
Abstract
Previous studies suggest that delayed neuronal death occurs in patients with inflicted traumatic brain injury (TBI) from child abuse. It is unknown whether the mode of this delayed neuronal death represents apoptosis or necrosis, a distinction that carries therapeutic ramifications. Cytochrome c, an electron transport chain component, can be released from mitochondria under conditions of cellular stress, whereupon it can initiate and serve as a biomarker of apoptosis. To resolve this issue, cytochrome c concentration was determined in 167 ventricular cerebrospinal fluid (CSF) samples from 67 infants and children with TBI (including 15 patients diagnosed with child abuse) by ELISA. Controls included lumbar CSF from 19 infants and children without trauma or meningitis. A multivariate model adjusted for multiple within-subject observations was used to identify clinical variables associated with CSF cytochrome c. Other apoptosis-related proteins were also examined in a subset of TBI patients. Increased CSF cytochrome c was independently associated with inflicted TBI (P = 0.0001) and female gender (P = 0.04), but not age, Glasgow coma scale score, or survival. Other apoptosis-related proteins including Fas and caspase-1 were increased in CSF after TBI, but did not independently discriminate between accidental and inflicted TBI. These data suggest that apoptosis, as detected by the presence of cytochrome c in CSF, is uniquely prominent among the subset of TBI patients diagnosed with child abuse. The degree of apoptosis after TBI also appears to be gender-dependent. Development of strategies targeting apoptosis after TBI, particularly in victims of child abuse and in girls, appears justified.
Introduction
Traumatic brain injury (TBI) initiates a variety of biological cascades in injured tissue, including apoptosis (also termed programmed cell death) and inflammation. Evidence for these processes can be readily detected in cerebrospinal fluid (CSF) (Clark et al, 2000; Kochanek et al, 2000). While any moderate to severe, blunt, or penetrating trauma can initiate these cascades, the multifaceted and complex biologic and social factors associated with inflicted injury from child abuse appear to yield a unique biochemical footprint in CSF. Factors may include (1) age of victim and developmental brain stage as child abuse victims are typically younger, (2) delayed presentation compared with ‘witnessed TBI’ such as seen in motor vehicle-related accidents, (3) physical mechanisms unique to child abuse such as violent shaking and/or blunt force injury, repeated insults, and the potential for secondary cerebral ischemia related to infants presenting in agonal states (Ruppel et al, 2002). For example, induction of the stress response (Lai et al, 2004), elevation of cytokines (Bell et al, 1997), excitotoxicity (Ruppel et al, 2001), and apoptosis (Clark et al, 2000), all appear more prevalent in inflicted TBI from child abuse compared with accidental TBI. Furthermore, child abuse victims represent a subset of TBI patients that have notoriously poor outcomes (Duhaime et al, 1998).
Previous studies suggested that delayed neuronal death occurs in infants with inflicted TBI from child abuse (Berger et al, 2002; Clark et al, 2000). While it is plausible that the mode of this delayed neuronal death represents apoptosis rather than necrosis, establishing whether this is indeed the case is important given the therapeutic ramifications. Delayed apoptotic neuronal death would conceivably allow a wider therapeutic window after TBI and may offer unique pharmacologic opportunities. Cytochrome c is an integral component of the electron transport chain that under conditions of cellular stress can be released from mitochondria where it has the capacity to initiate apoptosis (Zhivotovsky et al, 1998). Extracellular release of cytochrome c has recently been reported to be specific for apoptosis in humans (Renz et al, 2001) and sufficient to induce apoptosis in cell culture (Ahlemeyer et al, 2002). Thus, the presence of extramitochondrial cytochrome c serves as a biomarker of apoptosis. Accordingly, we examined CSF for cytochrome c in a large cohort of TBI patients that included victims of child abuse, and examined the relationship to inflicted versus noninflicted TBI and other relevant clinical variables. In a subset of these patients, biochemical markers affiliated with both apoptosis and inflammation were analyzed, including caspase-1 or interleukin-1β (IL-1/β) converting enzyme (ICE), pro-IL-1β, IL-1/β, the tumor necrosis factor (TNF) receptor family-related proteins Fas and Fas ligand (FasL), and caspase-3.
Materials and methods
Patients
Approval for this study was obtained from, and the requirement for informed consent for all clinical materials used in this study was waived by, the Children's Hospital of Pittsburgh Human Rights Committee. We evaluated CSF from a nonconsecutive series of 67 infants and children after severe TBI obtained over a period of 0 to 10 days after injury randomly selected from a TBI CSF tissue bank (Bell et al, 1997; Berger et al, 2002; Clark et al, 2000; Lai et al, 2004; Ruppel et al, 2001). All patients were admitted to the Pediatric Intensive Care Unit at the Children's Hospital of Pittsburgh. All patients had an extraventricular drain placed for intracranial pressure monitoring and CSF drainage as part of standard neurointensive care for our institution. Control CSF represented excess CSF obtained via lumbar puncture from infants and children who were found not to have meningitis based on cell count, culture, and Gram stain. Brain tissue was obtained from pediatric TBI patients undergoing decompressive craniectomy for uncontrolled intracranial hypertension. Control brain tissue represented brain-banked postmortem tissue from adult patients without CNS injury. All samples were stored at −70°C.
Clinical data collected included demographic information, mechanism of injury, Glasgow Coma Scale (GCS) score at presentation, and survival status. Inflicted TBI from child abuse was diagnosed by the Children's Hospital of Pittsburgh Child Protection Team, and was independent of enrolment into the study and intensive care unit management.
Cerebrospinal Fluid Collection and Enzyme-Linked Immunoabsorbant Assay
Cerebrospinal fluid was collected under sterile conditions from the closed extraventricular drain apparatus. Samples were centrifuged at 3,000 rpm for 10 mins to remove cellular debris and supernatants were decanted and stored at −70°C until batch analysis. ELISA kits for cytochrome c, Fas, FasL, caspase-1, pro-IL-1β, IL-1β, and caspase-3 were purchased from R&D Systems (Minneapolis, MN, USA), Bender MedSystems (Vienna, Austria), Oncogene Research Products (Cambridge, MA, USA), and Medical and Biological Laboratories LTD (Nagova, Japan). Optimal dilutions were determined and ELISAs were performed according to the manufacturer's instructions.
Western Blot Analysis
Western blot analysis for caspase-1 and caspase-3 were performed as previously described (Clark et al, 1999). Brain tissue was homogenized in lysis buffer containing 20 mmol/L HEPES (pH 7.8), 10 mmol/L NaCl, 1.5 mmol/L MgCl2, 1 mmol/L EDTA, 1 mmol/L EGTA, 1 mmol/L dithiothreitol, 1 mmol/L phenylmethylsulfonyl fluoride, and 2 μg/mL aprotinin. Aliquots were incubated at 4°C for 30 mins and centrifuged at 16,000g, and supernatants were boiled in standard loading buffer for 5 mins. Fifty-microgram protein samples were loaded on sodium dodecyl sulfate (SDS)-polyacrylamide gels, separated electrophoretically, and transferred to polyvinylidene difluoride membranes (Amersham, Arlington Heights, IL, USA) overnight. The transferred membranes were incubated for 1 h at room temperature in a polyclonal antibody that recognizes the 45 kDa intact and 10 kDa processed fragment of caspase-1 (1:500 dilution; Santa Cruz Biotechnologies, Santa Cruz, CA, USA), or a monoclonal antibody that recognizes the 32 kDa intact and 12 and 17 kDa processed fragments of caspase-3 (1:1000 dilution; Transduction Laboratories, Lexington, KY, USA). Membranes were then washed in phosphate-buffered saline (PBS) containing 0.1% Tween-20 and incubated in the appropriate secondary antibody (1:3000) for 1 h. The membranes were washed in PBS containing 0.1% Tween-20, incubated in chemiluminescence reagents (NEN Life Science Products, Boston, MA, USA), and exposed to X-ray film.
Statistical Analysis
All values are expressed as median [range] or mean ± standard deviation (SD) where appropriate. An indicator of high or low CSF biomarker levels between TBI patients and controls was calculated by determining the number of samples above and below the median value for controls, because control values for these biomarkers were unknown. Associations between high and low biomarker levels and the clinical variables age, gender, postinjury day, mechanism of injury, initial GCS score, and survival were determined using bivariate and multivariate logistic regression models that accounted for multiple data points for a given individual (intraclass correlation) as previously described (Clark et al, 2000; Lai et al, 2004). Age was examined as a dichotomous variable using age ≤4 versus >4 years based on previous studies from the National (USA) Traumatic Coma Databank showing differences in outcome after TBI in patients <4 years (Levin et al, 1992). Mechanism of injury was also examined as a dichotomous variable comparing noninflicted with inflicted TBI because inflicted TBI as a result of child abuse is known to have differences in the biochemical response to injury as well as outcome, when compared with noninflicted TBI (Clark et al, 2000; Lai et al, 2004). P<0.05 was considered significant.
Results
Patient Demographics
Demographics for control and TBI patients are shown in Table 1. Demographic data were available for 13 control patients. Patients in the TBI group were older and predominately male compared with the control group (P<0.05). This is consistent with the fact that in our control population most patients undergoing lumbar punctures as part of sepsis evaluations were infants, and therefore were younger than the TBI population. Demographic data for the accidental and inflicted TBI associated with child abuse subpopulations are also provided in Table 1. The subpopulation with inflicted TBI associated with child abuse, but not the accidental TBI subpopulation, has demographic characteristics similar to the control population. There was no difference between boys and girls TBI patients with respect to age (6.8 ± 4.6 versus 5.4 ± 5.0 years, respectively).
Patient demographics
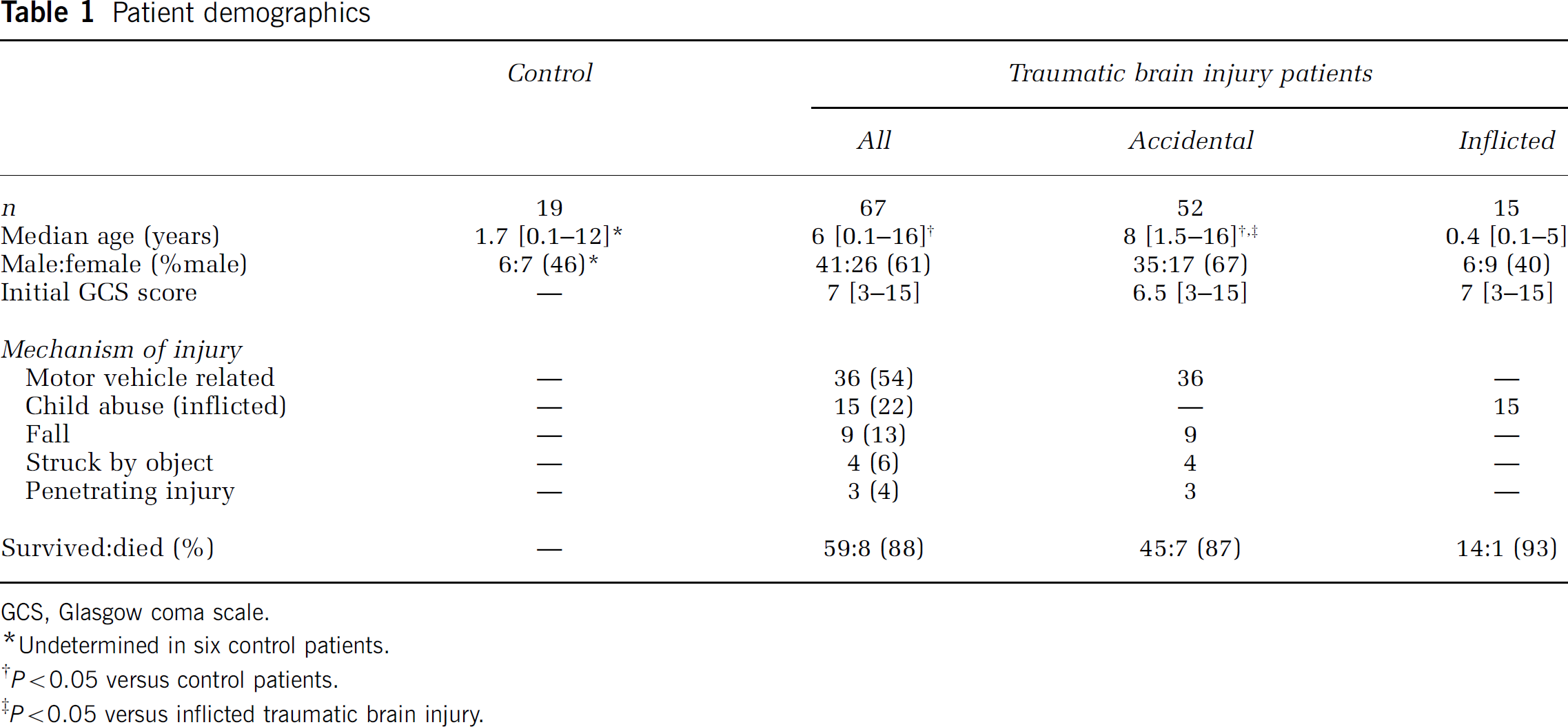
GCS, Glasgow coma scale.
Undetermined in six control patients.
P<0.05 versus control patients.
P<0.05 versus inflicted traumatic brain injury.
Concentrations of each biomarker were first determined in cohorts of 6 to 15 control patients and 19 to 32 TBI patients from the CSF tissue bank. These data were then analyzed using bivariate and multivariate statistical models. Results of this analysis suggested that of the biomarkers measured, CSF cytochrome c was the most sensitive in terms of predicting TBI patients who were victims of child abuse. Therefore, sample sizes were expanded for additional determination of CSF cytochrome c concentration, to verify the relationship between CSF cytochrome c as a biomarker of apoptosis and inflicted TBI from child abuse.
Cerebrospinal Fluid Cytochrome c
Cerebrospinal fluid cytochrome c was not different in all TBI patients compared with control patients (P>0.05; Table 2), nor were temporal changes detected, although higher CSF cytochrome c levels were seen at later time points after trauma (P> 0.05; Figure 1A). However, CSF cytochrome c was increased in patients with inflicted TBI from child abuse versus noninflicted TBI (2.10 [0.07 to 63.82] versus 0.62 [0.30 to 26.22] ng/mL, respectively), female versus male gender after TBI (1.63 [0.30 to 63.82] versus 0.61 [0.07 to 26.22] ng/mL, respectively), and GCS score on presentation ≥5 versus 3 or 4 (1.51 [0.07 to 21.59] versus 0.67 [0.30 to 63.82] ng/mL, respectively) by bivariate analysis (all P<0.05; Tables 2 and 3). Increased CSF cytochrome c was independently associated with inflicted TBI from child abuse (P<0.0001, β = 2.72; Table 3) and female gender (P<0.04, β = 1.14; Table 3) by multivariate analysis.
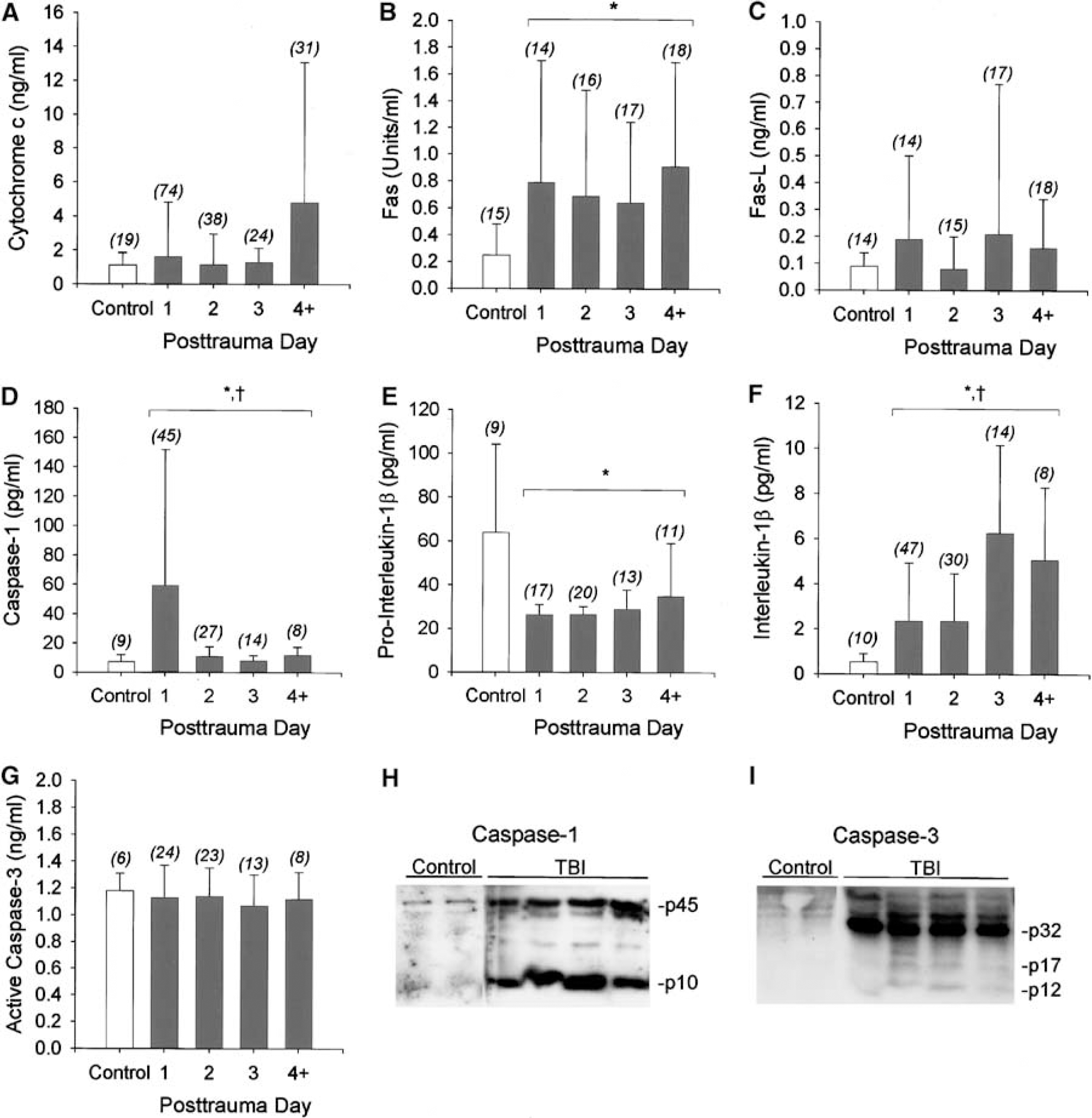
Temporal profiles of cytochrome c (
Cerebrospinal flid apoptosis- and inflammation-related protein concentrations by patient group
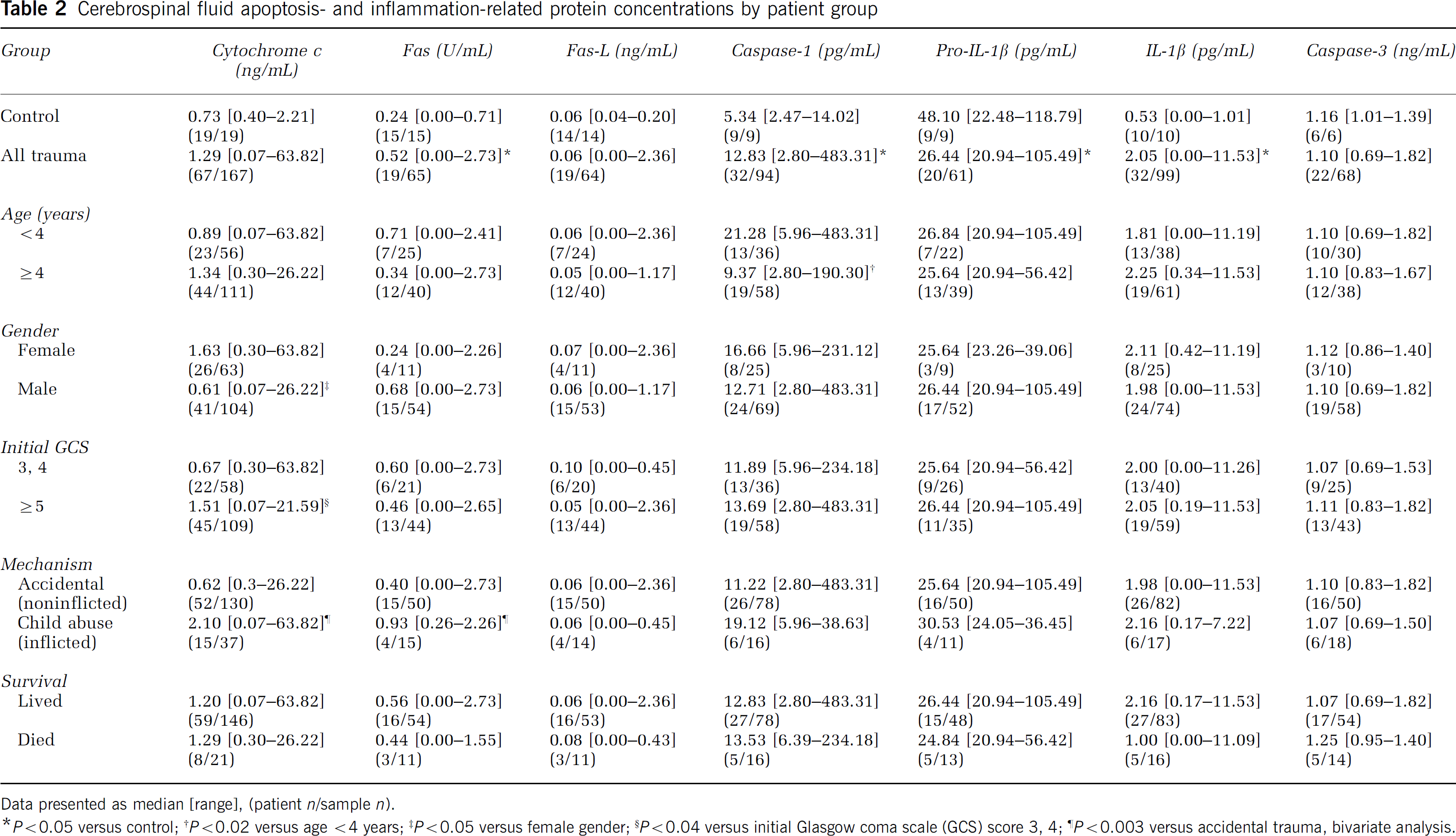
Data presented as median [range], (patient n/sample n).
P<0.05 versus control;
P<0.02 versus age <4 years;
P<0.05 versus female gender;
P<0.04 versus initial Glasgow coma scale (GCS) score 3, 4;
P<0.003 versus accidental trauma, bivariate analysis.
Logistic regression analysis for selected CSF biomarkers
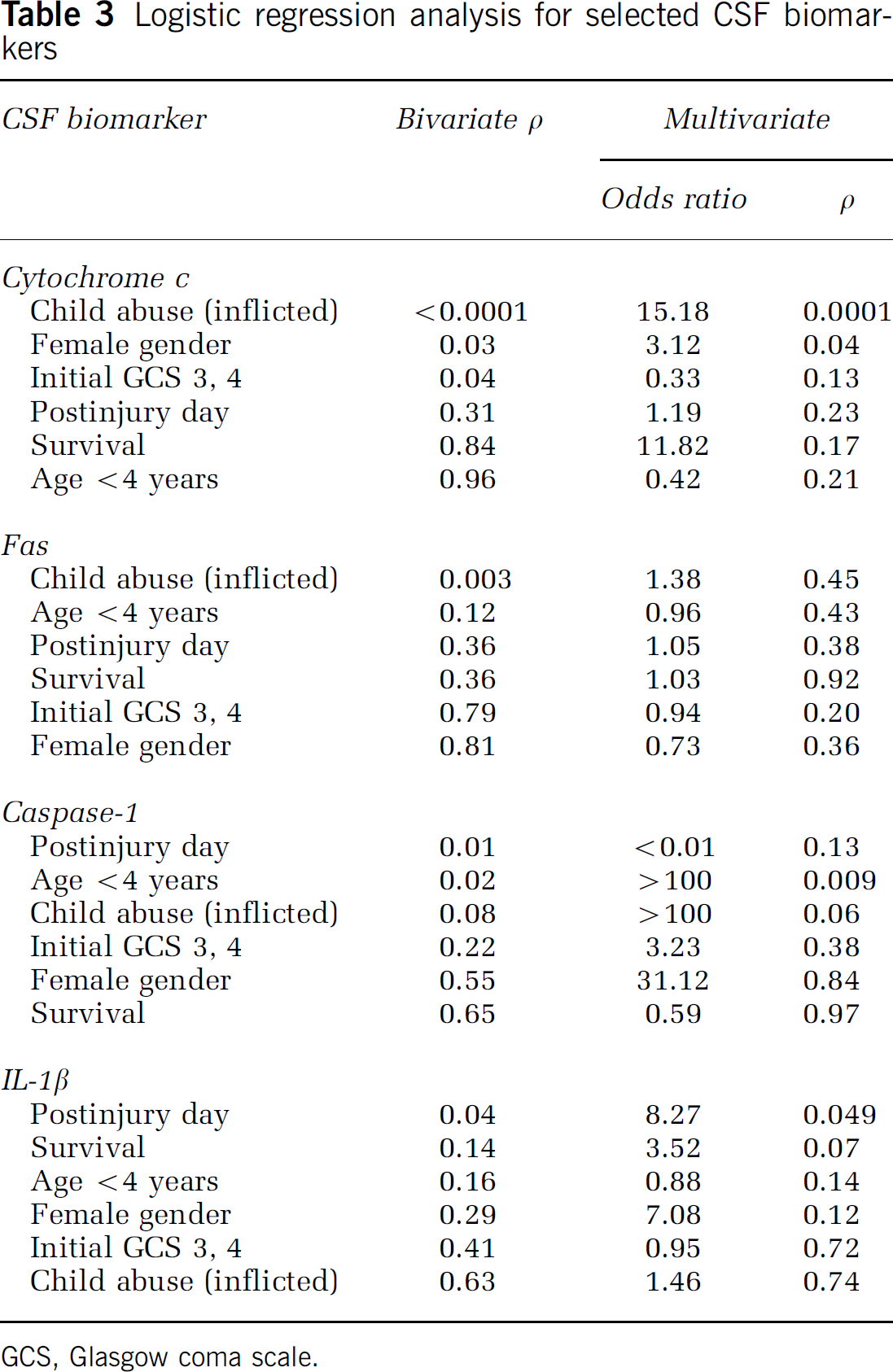
GCS, Glasgow coma scale.
Other Biomarkers of Apoptosis and Inflammation
Cerebrospinal fluid levels of Fas, caspase-1, and IL-1β were increased in all TBI patients compared with control patients (P<0.05; Tables 2 and 3, Figure 1). Pro-IL-1β, the precursor to IL-1β, was decreased in all TBI patients compared with control patients (P<0.05; Tables 2 and 3, Figure 1). CSF caspase-1 was increased on day 1 after TBI, whereas CSF IL-1β was increased after day 2 (P<0.05; Table 3, Figure 1). The findings that caspase-1 and IL-1β are increased, whereas pro-IL-1β was reduced, in CSF from TBI patients are biologically consistent with proteolytic processing of pro-IL-1β to mature IL-1β by caspase-1. Fas-L and active caspase-3 levels in CSF were not different between TBI and control patients.
Cerebrospinal fluid Fas was increased in patients with inflicted TBI from child abuse versus noninflicted TBI (0.93 [0.26 to 2.26] versus 0.40 [0.0 to 2.73] U/mL, respectively; P<0.003; Table 2). Cerebrospinal fluid caspase-1 was increased in TBI patients <4 years old (y.o.) versus ≥4 y.o. (21.28 [5.96 to 483.31] versus 9.37 [2.80 to 190.30] pg/mL, respectively; P<0.02; Table 2). Increased CSF caspase-1 was independently associated with age <4 years (P<0.01, β = 5.89; Table 3) by multivariate analysis. Increased CSF IL-1β was independently associated with postinjury day (P<0.05, β = −2.11; Table 3) by multivariate analysis. Associations between Fas-L, pro-IL-1β, and caspase-3 and any of the clinical variables included in the logistic regression models were not detected.
Caspase-1 and Caspase-3 in Injured Brain Tissue
We expected active caspase-3 levels to be increased in CSF from TBI versus control patients, or within subpopulations of TBI patients, given that cytochrome c is upstream of caspase-3 activation in the apoptotic cascade (Liou et al, 2003). One possible explanation for this is that caspase-3 levels are altered and/or caspase-3 is activated after TBI; however, caspase-3 is not released into CSF. To determine whether or not this might be the case, we examined brain tissue samples resected from a limited number of pediatric patients who underwent decompressive craniectomy for refractory intracranial hypertension after TBI. These samples were compared with adult brain-banked control samples from patients without CNS trauma, since pediatric brain-bank tissue was not available. Western blot analysis showed that the intact proform of caspase-3 was increased in pediatric TBI patients compared with adult control patients (Figure 1I), suggesting that either caspase-3 is developmentally regulated or increased after TBI, or both. Twelve and 17 kDa bands were also detected in some patients, suggesting caspase-3 activation and consistent with the possibility that active caspase-3 is not present within the CSF. Western blot was also performed for caspase-1. An increase in both intact and processed caspase-1 was seen in the injured brain from TBI patients versus controls (Figure 1H).
Discussion
The novel findings of this study can be summarized as follows. First, increased CSF cytochrome c was independently associated with the subpopulation of pediatric TBI patients with child abuse, suggesting that the delayed neuronal death observed in previous studies represents apoptosis. There was also an independent association between CSF cytochrome c and female gender. Second, Fas is increased in CSF from infants and children after TBI. Third, caspase-1 is increased in CSF from infants and children after TBI and is independently associated with younger age. Concurrent reductions in pro-IL-1β and increases in mature IL-1β, and examination of injured brain tissue, suggests that caspase-1 is activated after TBI. Fourth, while alterations in active caspase-3 were not detected in CSF after TBI, examination of injured brain tissue indicates that caspase-3 is increased, but not bioavailable in CSF.
Neuronal and other brain cell death after TBI is thought to be comprised of primary cell death, caused by mechanical destruction, and secondary or delayed cell death, caused by several pathologic cascades initiated by trauma (Kochanek et al, 2000). In humans, these cascades include ischemia (Bouma et al, 1992), excitotoxicity (Ruppel et al, 2001), and inflammation (Bell et al, 1997), each of which can trigger apoptosis (Clark et al, 1999, 2000). Apoptosis is a tightly regulated, homeostatic process important in embryologic development and in tissues with cell turnover; however, dysregulation of apoptosis can contribute to many human disease processes (Thompson, 1995). Reactivation of programmed cell death cascades in neurons culminating in apoptotic cell death has been reported in pediatric patients (Clark et al, 2000) as well as adults (Clark et al, 1999; Zhang et al, 2003) after TBI. Apoptosis can be triggered by cell surface death receptor-ligand interactions, for example, Fas and Fas-L, or by mitochondrial perturbations, for example, release of cytochrome c or apoptosis-inducing factor, referred to as extrinsic and intrinsic pathways, respectively (Liou et al, 2003). Perpetuation of apoptosis occurs via activation of initiator caspases, such as caspase-1, −8, and −9, and is executed by effector caspases, such as caspase-3 (Liou et al, 2003).
Our data suggest that apoptotic cell death, as defined by increased CSF cytochrome c, is prominent in patients with inflicted TBI from child abuse. Apoptosis could be all or in part responsible for the prominent delayed neuronal death seen in child abuse victims with TBI (Berger et al, 2002), and exaggerated apoptotic cell death in this population could help explain the notoriously poor outcomes reported (Duhaime et al, 1998). Our data have important clinical implications, given that several therapies are being developed whose actions include prevention of cytochrome c release from mitochondria, including the immunophilins cyclosporine A and FK506 (Snyder et al, 1998). Thus, it is possible that agents targeting cytochrome c release might be uniquely effective treatments for delayed neuronal death after inflicted TBI from child abuse, and that CSF cytochrome c might be an effective means of therapeutic drug monitoring. Serum cytochrome c has been used to monitor the effectiveness of cancer chemotherapy (Renz et al, 2001).
The association between CSF cytochrome c elevation after TBI and female gender might also be of clinical significance. The influence of gender on outcome and disease prevalence is well realized in adults, believed to be related predominately to effects of circulating sex steroids (Behl, 2002). However, recent experimental data suggest that innate gender differences exist at the neuronal level, independent of circulating sex steroids (Du et al, 2004). Consistent with the results of the present study, cytochrome c release and related apoptotic cell death is more prominent in neurons from female compared with male rats (Du et al, 2004). In clinical TBI, boys are felt to have less favorable outcomes after TBI compared with girls, although this is somewhat controversial (Donders and Hoffman, 2002; Morrison et al, 2004). Our data support a gender-stratified approach to treatment of pediatric TBI, particularly when targeting mechanisms involving cytochrome c.
The extrinsic pathway of apoptosis is initiated by the coupling of death receptors of the TNF family and their respective ligands, culminating in the formation of a death-inducing signaling complex (Qiu et al, 2002). Soluble Fas is increased in CSF from adult patients after TBI, with levels (0.06 to 4.33 U/mL) comparable to those seen in our study in pediatric patients (Lenzlinger et al, 2002). We have previously reported that Fas, but not Fas-L, is increased in injured brain tissue resected from adults after TBI (Zhang et al, 2003). A similar pattern of increased Fas but not Fas-L was seen in CSF from pediatric patients in the present study. While CSF Fas was associated with child abuse using bivariate analysis, it was not independently associated with child abuse after multivariate analysis.
Caspase-1, found to be homologous to ICE, converts pro-IL-1β to mature IL-1β, and plays a role in apoptosis independent of IL-1β (Tatsuta et al, 2000). Inhibition of caspase-1 reduces tissue damage after experimental TBI in mice (Fink et al, 1999). We have reported previously that caspase-1 is increased and cleaved in injured brain tissue resected from adults after TBI (Clark et al, 1999). A similar pattern was seen in brain tissue from pediatric patients in the present study. In addition, CSF caspase-1 was independently associated with younger age. Although we are unaware of data directly examining the effect of age on caspase-1 activation after acute brain injury, apoptosis is more robust in the developing brain (Yakovlev et al, 2001). Mature IL-1β is increased in CSF from adult patients after TBI; however, levels (20.6 ± 2.4 pg/mL) were ~10-fold higher than those seen in our study in pediatric patients (Marion et al, 1997). Regardless, the presence of cleaved caspase-1 along with concomitant increases in IL-1β and reductions in pro-IL-1β in CSF strongly suggest that caspase-1 is activated in pediatric patients after TBI, and support further preclinical testing of caspase-1 inhibitors, particularly in age-appropriate TBI models.
In contrast to caspase-1, caspase-3 is believed to function primarily in apoptosis with little or no direct effect on inflammatory processes. Indeed, caspase-3 activation is thought to be a key committed step in the apoptotic cascade (Liou et al, 2003). Caspase-3 mediates apoptosis after experimental TBI, and pharmacologic inhibition of caspase-3 results in improved outcome (Clark et al, 2000; Yakovlev et al, 1997). We expected active caspase-3 levels to be increased in CSF from TBI versus control patients, or within subpopulations of TBI patients (Liou et al, 2003). Similar to our previous report in adults after TBI (Clark et al, 1999), caspase-3 was increased and cleaved in injured brain tissue resected from pediatric patients, suggesting that caspase-3 is increased and activated after TBI; however, caspase-3 is not released into CSF. Increased caspase-3 activity in CSF using direct measurement, rather than an ELISA to detect processed/activated fragments, has been reported in adults after TBI (Harter et al, 2001). Because injured brain tissue samples were from pediatric patients, whereas control samples were from adult patients, it is possible that increased caspase-3 density seen on Western blot was related in part to developmental differences, more so than TBI, although it is clear that caspase-3 levels can be further increased by injury in the developing brain (Blomgren et al, 2001; Yakovlev et al, 2001).
While several novel observations were made in this study using CSF and brain tissue to investigate pathologic mechanisms operative in infants and children after TBI, limitations do exist in regard to the use of these human samples. Most notable is the difficulty in obtaining matched controls. Here CSF, control samples were obtained from infants primarily, as well as older children undergoing lumbar puncture. Thus, there are age discrepancies between the control and TBI subject populations, although the control and inflicted TBI from child abuse populations are age-matched. There are also differences in the site where CSF was obtained (lumbar versus ventricular). The insertion of a ventricular drain may produce local trauma in-and-of-itself, thereby influencing levels of inflammatory and apoptotic markers in CSF; however, this influence should be relatively minor relative to the original insult, and in terms of comparisons between TBI subgroups, should be of similar magnitude. For the brain, no pediatric control tissue was available, and therefore adult postmortem brain-banked controls were used. In defense of this strategy, the best available controls were used, and these methodologies have yielded important mechanistic insights into the pathobiology of human TBI (Kochanek et al, 2000).
This study attempts to elucidate the relative importance of apoptosis as a putative cell death pathway in pediatric TBI, and identify clinical features that may provide insight into potential target patient populations. Taken together, our data show that apoptosis associated with cytochrome c release is a prominent feature of TBI in victims of child abuse, and suggest that therapies targeting apoptosis in victims of inflicted TBI from child abuse should be explored, and that monitoring of CSF cytochrome c should be used to evaluate treatment efficacy.
Footnotes
Acknowledgements
The authors are grateful to Keri L Janesko for expert technical assistance, maintenance of the CSF tissue bank, and performance of many of the ELISAs.