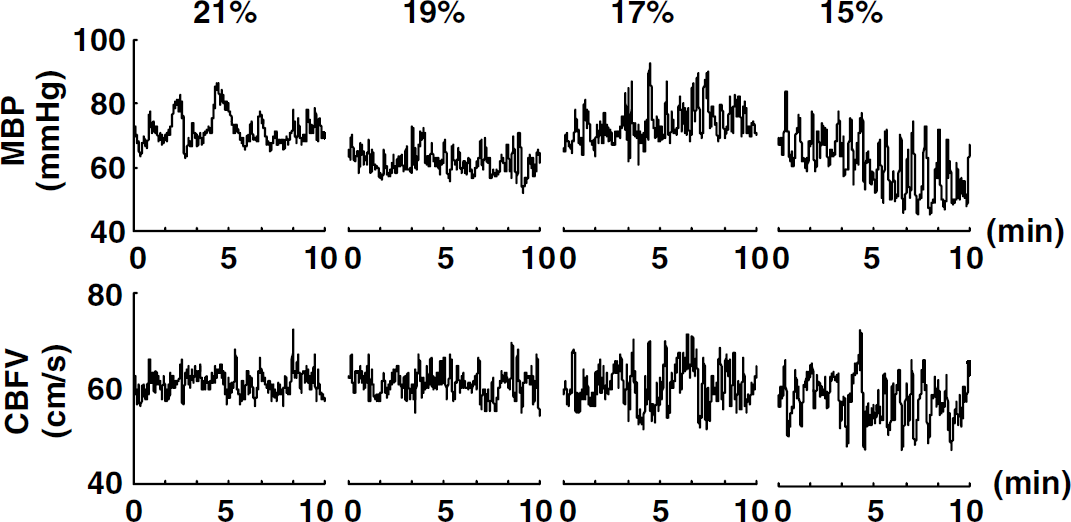
Oscillations in representative data of the time course for mean blood pressure and CBFV in a subject for the full 10 mins of 21% O2 baseline and the three hypoxia stages (19%, 17%, and 15% O2 in Stepwise protocol).
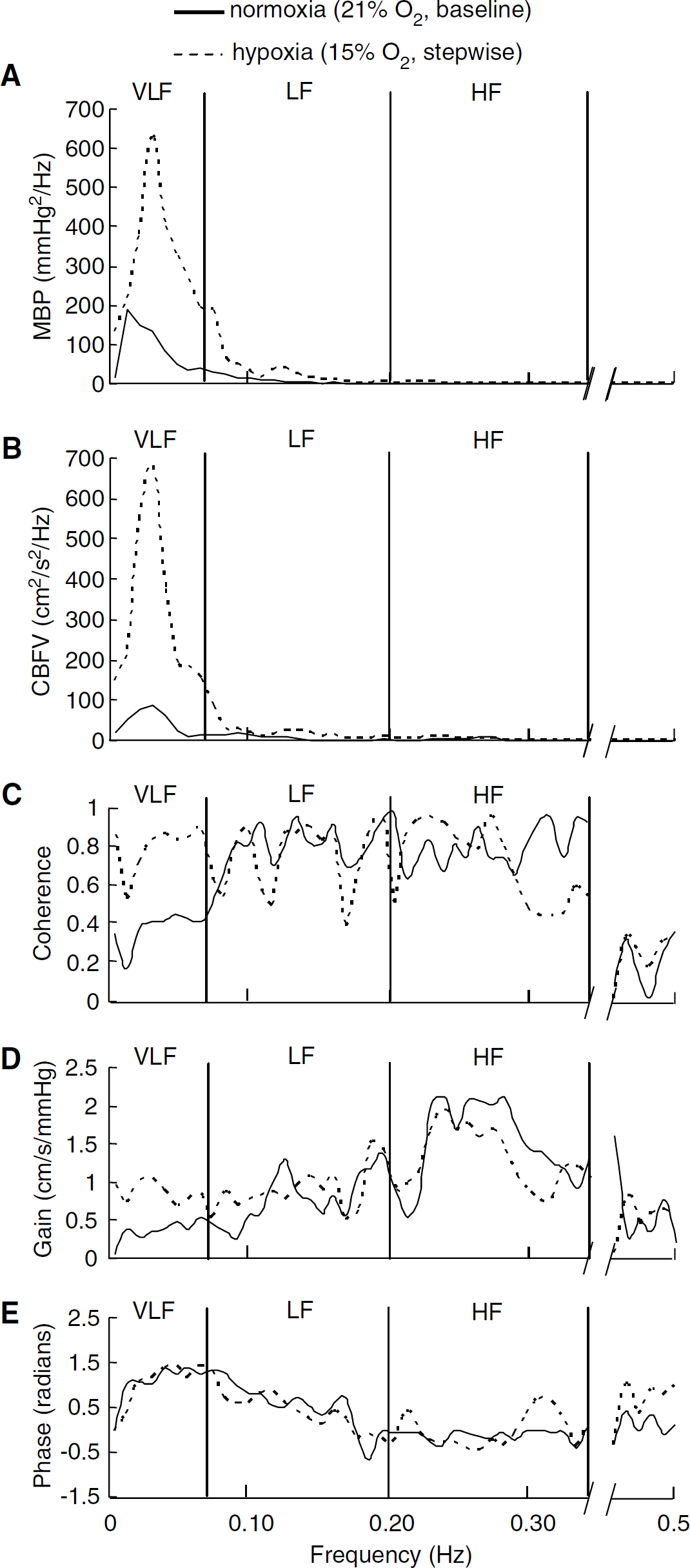
Representative frequency-domain analysis of changes in MBP and CBFV in a subject before (21% O2 baseline) and during hypoxia (15% O2 of Stepwise protocol). (
Transfer function H(f) between the two signals was defined as
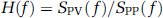
where SPP(f) is the Fourier-transformed autocorrelation function (autospectrum) of changes in MBP and SPV(f) is the Fourier-transformed cross-correlation function (cross-spectrum) between MBP and CBF velocity signals. Transfer function magnitude |H(f)| and phase spectrum Φ(f) were derived from the real part [HR(f)] and imaginary part [HI(f)] of the complex function |H(f)| as
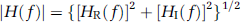
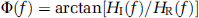
The mean squared coherence (MSC) function MSC(f) was defined as:
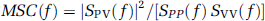
where SVV(f) is the autospectrum of CBF velocity.
Mean MSC for a particular frequency interval represents the ‘coherence' for that interval. Spectral power of MBP and CBF velocity, mean value of transfer function gain, phase, and coherence were calculated in the very-low-(0.02 to 0.07 Hz), low- (0.07 to 0.20 Hz), and high-(0.20 to 0.35 Hz) frequency ranges. These ranges were specifically chosen to reflect different patterns of the dynamic pressure—flow relationship, as previously identified by transfer function analysis (Zhang et al, 1998, 2002).
Coherence can range from 0 to 1. Linear relationships are characterized by coherence values near 1. Low coherence values (< 0.5) generally characterize complex, nonlinear systems, particularly those where output can be influenced by multiple inputs. Low coherence (< 0.5) has generally been attributed to tight autoregulation, with shifts towards 1.0 implying poor autoregulation. Another measure relevant to linearity and autoregulation is the gain. For a given frequency range, gain gives the extent to which stimulus is transmitted to response in that frequency range. Worsening of dynamic cerebral autoregulation should thus lead to both increased coherence and transfer function gain, reflecting the increased influence of ABP oscillations on CBF velocity oscillations. Phase shift was used to estimate temporal relationships between these two variables.
Statistical Analysis
Results are presented as mean ± s.e.m. To strengthen the experimental design of repeated measures with the same 15 subjects acting as their own controls, and to compare possible changes in baseline data between the two conditions, variables were compared using two-way repeated-measures analysis of variance within subject Condition (stepwise hypoxia, time-control) × Stage (21% to 15%, 10 to 40 mins) (n = 15) or within subject Condition (stepwise hypoxia, single-dose hypoxia) × Stage (21%, 15%) (n = 9). The interaction effect was considered the most relevant for determining any effect of hypoxia. To determine where significant differences occurred, Bonferroni t-tests were conducted. Values of P < 0.05 were considered statistically significant. Original degrees of freedom and P-values are reported.
Results
Stepwise Protocol and Time-Control Protocol
No significant changes were noted for any indices in the time-control protocol (Tables 1–3).
Respiratory conditions
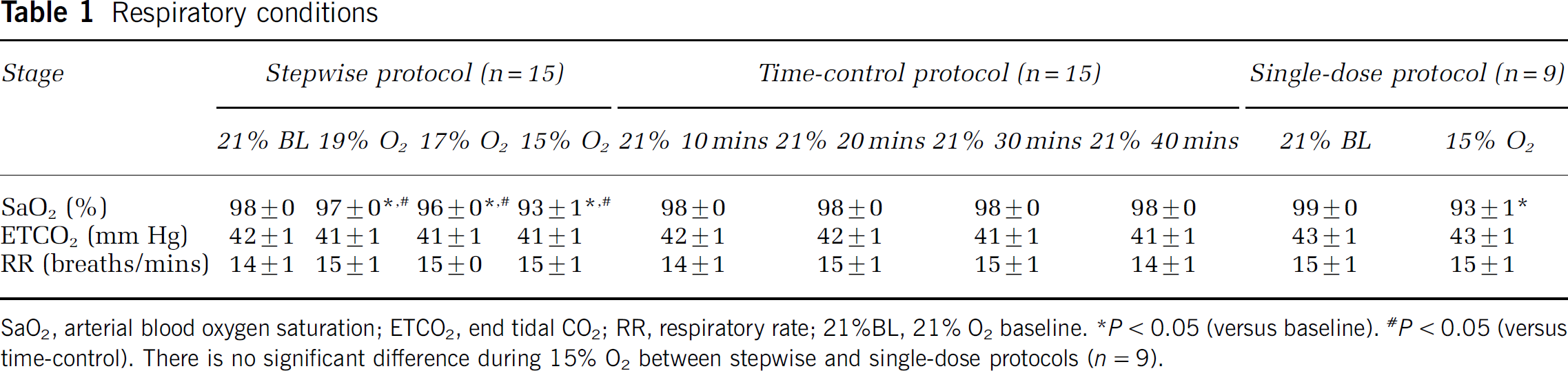
SaO2, arterial blood oxygen saturation; ETCO2, end tidal CO2; RR, respiratory rate; 21%BL, 21% O2 baseline.
P < 0.05 (versus baseline).
P < 0.05 (versus time-control). There is no significant difference during 15% O2 between stepwise and single-dose protocols (n = 9).
Steady-state hemodynamics
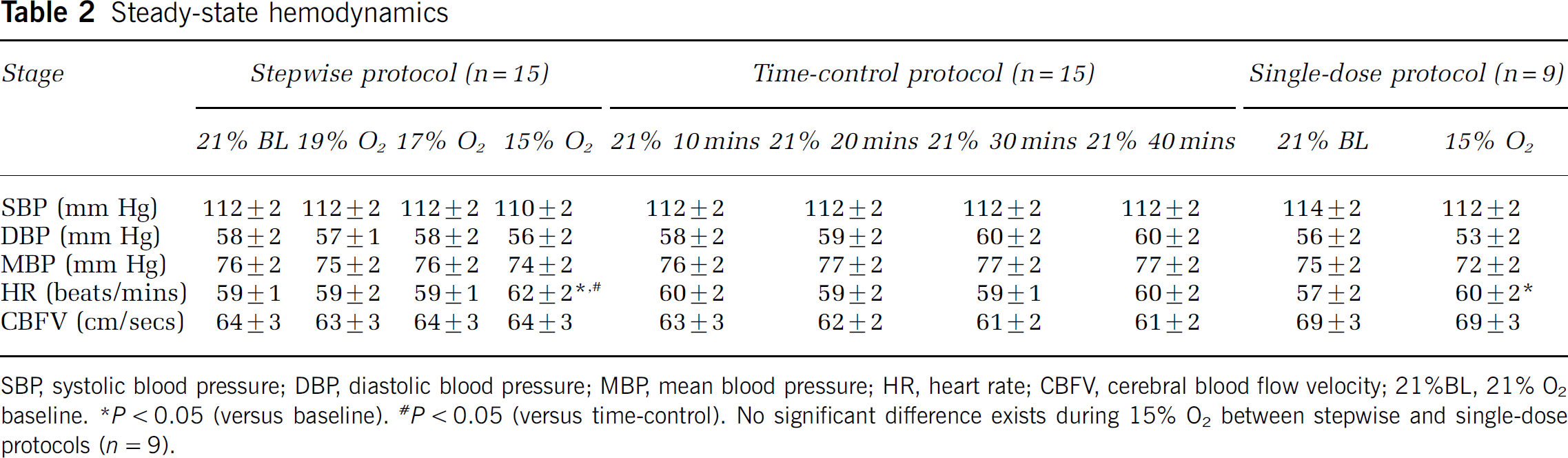
SBP, systolic blood pressure; DBP, diastolic blood pressure; MBP, mean blood pressure; HR, heart rate; CBFV, cerebral blood flow velocity; 21%BL, 21% O2 baseline.
P < 0.05 (versus baseline).
P < 0.05 (versus time-control). No significant difference exists during 15% O2 between stepwise and single-dose protocols (n = 9).
Spectral analysis of MBP variability and CBFV variability
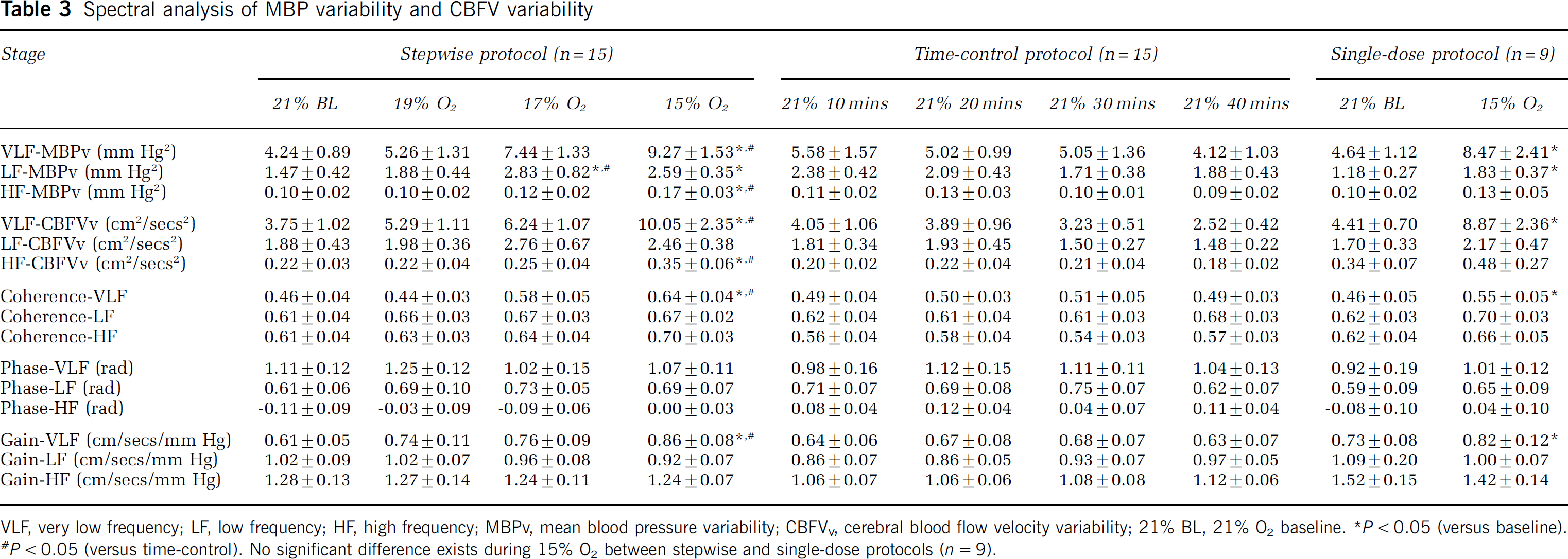
VLF, very low frequency; LF, low frequency; HF, high frequency; MBPV, mean blood pressure variability; CBFVV, cerebral blood flow velocity variability; 21% BL, 21% O2 baseline.
P < 0.05 (versus baseline).
P < 0.05 (versus time-control). No significant difference exists during 15% O2 between stepwise and single-dose protocols (n = 9).
As expected, SaO2 decreased significantly (significant interaction effect, F3,42 = 121.784, P < 0.001), while ETCO2 and respiratory rate did not change during acute hypoxia (Table 1). Diastolic blood pressures and MBP, tended to decrease with hypoxia at 15% O2, but no significant difference was noted (no significant interaction effects, diastolic blood pressures, F3,42 = 2.572, P = 0.067; MBP, F3,42 = 2.059, P = 0.12). Cerebral blood flow velocity did not change. Heart rate increased significantly with hypoxia at 15% O2 and heart rate at 15% O2 was significantly higher than that at 21% O2 for 40 mins in the time-control protocol (significant interaction effect, F3,42 = 4.410, P = 0.009) (Table 2).
Changes in spectral and transfer function indices are shown in Table 3. Mean blood pressure variability in low- and high-frequency ranges increased (significant interaction effects, low frequency, F3,42 = 4.077, P = 0.013; high frequency, F3,42 = 2.845, P = 0.049). However, CBF velocity variability at low frequency remained unchanged. Cerebral blood flow velocity variability at high frequency increased significantly with hypoxia at 15% O2 (significant interaction effect, F3,42 = 4.723, P = 0.006).
Mean blood pressure variability at very low frequency increased significantly by 185% ± 43% with 15% O2, and this index at 15% O2 was significantly higher by 257% ± 92% than that at 21% O2 for 40 mins in the time-control protocol (significant interaction effects, very low frequency, F3,42 = 4.452, P = 0.008). Cerebral blood flow velocity variability at very low frequency increased significantly by 282% ± 73% with hypoxia at 15% O2, and this index at 15% O2 was significantly higher by 562% ± 190% than that at 21% O2 for 40 mins in the time-control protocol (significant interaction effect, F3,42 = 5.931, P = 0.002) (Figures 1 and 3A).
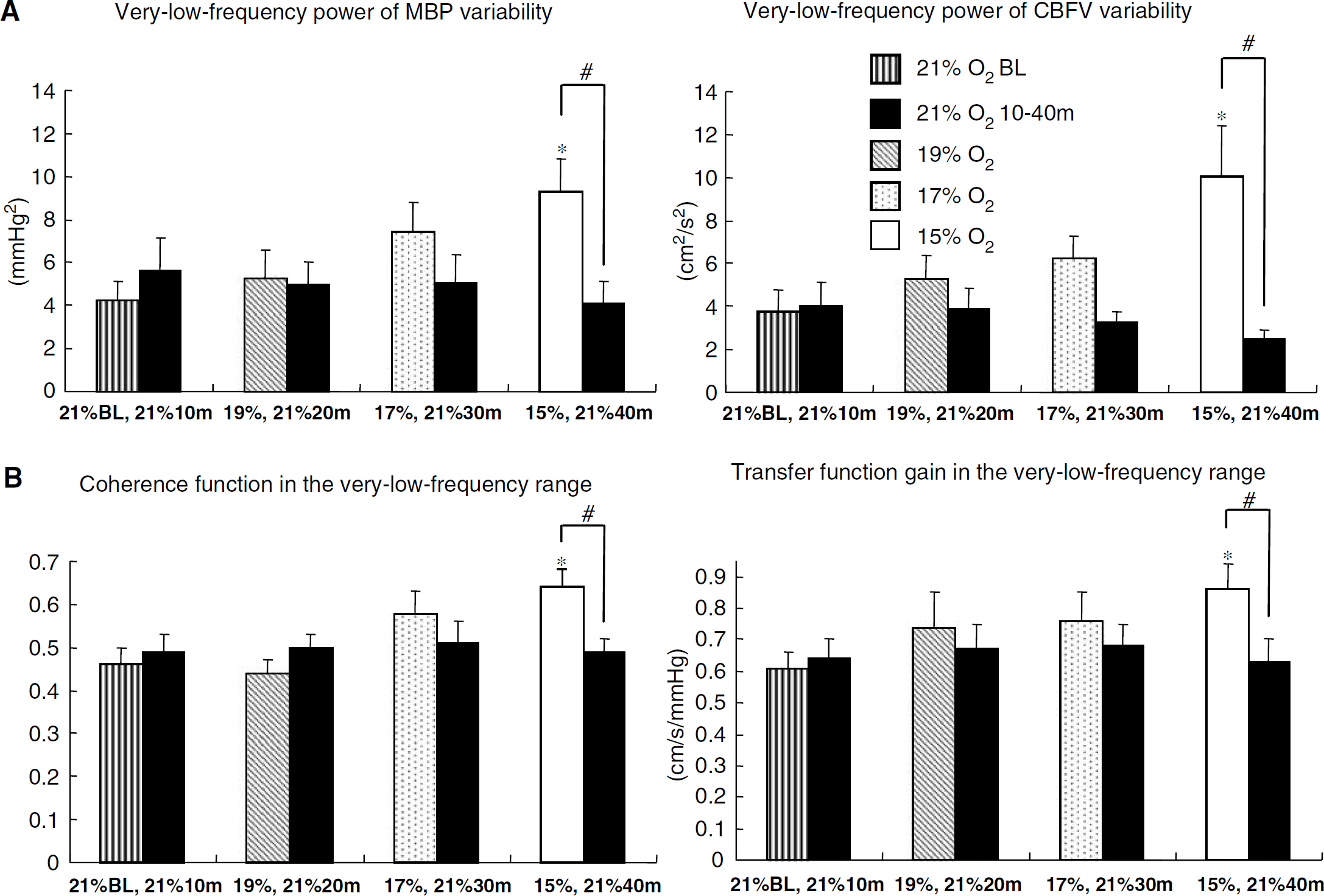
Mean changes (±s.e.m.) in frequency domain indices during exposure to hypoxia. (
Coherence and transfer function gain in the very-low-frequency range increased significantly by 53% ± 17% and 48% ± 21% with 15% O2, and these indices at 15% O2 were significantly higher by 38% ± 15% and 51% ± 20%, respectively, compared with those at 21% O2 for 40 mins in the time-control protocol (significant interaction effects, coherence, F3,42 = 3.347, P = 0.028; gain, F3,42 = 3.152, P = 0.035) (Figure 3B). However, transfer function gain and coherence in the low- and high-frequency ranges did not change. Phase did not change in any of the three ranges.
Single-Dose Protocol
All indices during the Single-dose protocol showed virtually the same trend as the results at 15% O2 in the Stepwise protocol (Tables 1–3). No significant interaction effect was noted with a comparison between the 15% O2 portion of the stepwise and single-dose protocols (e.g., CBF velocity variability at very low frequency, significant effect of stage, F1,8 = 11.298, P = 0.010; no significant interaction effect, F1,8 = 0.00445, P = 0.948).
Discussion
The present study found that acute normobaric hypoxia increases arterial pressure oscillations and CBF oscillations in the very-low-frequency range (0.02 to 0.07 Hz, 14 to 50 secs/cycle). Moreover, both coherence (strength of association) and transfer function gain (magnitude of transfer) between MBP and CBF velocity variabilities were increased in this frequency range, in which little linear dependence of CBF oscillations on ABP oscillations has been reported by numerous previous studies. Acute normobaric hypoxia therefore increases ABP oscillations and the dependence of CBF oscillations on ABP oscillations, resulting in apparent increases of CBF fluctuations in this frequency range. However, these changes were significant only with hypoxia at 15% O2, suggesting a possible threshold for such changes.
Transcranial Doppler ultrasonography was used for noninvasive and beat-to-beat estimation of CBF velocity in the present study and previous reports concerning cerebral autoregulation during hypoxia (Jansen et al, 2000; Levine et al, 1999; Van Osta et al, 2005). This approach is based on the assumption that diameter of the middle cerebral artery changes minimally (< 4.0%) during hypoxia, hypotension, or a variety of stimuli (Giller et al, 1993; Larsen et al, 1994; Poulin and Robbins, 1996; Serrador et al, 2000). Tonometry was used for non-invasive and beat-to-beat estimation of blood pressure. The accuracy of this device during hypotension has been confirmed by comparisons with tonometric and intra-arterial pressure monitors (Kemmotsu et al, 1991).
Diastolic blood pressure and MBP tended to decrease during exposure to hypoxia with 15% O2. Vasodilation by the direct effect of hypoxia on systemic vessels (Messina et al, 1992; Wadsworth, 1994) may not be completely counteracted by vasoconstriction after sympathetic activation in the systemic circulation during this level of hypoxia, resulting in a tendency toward decreasing ABP. However, 5 mins averages for CBF velocity were unchanged. According to the traditional concept of cerebral autoregulation, steady-state CBF remains relatively constant against changes in ABP via regulation of cerebrovascular resistance (Paulson et al, 1990). Despite a tendency toward decreases in blood pressure, 5 mins averages in CBF velocity remained unchanged in the present study, suggesting that static cerebral autoregulation may be maintained during acute exposure to hypoxia at 15% O2. The present result is consistent with previous studies reporting that mild hypoxia (effective altitude, 2400 or 3000 m) does not alter steady-state CBF (Buck et al, 1998; Otis et al, 1989). The important findings in the present study may be that mild hypoxia augmented fluctuations around the unchanged averages in CBF velocity, and increased in the maximum and decreased in the minimum of beat-to-beat CBF velocity.
For static cerebral autoregulation, adjustments in cerebral arteriolar beds maintain CBF relatively constant and independent from changes in ABP. However, it is not clear how changes in this static system translate or influence changes that ABP oscillations impose on CBF oscillations. The present study found that acute normobaric hypoxia increases both MBP and CBF velocity oscillations in the very-low-frequency range. In dynamic situations, although beat-to-beat CBF velocity fluctuates spontaneously in response to short-term oscillations in ABP, CBF velocity oscillations do not display linear dependence on slower oscillations in ABP, particularly at very low frequency (< 0.07 Hz, slower than every 14 secs), as indicated by low coherence function (Eames et al, 2004; Edwards et al, 2002; Kuo et al, 1998; Muller and Marziniak, 2005; Ogawa et al, 2006; Panerai et al, 1999, 2005; Schondorf et al, 2001; Zhang et al, 1998, 2002). In the present study, estimates of coherence function increased significantly to > 0.6 in the very-low-frequency range with 15% O2 in the stepwise protocol, indicating increased dependence of CBF oscillations on ABP oscillations. Moreover, transfer function gain increased significantly in this range with 15% O2, suggesting greater oscillations in CBF with any given oscillations in blood pressure. These changes of dynamic regulation induced by 15% O2 indicate that the influence of blood pressure oscillations on CBF fluctuations is increased for this frequency range.
The results of 5 mins exposure to 15% O2 in the Single-dose protocol showed virtually the same trend as the results for 30 mins exposure to 15% to 19% O2 in the Stepwise protocol. Although our inability to distinguish any significant interaction could be due to the small number of subjects (n = 9, single-dose protocol), no significant interaction effects were identified with a comparison between the 15% O2 portion of the Stepwise and Single-dose protocols. Thus, even a 5 mins exposure to 15% O2 seems to exert effects similar to a 30 mins stepwise exposure.
Potential mechanisms influencing static cerebral autoregulation include metabolic factors, myogenic factors, the intracranial nervous system, the autonomic nervous system, and endothelial factors (Paulson et al, 1990). Dynamic cerebral autoregulation may be more sensitive to changes in these factors. Dynamic cerebral autoregulation in the very-low-frequency range is partly modulated by the autonomic nervous system (Zhang et al, 2002). Stimulation of peripheral chemoreceptors by acute hypoxia alters autonomic nervous activities (Hughson et al, 1994; Saito et al, 1988). In fact, heart rate and MBP variability at low frequency as an index of sympathetic vasomotor activity increased during hypoxia with 15% O2 in the present study. One of the mechanisms of increases in dependence of CBF oscillations on blood pressure oscillations and magnitude of transfer during acute normobaric hypoxia would thus involve the alteration of autonomic nervous activities. Hypoxia also alters the release of endothelial factors and smooth muscle function. For example, acute hypoxia induces cerebral vasodilation, mainly through the release of NO (Van Mil et al, 2002). Changes in myogenic and/or endothelial processes may alter dynamic regulation of cerebral circulation. No clear conclusions can be drawn, since the present study did not attempt to elucidate the mechanisms underlying changes in dynamic cerebral autoregulation.
Arterial carbon dioxide exerts significant influences on steady-state CBF and CBF autoregulation (Giller et al, 1993; Zhang et al, 1998). However, although doubts may exist regarding the accuracy of end-tidal carbon dioxide measurements, no significant changes in ETCO2 or respiratory rate were noted in the present study. Even if hypoxia induces hyperventilation, lower arterial PCO2 may induce decreases (Muller et al, 2003) or no changes (Eames et al, 2004; Edwards et al, 2002) in coherence and transfer function gain, and increases of phase in the very-low-frequency range (Birch et al, 1995). Changes in respiration thus may not represent the cause of the present results in the very-low-frequency range. However, even undetected slight hyperventilation may alter respiratory blood pressure oscillation or baroreflex regulation at low- and high-frequency (Eames et al, 2004; Frederiks et al, 2000), so the results, particularly at high frequency, might then be partially or totally achievable by slight changes in respiratory patterns rather than by low arterial PO2. The lack of changes in ETCO2 and respiratory rate during hypoxia in the present study may be related to the duration of exposure and levels of hypoxia, which may have been too short and/or mild to cause significant effects of hyperventilation, particularly since group-averaged SaO2 did not decrease to <93%. The present results for ETCO2 are consistent with a previous study showing minor changes in arterial PCO2 during acute exposure to an effective altitude of 3000 m (Buck et al, 1998).
Only two previous studies have investigated the effects of altitude on dynamic cerebral autoregulation in humans, and the findings of these reports were contradictory. Levine et al (1999) reported that dynamic cerebral autoregulation was altered by chronic severe hypoxia. However, Van Osta et al (2005) reported that dynamic cerebral autoregulation did not differ between sea level and an altitude of 4559 m, although dynamic cerebral autoregulation was found to be impaired in subjects with a high acute mountain sickness score. At first glance, the present finding of altered dynamic cerebral autoregulation at mild hypoxia (15% O2) was considered inconsistent with the results of Van Osta et al, however, these discrepancies may be explained by differences in analysis techniques estimating dynamic autoregulation. Van Osta et al estimated dynamic autoregulation using an index calculated from approximately 5 secs of CBF restoration to baseline. The time scale of estimates for dynamic autoregulation was similar to that of transfer function indices in the low- and high-frequency ranges in the present study. Their result is thus consistent with our absence of changes in transfer function indices for low- and high-frequency ranges. Another possible explanation presents itself. Adapted humans could display a dynamic regulation differing from that of non-adapted humans, but which might still be considered normal. A recent high-altitude study showed that adapted individuals have lower cerebrovascular responses to hypoxia at high altitude compared with unadapted controls at sea level (Norcliffe et al, 2005). Future studies using transfer function analysis on adapted individuals or chronic exposure to hypoxia may be necessary to reconcile this dispute.
Acute mountain sickness characterized by headache is a syndrome seen in humans exposed to high altitudes, and the pathophysiological mechanisms remain unclear. Acute mountain sickness may represent an early stage of high-altitude cerebral edema, and could be caused by impaired cerebral autoregulation (Hackett and Roach, 2001). In the present study, changes in the indices of dynamic cerebral autoregulation were significant only for hypoxia with 15% O2 (effective altitude, about 2600 m). These significant changes in cerebral autoregulation above an effective altitude of 2600 m may be consistent with the observation that virtually no cases of altitude illness occur below 2000 m, with a significant increase in frequency above 2500 m (Hackett and Roach, 2001).
In light of the accumulated evidence regarding the interpretation of transfer function analysis between MBP and CBF velocity variabilities, as mentioned above, the present results may suggest that hypoxia can reduce the efficiency of dynamic cerebral autoregulation. Even if transfer function analysis is misleading when CBF velocity oscillations are modulated by other physiological variables (e.g., intrinsic cerebral vasomotion, CO2, etc.), the present results indicate that acute exposure to normobaric hypoxia markedly increases CBF instability in the very-low-frequency range. Moreover, significant changes occurred only with hypoxia at 15% O2, suggesting the presence of a threshold for these changes in the regulation of CBF in response to acute hypoxia.
Footnotes
Acknowledgements
The authors wish to thank Dr Sayaka Kita for her assistance with the acute hypoxia experiment. The authors would also like to thank Drs R Zhang and BD Levine for their support during the project and their comments on the manuscript.