
Abstract
CC chemokine receptor 2 (CCR2) plays important roles in extravasation and transmigration of monocytes under inflammatory conditions. CCR2 and its ligands have been extensively studied in a range of inflammatory diseases in the central nervous system (CNS), including multiple sclerosis, Alzheimer's disease and ischemic stroke. This brief review summarizes our current understanding of the physiologic and pathologic roles of CCR2, focusing on its involvement in CNS inflammatory diseases. There appears to be a rationale for exploring therapies involving CCR2 inhibition in multiple sclerosis and ischemic stroke, but there is also evidence for immunomodulatory and protective effects of CCR2 activity during CNS inflammation. The critical balance between protective and detrimental roles of CCR2-dependent recruitment of leukocytes must therefore be carefully examined to guide safe and effective development of any therapies involving CCR2 modulation.
INTRODUCTION
Inflammation is now recognized as an important contributor to central nervous system (CNS) injury and neurodegenerative disease. Immune cells such as neutrophils, monocytes, macrophages, dendritic cells, and lymphocytes have been shown to be involved in numerous CNS diseases, including multiple sclerosis, Alzheimer's disease, and ischemic stroke.1–4 Recruitment of leukocytes requires chemokines and activation of chemokine receptors. Selective targeting of chemokine receptors may thus allow manipulation of specific immune cell types entering the brain after CNS injury. Growing evidence suggests that bone-marrow-derived monocytes/macrophages are examples of immune cells that may potentially exacerbate, but in some instances protect against further brain injury after their recruitment. CC chemokine receptor 2 (CCR2) is the chemokine receptor, which regulates the mobilization of monocytes from bone marrow to the inflammatory sites, and it has been extensively studied in CNS inflammation. In this brief review, we describe the known roles of CCR2 under both physiologic and pathologic conditions, and outline evidence for its involvement in inflammation of the CNS as well as the potential benefits of CCR2 inhibition in certain diseases.
CCR2: A CHEMOKINE RECEPTOR THAT REGULATES IMMUNE CELL MOBILIZATION
Chemokine receptors are G-protein-coupled receptors with seven transmembrane domains that are activated by chemokines. Chemokines are a family of small proteins (8 to 10kDa) that play an important role in the immune system by regulating the migration of immune cells. There are four chemokine subfamilies (CC, CXC, CX3C, and XC) based on their structural and functional differences.5,6 CCR2 is a chemokine receptor involved in monocyte mobilization. It consists of seven transmembrane domains linked by intracellular and extracellular loops, an extracellular N-terminal that binds to ligands and is important for specificity; and an intracellular C-terminal that contributes to intracellular signaling.7,8
CHEMOKINE LIGANDS FOR CCR2
CCR2 is activated by several chemokines, including CCL2 (formerly known as MCP-1), CCL7 (MCP-3), CCL8 (MCP-2), CCL12 (MCP-5, mouse only), CCL13 (MCP-4), and CCL16 (HCC-4, human only). Activation of CCR2 results in directional migration of receptor-bearing cell types. CCL2 is the most potent activator of CCR2 signaling, leading to monocyte transmigration.9–11 CCL7, CCL8, and CCL13 are also agonists for CCR1.12–14 The multiple ligands for individual chemokine receptors could enable complex regulatory functions and a degree of redundancy within the chemokine system, so as to ensure effective mechanisms of immunologic defense.
EXPRESSION OF CCR2
CCR2 was first identified on monocytes, which constitutively express the receptor, and which is downregulated after differentiation into macrophages. 15 In mice, CCR2 is mainly expressed on a subset of monocytes that express high levels of Ly6C, but low levels of the CX3C-chemokine receptor 1 (CX3CR1). These are often referred to as either ‘Ly6Chi’ or ‘inflammatory’ monocytes, as a large number of Ly6Chi monocytes are recruited to inflamed tissue in a CCR2-dependent manner, and then produce Th1-type pro-inflammatory cytokines. 16 A small proportion (˜18%) of ‘Ly6Clo’ monocytes also express cell-surface CCR2. 17
CCR2 mRNA has been detected in T cells, immature B cells, natural killer cells, basophils, and dendritic cells. 18 Although these cell types do not express CCR2 protein constitutively, 17 a small population of natural killer cells, T cells, and basophils have been shown to express CCR2 protein under inflammatory conditions.19–21 Furthermore, CCR2 is also expressed on endothelial cells during inflammation, consistent with a potential role in wound repair. 22
Information regarding the expression of CCR2 on non-hematopoietic cells in the CNS is currently limited and inconsistent. It is reported that CCR2 has been detected by immunohistochemistry in neurons, endothelial cells, astrocytes and microglia in various regions of human brain, with its highest expression in hippo-campus. 23 Similarly, an immunohistochemical study of rat brain observed neuronal expression of CCR2 in various regions such as cerebral cortex, hippocampus, caudate putamen, and substantia nigra. 24 Furthermore, CCR2 colocalizes with dopaminergic neurons in the substantia nigra pars compacta and ventral tegmental area, as well as cholinergic neurons in substantia innominata and caudate putamen, suggesting potential neuronal modulating roles of CCR2. 24 Examinations using receptor radiography identified regions of CCL2 binding sites similar to CCR2 receptors identified using immunohistochemistry. 24 CCR2 protein has also been detected in reactive astrocytes of the spinal cord in a mouse model of amyotrophic lateral sclerosis, 25 and in neurons and astrocytes of the hippocampus after status epilepticus in rats. 26 However, some immunohistochemical findings of CCR2 expression were not confirmed by a recent study using CCR2-red fluorescent protein (RFP) knock-in mice. In that study, CCR2-RFP+ cells could not be detected in healthy mouse brain, nor in neuronal, microglial, or epithelial cells in brains from CCR2-RFP knock-in mice with experimental autoimmune encephalitis (EAE). 17 It is noteworthy that common complications associated with using knock-in animal models include position-effect variegation and stochastic gene silencing, which could potentially lead to inactivation of genes in certain cells and affect the outcomes of results. 27
PHYSIOLOGIC FUNCTIONS OF CCR2
Our current understanding of the role of CCR2 under physiologic conditions is limited. CCR2-knockout mice show no obvious deficits in growth, development, and fertility compared with wild-type animals when maintained in a pathogen-free environment. 9 However, a recent study suggests that CCL2/CCR2 is involved in the proliferation and growth of decidual stromal cells, which may play a role in the establishment and maintenance of early pregnancy. 28 CCR2−/− mice do exhibit some abnormalities in pulmonary vascular development, possibly via dysregulation of Notch signaling, which contributes to more severe pulmonary arterial hypertension after hypoxic stress compared with wild-type mice. 29
The most prominent role of CCR2 is believed to be in the mobilization of monocytes under physiologic as well as inflammatory conditions. CCR2−/− mice have an increased number of monocytes in the bone marrow and a decreased number of circulating monocytes, consistent with an important role of CCR2 in monocyte egress from bone marrow into the peripheral circulation. 30
CCR2 also appears to play an important role in maintaining physiologic levels of chemokines. CCR2−/− mice have high levels of circulating CCL2, which may in turn downregulate other chemokine receptors such as CCR1. 31 In vitro studies have shown that CCR2 mediates clearance of its ligand CCL2, 32 and CCL2 is consumed by migrating inflammatory cells as they cross the blood-brain barrier, 33 suggesting autoregulatory feedback mechanisms between CCR2 and CCL2 (Figure 1). Furthermore, a recent study suggests a potential role of CCR2 in bone homeostasis, as CCR2−/− mice have an increased bone mass owing to a decrease in the number, size, and function of osteoclasts. 34 It is also suggested that CCR2 is involved in the transmigration of bone-marrow-derived fibroblasts, as CCR2−/− mice exhibit impairments in accumulation of myeloid fibroblasts, and in myofibroblast formation in a mouse model of renal fibrosis. 35
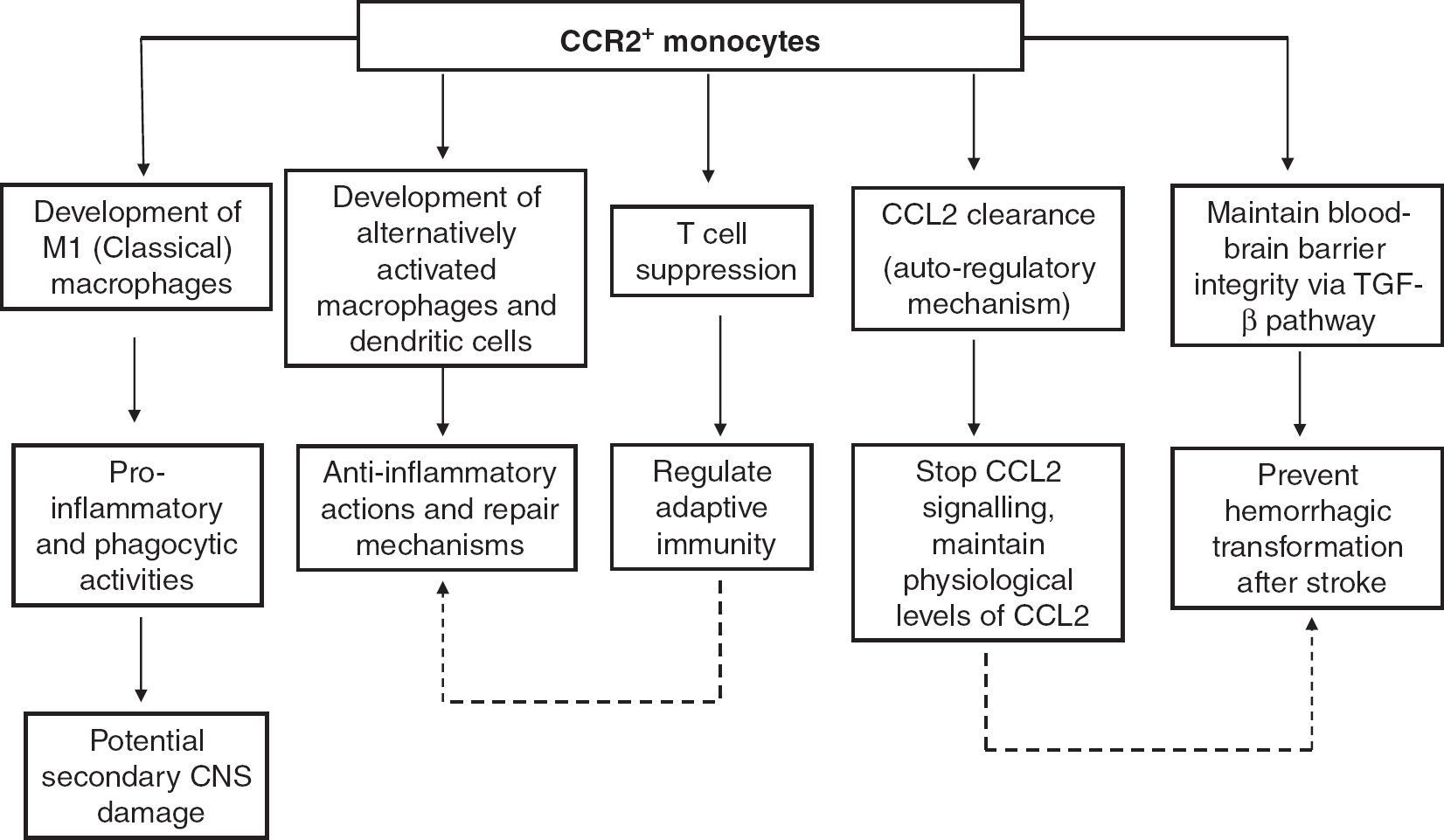
Schematic diagram depicting multiple potential roles of CCR2+ (CC chemokine receptor 2) monocytes during inflammation of the CNS. CNS, central nervous system; TGF-β, transforming growth factor.
CCR2 IN PATHOLOGIC ASSESSMENT
CCR2 appears to play an important role during inflammatory conditions. A study in CCR2−/− mice suggested that CCR2 is important for extravasation, adhesion, and transmigration of monocytes into inflamed tissue. 10 Further, it is thought that CCR2 is essential for the migration of Ly6Chi monocytes from bone marrow to peripheral circulation under both homeostatic and inflammatory conditions, with CCL2 and CCL7 being the agonists responsible for CCR2-dependent monocyte migration from bone marrow. 30
In particular, CCR2-dependent recruitment of monocytes is an important feature of innate host defense mechanisms. For example, Ly6Chi monocytes are recruited to the site of pathogen infection in a CCR2-dependent manner, and CCR2−/− mice have an inability to control replication of parasites, high influx of neutrophils, and rapid intestinal damage. 36 Furthermore, CCR2−/− mice have defects in delayed-type hypersensitivity responses and production of Th1 cytokines in mouse models of granulomatous lung disease and atherosclerosis. 9 Effects of monocytes can be regarded as a double-edged sword, promoting host defense and tissue repair, as well as causing further tissue injury during inflammation. In a model of myocardial infarction, CCR2-dependent infiltration of Ly6Chi monocytes is involved in the early phase of leukocyte influx after the ischemic insult, and these cells exhibit phagocytic and inflammatory activities. By contrast, Ly6Clo monocytes dominate the later phase of inflammation, expressing vascular endothelial growth factor, which promotes healing and angiogenesis. 37
EVIDENCE FOR A ROLE OF CCR2 IN CENTRAL NERVOUS SYSTEM DISEASES
Owing to the important role CCR2 plays in leukocyte trafficking, especially for monocyte recruitment to sites of inflammation and subsequent transformation to macrophages or dendritic cells, this chemokine receptor has been studied in numerous conditions involving inflammation within the CNS, including multiple sclerosis, Alzheimer's disease, and ischemic stroke. 6
Multiple sclerosis
In EAE, a common animal model of multiple sclerosis, CCR2 expression is increased in the priming and peak phases of the disease, and expression of the ligand, CCL2, correlates with the severity of the disease. 38 Interestingly, Fife et al 39 found that CCR2−/− mice have fewer infiltrating T cells and F4/80+ macrophages and are resistant to acute EAE. Furthermore, using three different mouse strains that were all deficient in CCR2, Gaupp et al 40 showed that CCR2−/− mice have a delayed symptom onset and reduced clinical severity in EAE compared with wild-type mice. Such evidence supports the concept that CCR2 blockade may have therapeutic potential for treatment of multiple sclerosis. However, this may not necessarily be the case as increased numbers of neutrophils were also observed in the EAE lesions of CCR2−/− mice, consistent with the existence of compensatory mechanisms in the absence of CCR2, which ultimately resulted in demyelination of neurons and full progression of the disease at a later time point. 40 Furthermore, it has also been suggested that CCR2+ monocytes are potential regulators of adaptive immunity, being powerful suppressors of activated T cells after EAE 41 (Figure 1). Hence, negating this aspect of CCR2 function would presumably serve to exacerbate the disease.
Alzheimer's disease
Alzheimer's disease is a neurodegenerative disorder characterized by the formation of intracellular neurofibrillary tangles and extracellular deposits of β-amyloid. 42 Increased levels of CCL2 have been observed in the brains of Alzheimer's disease patients and in transgenic mouse models of the disease. It has been shown that CCL2 expression is induced by increased levels of β-amyloid, and this can in turn disrupt the blood-brain barrier and facilitate recruitment of leukocytes into the CNS. 43 In mouse models of Alzheimer's disease, monocytes have been showed to migrate from the bone marrow and infiltrate into brain in a CCR2-dependent manner, and then differentiate into microglia or macrophages. There is growing evidence that CCR2+ cells may play protective rather than detrimental roles in Alzheimer's disease. CCR2−/− mice have been found to exhibit augmented cognitive impairment and amyloid pathology in a model of Alzheimer's disease. 44 Such features are associated with impaired microglial accumulation, which results in decreased clearance of β-amyloid and increased mortality. 45 Importantly, transplantation of CCR2 competent bone marrow cells restores cognitive capacity and reduces β-amyloid accumulation in CCR2−/− mice after Alzheimer's disease, 46 consistent with a protective role of CCR2-dependent microglial accumulation in the early stage of the disease.
Clinical studies have shown that monocytes from Alzheimer's disease patients have an impaired ability to differentiate into macrophages in vitro compared with monocytes from age-matched controls. 47 Monocytes from aging individuals have been reported to have impaired phagocytosis and increased levels of intracellular tumor necrosis factor-α, suggesting a dysregulation of monocyte function may exist in the aging population. 48 Together, the evidence currently suggests that Alzheimer's disease might involve impaired immune cell regulation involving CCR2.
Ischemic stroke
CCR2 and its ligand CCL2 are known to be involved in the inflammatory response of the injured brain after cerebral ischemia. Gene expression of CCL2 and CCL7 were found to be increased at both 24 hours and 72 hours after cerebral ischemia-reperfusion. 49 Globally, CCL2-deficient mice exhibit reduced phagocytic macro-phage accumulation in the brain and smaller infarcts after permanent middle cerebral artery occlusion compared with wild-type controls. 50 In vitro studies suggest a role for CCL2 in regulating blood-brain barrier permeability during transmigration of monocytes. Absence of CCR2 and CCL2 owing to treatment with neutralizing antibodies decreases the permeability of the brain endothelial barrier during in vitro ischemia-reperfusion. 51 Furthermore, CCR2-deficient mice are reported to have smaller infarcts, reduced blood-brain barrier permeability and reduced expression of inflammatory cytokines after ischemia-reperfusion compared with wild-type littermate controls. 52 Another study has shown that, compared with wild-type animals, CCR2-deficient mice have reduced levels of macrophages and neutrophils in the ischemic brain after ischemia-reperfusion, but without any difference in infarct volume. 53 There is also evidence that infiltrating CCR2+ Ly6Chi monocytes play a detrimental role in functional outcome after cerebral hemorrhage. 54 In addition, CCR2 deficiency has been reported to prevent hippocampal-dependent cognitive impairment in a mouse model of combined radiation/traumatic brain injury. 55 Thus, overall these data suggest that CCR2 inhibition could be beneficial for treatment of acute ischemic stroke. However, apart from causing secondary brain damage, it has also been suggested that bone-marrow-derived monocytes/macrophages recruited by CCR2 can prevent hemorrhagic infarct transformation via transforming growth factor-β signaling, and as such these cells may be important for maintaining neuro-vascular unit integrity after cerebral ischemia 56 (Figure 1). Moreover, there is evidence that CCR2/CCL2 mechanisms are important for the migration of neuroblasts from neurogenic regions to damaged regions of the brain after transient focal cerebral ischemia in mice, suggesting additional roles of CCR2 for tissue recovery after ischemic stroke. 57
Effects of CCR2 targeting in models of CNS diseases
Based on the above evidence for an involvement of CCR2 in a number of CNS inflammatory diseases, some studies have now evaluated CCR2 as a therapeutic target, particularly in multiple sclerosis for which there is strongest evidence for a detrimental role of CCR2. A series of CCR2 antagonists have been developed by Incyte (Wilmington, DE, USA), including INCB3344, which acts on both human and murine CCR2. 58 In a mouse model of EAE, INCB3344 (100 mg/kg daily) is reported to reduce CNS accumulation of monocytes/macrophages and to alleviate disease severity. A related compound, INCB8696, has entered a phase II clinical trial for multiple sclerosis. Merck (Whitehouse Station, NJ, USA) has also developed a potent CCR2 inhibitor, MK-0812, which has entered clinical trials for both rheumatoid arthritis and multiple sclerosis. However, the rheumatoid arthritis trial was terminated because of lack of favorable outcomes when MK-0812 failed to show any early clinical improvement. The outcome of the multiple sclerosis trial of MK-0812 is yet to be reported but it is suggested to also have negative outcomes. 59 To our knowledge, selective CCR2 antagonists have not yet been tested in animal models of ischemic stroke. However, a previous study using TAK-779, which is a dual CCR5/CCR2 antagonist, reported a dose-dependent reduction in infarct volume after transient cerebral artery occlusion, providing the first evidence that a chemokine receptor antagonist may be useful to protect against ischemic brain injury. 60
It is important to recognize that a number of factors may influence the outcome of CCR2 inhibition in CNS diseases (Figure 1). First, ligands of CCR2 can also act on other chemokine receptors, which may also contribute to immune cell infiltration into the brain. Thus, as more than one chemokine receptor may be involved in certain disease mechanisms, such redundant mechanisms may limit the effectiveness of CCR2 inhibition alone. Second, as recent evidence suggests that CCR2+ monocytes may regulate immunity by suppressing T-cell infiltration in EAE, 41 and these cells may develop into anti-inflammatory CD11c+ macrophages after spinal cord injury, 61 CCR2 antagonists could conceivably block these mechanisms and significantly limit their therapeutic benefit. Third, it is worth noting that the majority of studies suggesting the beneficial effects of CCR2 inhibition in either multiple sclerosis or ischemic stroke have been performed using lifelong globally CCR2-deficient animals, which may not adequately represent a clinical setting in which pharmacological intervention is started using a CCR2 antagonist. Lastly, it is important to consider possible off-target effects of CCR2 inhibition in specific disease settings. Central nervous system injuries such as stroke can lead to secondary immunodeficiency that markedly increases the body's susceptibility to infections that contribute to a greater overall morbidity and mortality.62,63 Indeed, it is established that CCR2 is important for the maintenance of levels of circulating monocytes, which are essential for immune surveillance and host defense. Any pharmacological manipulations targeting immune mechanisms must be carefully examined to avoid further impairment of host defense after CNS injury.
CONCLUSIONS
CCR2 plays an important role in the extravasation and transmigration of monocytes during inflammation. In inflammatory diseases of the CNS, CCR2 appears to control the entry of bone-marrow-derived Ly6Chi monocytes into the brain. Certain CNS conditions, such as Alzheimer's disease, might involve dysregulation of CCR2. CCR2-dependent recruitment of monocytes can have detrimental effects in other inflammatory diseases. Thus, there appears to be some rationale for predicting therapeutic benefits of CCR2 inhibition in multiple sclerosis and ischemic stroke, but there is also evidence that these cells may play important anti-inflammatory roles. Future studies examining the effects of pharmacological inhibition of CCR2 in CNS inflammatory diseases must therefore carefully evaluate such possibilities. As monocytes are highly dynamic immune cells with actions that can vary according to their environment, a thorough examination of the roles of CCR2+ monocytes in specific CNS disease settings is necessary to guide safe and effective development of any therapies involving CCR2 modulation.
Footnotes
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
CGS and GRD are Senior Research Fellows of the National Health and Medical Research Council of Australia (350327 and APP1006017).