
Abstract
An aging rat model of chronic brain hypoperfusion (CBH) that mimics human mild cognitive impairment (MCI) was used to examine the role of nitric oxide synthase (NOS) isoforms on spatial memory function. Rats with CBH underwent bilateral common carotid artery occlusion (2-vessel occlusion (2-VO)) for either 26 or 8 weeks and were compared with nonoccluded sham controls (S-VO). The neuronal and endothelial (nNOS/eNOS) constitutive inhibitor nitro-
Introduction
Vascular nitric oxide (NO) derived from endothelial nitric oxide synthase (eNOS) plays a key role in the regulation of microvessel tone and reactivity and is an important mediator of blood flow function (Moncada and Higgs, 1993; Huang et al, 1994; Iadecola, 1997; Maxwell, 2002). Release of vascular NO from cerebral endothelial cells produces vasodilation (Huang et al, 1994, 1996; Ignarro, 2002), while NO production from eNOS, inhibits platelet and leukocyte adhesion to the endothelium (Schini-Kerth, 1999), a process that may downregulate pro-inflammatory events (Kubes and Granger, 1992; Kubes et al, 1993).
There is now evidence that eNOS production in the presence of focal or global cerebral ischemia is an important factor in the pathogenesis of neuronal ischemic injury (Iadecola and Ross, 1997; Kader and Frazzini, 1993). During the early stages of cerebral ischemia, eNOS and neuronal NOS (nNOS) are upregulated in injured neurons producing large amounts of NO, but during necrosis NOS-containing neurons are lost and nNOS activity decreases, while the vascular effects of eNOS are no longer neuroprotective (Iadecola and Ross, 1997; Iadecola, 1997; Zhang et al, 1994).
Virtually nothing is known about the role vascular NO plays during neuronal dysfunction that results in spatial memory impairment after chronic brain hypoperfusion (CBH), a hemodynamic disturbance that is present before, during and after the early development of Alzheimer's disease (AD) (de la Torre and Mussivand, 1993; de la Torre, 2002a, 2002b, 2004; Johnson and Albert, 2000; Cabranes et al, 2004). What seems clear is that vascular NO is found in very high amounts in hippocampal pyramidal neurons, one of the principal and primary sites of AD pathology (Dinerman et al, 1994; Doyle and Slater, 1997; Kalaria, 2000).
This finding is relevant in understanding the physiopathology of AD because there is now considerable evidence that this dementia may develop from cerebrovascular events that lead to AD neurodegeneration (Breteler, 2000; de la Torre, 2000, 2002a, 2002b, 2004; de la Torre and Mussivand, 1993; Kivipelto et al, 2001; Meyer et al, 2000; Roher et al, 2003; Skoog, 1999; Vermeer et al, 2003; Iadecola, 2004).
In addition to the direct effects of CBH on energy metabolic abnormalities (de la Torre, 1997, 1999; Abdollahian et al, 1998), we have proposed that vascular NO is involved in the hemodynamic, humoral and inflammatory signals to which it is constantly exposed by circulating blood cells that have been subjected to CBH (de la Torre and Stefano, 2000) and that such signals can disrupt spatial memory via NOS pathways (de la Torre et al, 2003).
In the present study, we questioned whether pharmacologic manipulation of the three NOS isoforms, nNOS, inducible (iNOS) and eNOS, using selective and relatively selective inhibitors had an adverse effect on spatial memory function in aging rats after short-term CBH for 8 weeks. Preceding this study, we also examined the effect of nitro-
For the current study, we used a well-characterized aging rat model of CBH developed in our laboratory (de la Torre and Fortin, 1994; de la Torre et al, 1991, 1995, 1996) that mimics mild cognitive impairment (MCI) in humans (de la Torre et al, 1992, 2003). Aging rats subjected to CBH for less than 12 months develop spatial memory impairment but no other neurological deficits (de la Torre et al, 1991, 1997). Mild cognitive impairment has been defined as memory impairment that is abnormal for the individual's age and educational level but does not meet the criteria for dementia (Shah et al, 2000). It is estimated that more than 50% of subjects diagnosed with MCI convert to AD within 4 years (Hanninen et al, 2002; Morris et al, 2001), but that estimate might be greatly understated.
Our working hypothesis for this study was that given the role of vascular NO on cerebral blood flow regulation, disruption of its metabolism or reduction of its production should worsen spatial memory function in animals whose brain blood flow was already compromised after CBH.
The aim of this study was to examine the effect of CBH on spatial memory and to determine the role of NOS in this cognitive process using our rat model of MCI.
This question has therapeutic potential because the answer could provide evidence that the consequences of CBH lead to metabolic energy deficiency involving the endothelium and vascular NO before any spatial memory impairment. That being the case, targeting vascular NO or its synthetic pathway might be effective in the prevention or treatment of cognitive dysfunction.
Materials and methods
Chronic Brain Hypoperfusion Model
Male Harlan Sprague—Dawley rats (Harlan SD, Indianapolis, IN) 12 months old were used for the 6-month study and for the more acute 8-week study. In the 6-month study, rats were separated at random into two groups and assigned to receive CBH using 2-vessel occlusion (2-VO, N=22) or sham nonocclusion (SVO, N=21). Rats in the 2-VO group had the common carotid arteries doubly ligated for 6 months and rats in the S-VO group had carotid surgery but no occlusion.
In the 8-week study, rats were similarly subjected to 2-VO (N=50) and sham vessel occlusion (N=15) and at the end of 8 weeks, the 2-VO animals were further separated randomly into four groups to receive an NOS inhibitor or the vehicle. Previously, we have shown that mean arterial blood pressure (MABP), tympanic membrane temperature, hematocrit and arterial blood gases remain stable in the S-VO control and 2-VO animals for as long as 6 months after surgery (de Jong et al, 1999; de la Torre et al, 1995). Our previous studies and those of others have reported that two-vessel occlusion will result in a 22%–30% reduction of hippocampal blood flow that will stabilize after several weeks without further reduction (de la Torre et al, 1995, 1996; Tsuchiya et al, 1992).
All procedures conformed to the National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by the Case Ethics Committee for the humane treatment of animals.
Surgery
Surgery was performed under ketamine hydrochloride (100 mg/kg, IM) and methohexital sodium (50 mg/kg, IP) anesthesia. Atropine sulfate (0.1 mg IM) was administered to prevent respiratory distress. A ventral midline incision was made, and both common carotid arteries were gently separated from the carotid sheath and vagus nerve (de la Torre et al, 1992). Each common carotid artery was doubly ligated with 5-0 silk suture just below the internal—external carotid bifurcation (2-VO). S-VO nonoccluded control rats underwent similar surgery but no vessel ligation (S-VO). During surgery, temperature was maintained at 37°C with a rectal probe attached to a heating bulb. After surgery, rats were placed on a heating pad to maintain body temperature until full recovery. We have reported that the surgical procedure and recovery period do not modify serum electrolytes, pH, lactate, ATP, creatine phosphate or N-acetyl aspartate levels in brain (Abdollahian et al, 1998; de la Torre et al, 1991, 1992; de la Torre and Fortin, 1994).
Watermaze
Cognitive function was evaluated using the Morris watermaze test as we have described (de la Torre et al, 1992). This test has been shown to be to be sensitive to hippocampal function (Moser, 1995).
A 150-cm diameter pool in a room decorated with extramaze cues was divided into four quadrants (NW, SW, SE, and NE). A submerged platform (1.5 cm below the water surface) served as the escape platform. A working memory version of the Morris watermaze test was used, that is, the location of the escape platform was changed each day. The escape platform was placed near a perimeter platform and its location was chosen in a random fashion so as not to repeat its location over the 6 days of testing. Each rat was given 5 trials per day for 5 days before drug administration. On day 6, rats were administered a drug or vehicle, intraperitoneally, 30 mins before testing. Rats were allowed a maximum of 90 secs to find the platform.
If the rat was unable to find the platform in this time, it was guided to the platform and allowed a 30-sec rest on the platform. If the rat found the platform, the time it took (latency, in seconds) or distance travelled (in centimeters) was recorded.
Rats in the 6-month study were pretrained before surgery and thereafter every 2 weeks to allow the 2-VO and S-VO groups retain a similar learning response on the watermaze for 6 months when pharmacologic disruption of NOS production was attempted.
In the 8-week study, all rats were tested at 2 and 8 weeks post-surgery. Rats in the 8-week study were not pretested before surgery because previous experience has shown that a pretraining phase and repeated training after 2-VO results in much longer observation periods than 8 weeks before any spatial memory impairment is detected in this model (de la Torre et al, 1995). Testing at the 2-week observation period provided the baseline values for an early time period after 2-VO and retesting at the 8-week observation period provided a before and after evaluation of the effect of each NOS inhibitor on spatial memory performance.
After the final watermaze test, all rats were quickly rendered comatose by exposure to carbon dioxide and decapitated. For the 6-month study, one hemisphere was drop-fixed in paraformaldehyde for pre-embedding electron microscopic (EM) immunocytochemical detection of the eNOS isoform distribution and the other hemisphere served for light microscopic examination using Nissl staining.
Pharmacologic Treatments
In the 6-month study, 2-VO and S-VO control rats received 3 daily injections of the constitutive (nNOS and eNOS) inhibitor
In the 8-week study, 2-VO rats were further randomly separated into 4 groups at the end of 8 weeks to receive a single administration 1 h before watermaze testing of the following drugs:
Group 2-VO (vehicle only) 1 ml/kg peanut oil (N=14); Group 7-NI (7-nitroindazole) 25 mg/kg, a relatively selective inhibitor of nNOS (N=12); Group AG (aminoguanidine) 10 mg/kg, a selective inhibitor of iNOS (N=12); Group L-NIO ( Group S-VO control (surgery, no occlusion) was administered 1 ml/kg peanut oil (N=15).
The doses were chosen from previous trials to achieve temporary inhibition of each NOS isoform (Jiang et al, 2002; Wolff et al, 1998; Zhu et al, 2003; Escott et al, 1998; Moore et al, 1993).
L-NIO and aminoguanidine were dissolved in physiologic saline and 7-NI was dissolved in peanut oil. All drugs were injected intraperitoneally to give 1 ml/kg volume.
Preembedding EM Immunocytochemistry
Only rats subjected to 6 months of 2-VO were processed for routine transmission electron microscopy (TEM) and for preembedding immunogold techniques for eNOS as we have described (Aliev et al, 2002a, 2002b).
Vibratome sections were treated with hydrogen peroxide (3% in methanol) to block endogenous peroxidase activity for 30 mins, after which they were washed twice with Tris-buffered saline (TBS; 50 mmol/L Tris—HCl (pH 7.6), 150 mmol/L NaCl) for 10 mins at room temperature (RT), incubated in 10% normal goat serum (NGS) for 2 h at RT, and then incubated with a polyclonal antibody against eNOS (1:250) diluted in 1% NGS overnight at 4°C. After rinses in 10% NGS, gold-conjugated antibody to rabbit IgG (17 nm) was applied for 4 to 24 h and thoroughly rinsed in PBS. The sections were exposed to immunocytochemical detection of eNOS and routine TEM analysis, then post-fixed in 2.5% glutaraldehyde for 1 h and again rinsed with PBS. Finally, sections were exposed to osmium tetraoxide for 1 h at RT, rinsed, dehydrated through acetone and flat embedded in Spurr's embedding media. Randomly selected ultrathin sections were stained and viewed in a JEOL 100CX TEM at 80 kV.
Statistical Analyses
Multiple groups were compared using ANOVA and Scheffe's post hoc test. When comparing two groups, Student's t-test (unpaired variables, two-tailed) was used with Sigma Stat and Sigma Plot software (Jandel Scientific). All values were expressed as means±s.e.m. A probability value <0.05 was regarded as statistically significant.
Results
Six rats were lost to attrition in the 6-month and 8-week studies. Of the survivors, no neurologic deficits, loss of pupillary reflex or complications resulting from the surgical procedure were recorded during the observation period.
Figure 1 summarizes the results of
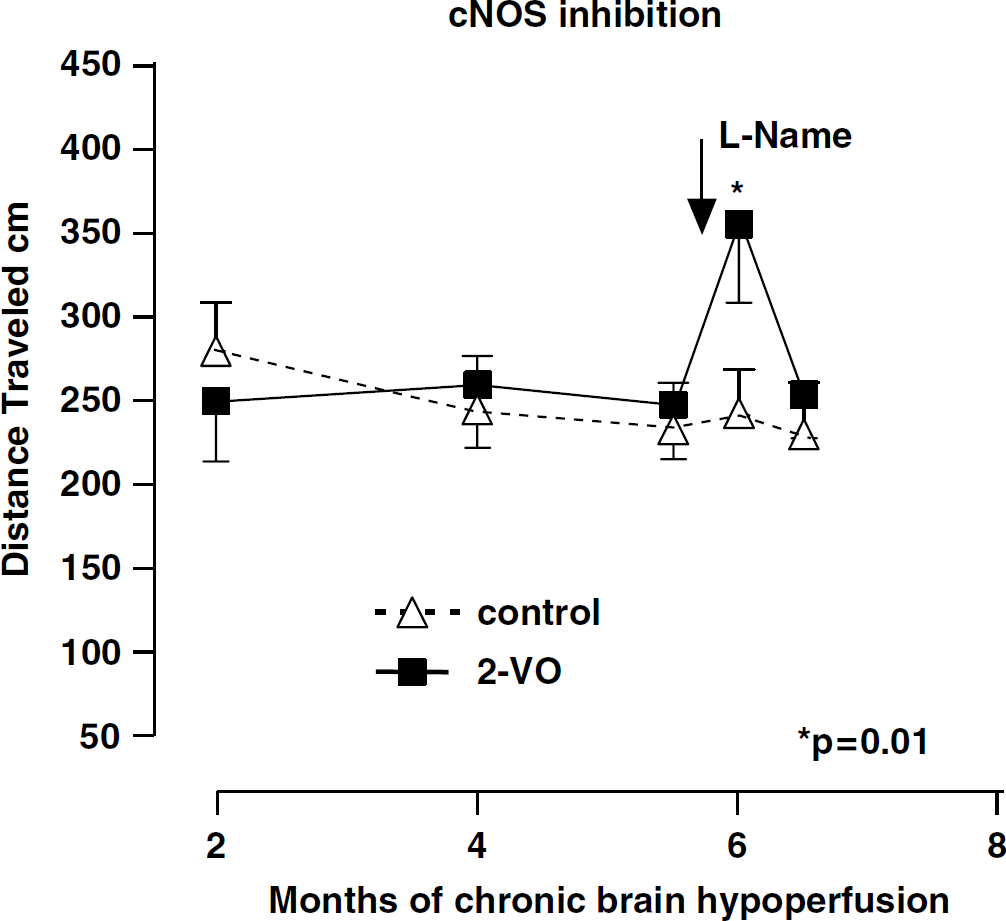
Spatial memory assessment in a modified Morris watermaze measuring mean distance travelled in rats subjected to two-vessel occlusion (-▪-, 2-VO) and control nonocclusion (-Δ-, control) for 2 to 6 months. At 6 months, total distance travelled increased significantly after 3 days daily intraperitoneal injections of 20 mg/kg of
After
Figures 2, 3 and 4 show the effects of the relatively selective inhibitors of nNOS, iNOS and eNOS on spatial memory function in rats subjected to CBH (Group 2-VO) or nonocclusion controls (Group S-VO) in the 8-week study when tested at the Morris watermaze at 2 and 8 weeks after 2-VO.
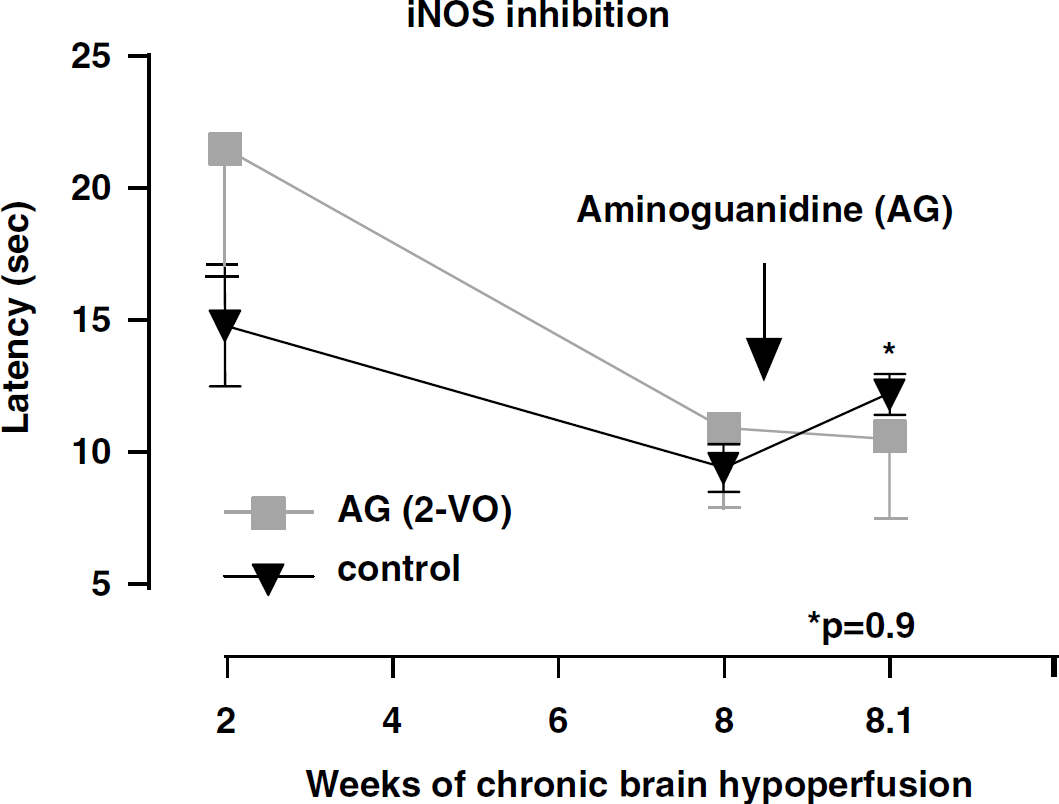
Spatial memory assessment in a modified Morris watermaze measuring escape latency (mean latency in sec) in rats subjected to 2-VO (-▪-) and control nonocclusion (-▾-, control) for 2 and 8 weeks. At 8 weeks, mean latency did not change significantly (P=0.9) after a single dose of 10 mg/kg AG, a selective inhibitor of iNOS, when compared with nonoccluded controls or pretreatment values.
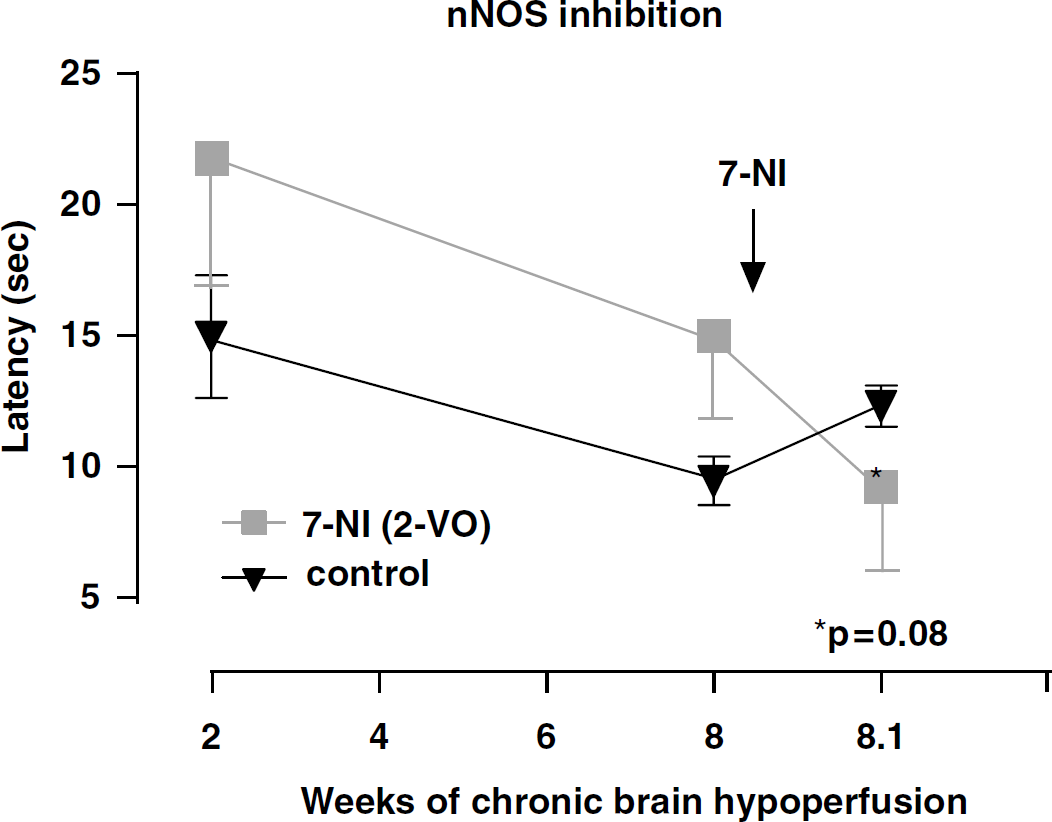
Spatial memory assessment (as above) measuring escape latency (mean latency in secs) in rats subjected to 2-VO (-▪-) and control nonocclusion (-▾-, control) for 2 and 8 weeks. At 8 weeks, mean latency showed a trend towards improvement, but did not reach significant level (P=0.08) after 25 mg/kg 7-NI, a selective inhibitor of nNOS, when compared with nonoccluded controls or pretreatment values.
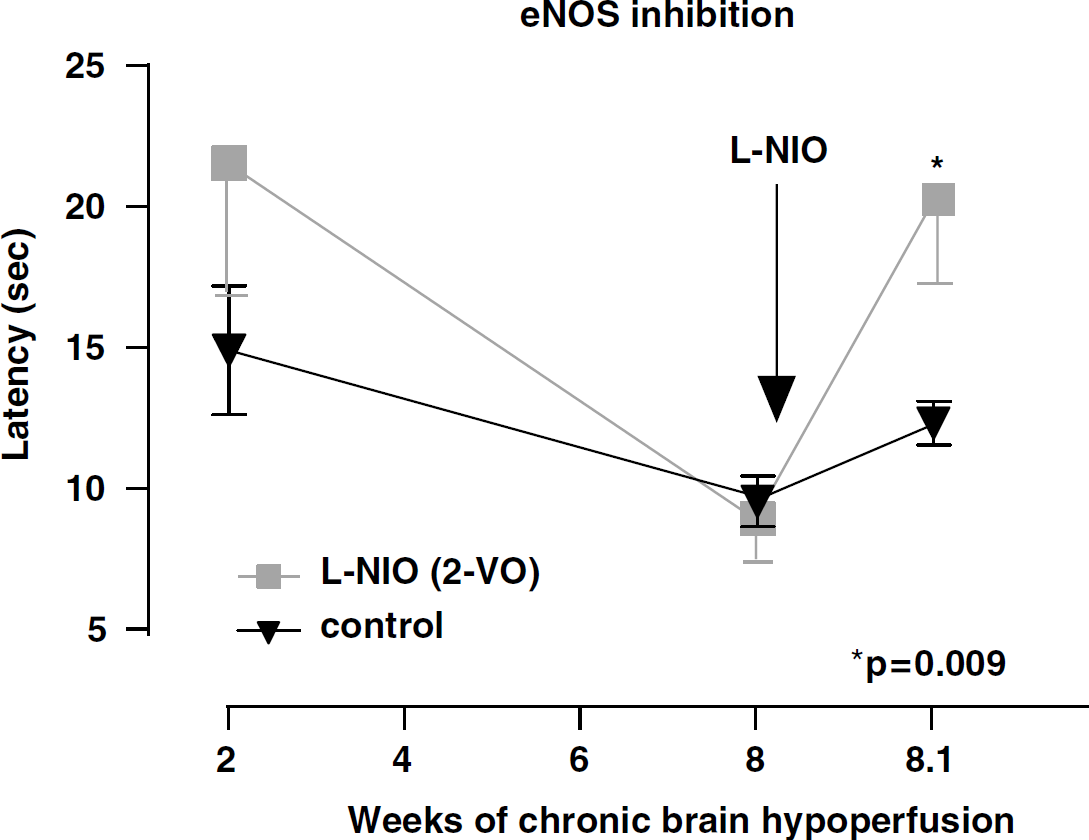
Spatial memory assessment (as above) measuring escape latency (mean latency in seconds) in rats subjected to 2-VO (-▪-) and control nonocclusion (-▾-, control) for 2 and 8 weeks. At 8 weeks, mean latency showed a significant deterioration (P=0.009) after 20 mg/kg L-NIO, a relatively selective inhibitor of eNOS, when compared with nonoccluded controls and pretreatment values.
At the 2-week testing period, and not unexpectedly, no differences between controls and 2-VO were seen. On week 8, 2-VO rats were randomly divided into 4 groups and retested on the watermaze together with nonocclusion controls. No significant spatial memory differences between groups were recorded at the 8-week time period. After administration of the nNOS (7-NI) and iNOS (AG) relatively selective inhibitors (Figures 2 and 3), a trend towards improved performance when compared with controls was observed only in the 7-NI-treated rats, but the difference did not reach statistical significance (P=0.08). By contrast, rats administered the relatively selective eNOS inhibitor
In the 8 week and 6 month study, light microscopic examination of the hippocampal region showed no morphologic differences between 2-VO and S-VO controls. In the 6 month study, ultrastructural examination of hippocampal CA1 capillaries in rats subjected to 2-VO revealed a smaller size EC containing damaged mitochondria characterized by transformation to lysosomal structures within the EC and in the perivascular area (Figure 5B). EC changes in 2-VO rats included a sharply contracted nucleus (Figure 5C) and condensed nuclear chromatin, and these abnormalities appeared to be associated with amyloid deposition found surrounding the capillary vessel wall (Figure 5D).
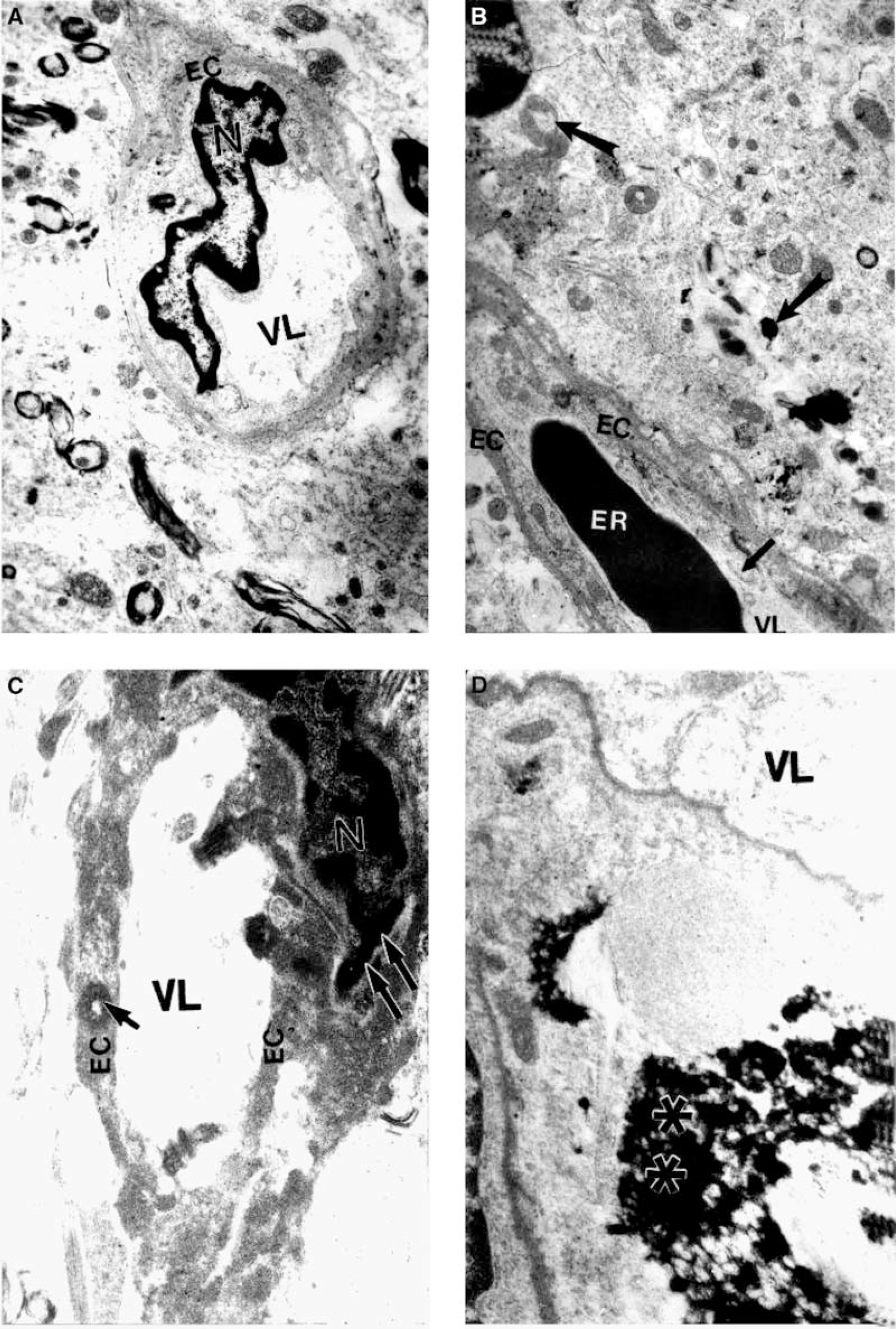
Ultrastructure of rat CA1 hippocampal capillaries from non-occluded control and rat subjected to 2-VO for 26 weeks. (
EM immunostaining in rats subjected to 2-VO for 6 months, showed almost an absence of eNOS-containing positive gold particles in the matrix of the vascular endothelium (Figure 6B). By contrast, increased density of eNOS-containing positive gold particles was observed in the cytoplasmic matrix of perivascular cells, together with electron-dense mitochondria indicative of a hypoxic insult (Figure 6B). Immunoreactive eNOS-containing positive gold particles were found markedly expressed in hippocampal neurons and in glial cells (Figures 6C and 6D). No significant changes were observed in the cellular and subcellular compartments of hippocampal capillaries, neurons, pericytes or glial cells in group S-VO, nonoccluded controls (Figures 5A and 6Aa) S-VO rats showed normal density of eNOS-containing positive gold particles in EC and normal-appearing mitochondria within the EC and in the perivascular area.
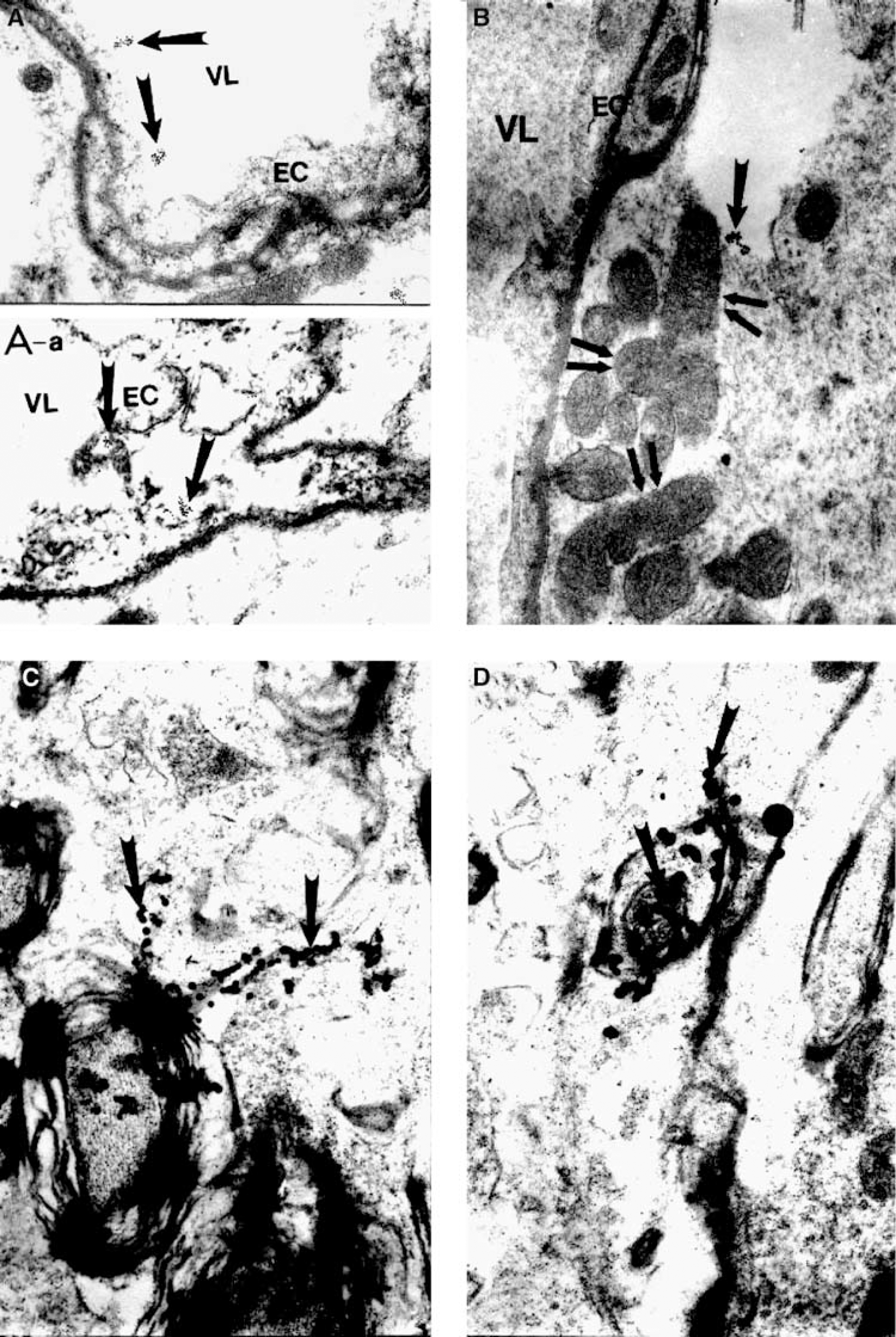
Ultrastructure of rat CA1 hippocampus showing eNOS immunoreactivity in nonoccluded control and rat subjected to 2-VO for 26 weeks. (
Discussion
The present findings support our working hypothesis that vascular NO is an important molecule in spatial memory function, at least in this CBH model. This conclusion is based on our present findings indicating that inhibition of eNOS using L-NIO significantly reduced spatial memory ability after 8 weeks of CBH. Inhibition of iNOS and nNOS isoforms using AG and 7-NI did not appear to exert the same negative effect on spatial memory in this study, but their relative importance on cognition after cerebral hypoperfusion needs to be further investigated.
Our results from the 6-month study suggest that repeated watermaze testing even in rats subjected to 2-VO is able to overcome the spatial memory impairment normally seen in these animals beginning shortly after 8 weeks of CBH. In the 26-week study,
Discriminant function analysis showed a correlation between CBH rats with microvessel abnormalities and poorer watermaze performance, and although the mean latencies for 2-VO and control groups were similar before
The marked depletion of eNOS-containing positive gold particles in the hippocampal EC of 2-VO rats and the overexpression of those eNOS particles in perivascular cells such as pericytes, glia and adjacent neurons do not yet invite a definitive explanation. Since normal laminar flow will convert to low, disturbed shear stress after CBH (de la Torre et al, 2003; de la Torre and Mussivand, 1993; de Nigris et al, 2003), eNOS expression within the EC is likely to be reduced or depleted over time. Disturbed flow has also been reported to increase the oxidative stress that can be induced by EC (Silacci et al, 2001) and may activate a variety of signal transduction events that in turn lead to further endothelial dysfunction (Gimbrone, 1999). However, overexpression of eNOS in neurons and glia (Figures 6C and 6D) may result from a hypoxic—ischemic process by CBH as a compensatory reaction to the eNOS depletion within the EC (Aliev et al, 2002c). It is of interest to note that the vascular tissue ultrastructural and immunocytochemical changes observed only in the 2-VO rats were evident in rats that developed significant spatial memory impairment even before
The implications of these findings need to be considered in light of the anatomic localization of vascular NO and its apparent key role in the regulation of vascular homeostasis (Iadecola et al, 1994, 1995; Schini-Kerth, 1999). Vascular NO is produced by the eNOS enzyme found mainly in endothelial cells, where it acts to prevent the inappropriate adhesion of leukocytes and platelets (Maxwell, 2002). However, when the physiologic action of vascular NO is compromised, as shown in our present study, vasospasm, platelet aggregation, smooth muscle cell proliferation and leukocyte adhesion may contribute to cerebrovascular pathology and cognitive loss because of reduced blood flow to the brain (Huang et al, 1996; Ignarro, 2002; Radomski et al, 1987; Moncada, 1999).
Our previous findings may help explain our present results. For example, our present results suggest that increased vascular NO production (from eNOS upregulation?) for the first 2 weeks might have prevented spatial memory impairment after 2-VO, but only temporarily. This mechanotransduction effect produced by low shear stress on the endothelium would increase vascular NO bioactivity as a response to reset normal homeostasis and diminish the damage caused by the cerebral hypoperfusion. Unable to achieve homeostasis, vascular NO levels ‘dive’ below baseline by 8 weeks of 2-VO and are only ‘marginably’ able to: (i) regulate normal vascular perfusion, (ii) block granulocyte adherence in blood vessels and (iii) prevent proinflammatory reactions intraneuronally (de la Torre and Stefano, 2000). With time, basal levels of vascular NO cannot maintain endothelial cell conformation resulting in endothelial cell distortions, a finding confirmed in the present study (Figures 5B and 5C). Blood flow dyshomeostasis affecting eNOS bioactivity is therefore likely to contribute to the spatial memory impairment already beginning but not fully expressed 8 weeks after 2-VO (de la Torre et al, 2003) and to long-term potentiation in hippocampal EC (Blackshaw et al, 2003). Moreover, pharmacologic inhibition of eNOS with L-NIO in the presence of mild but persistent CBH would worsen or accelerate spatial memory dysfunction as bioavailability of vascular NO is further depressed. This outcome may explain why rats administered L-NIO but not 7-NI or AG worsened their spatial memory ability.
Another interpretation of these findings is that L-NIO might have been toxic to vulnerable pyramidal neurons in CA1 and exerted a negative action on spatial memory by compromising this cell population. A neurotoxic effect is less likely, since it has been reported that L-NIO does not reduce the anoxic depolarization of CA1 neurons after cerebral ischemia (Jiang et al, 2002).
Despite the unknown signal transduction affecting the eNOS signaling complex, production of vascular NO appears to represent an important protective mechanism against cerebral ischemia (Dawson, 1994; Samdani et al, 1997; Lee, 2000; Veltkamp et al, 2002) and ostensibly also in CBH as shown by our present findings.
We conclude from our present data that brain endothelial cell function and release of NO play a critical role in spatial memory function, possibly by regulating microvessel tone and optimal cerebral blood flow. Vascular NO production could be a therapeutic target during CBH, a hemodynamic abnormality well known to exist during human MCI and continuing later in the development of progressive AD (de la Torre, 2002b; de la Torre et al, 1997, 2004; Johnson and Albert, 2000; Kalaria, 2000; Meyer et al, 2000).
Footnotes
Acknowledgments
We thank Mrs Iryna Vashchenko (Microscopy Research Center, Department of Pathology, CWRU) for excellent technical assistance.