
Abstract
The mechanisms of delayed damage and recovery after intracerebral hemorrhage (ICH) remain poorly defined. Two rodent models of ICH are commonly used: injection of the enzyme collagenase (cICH) and injection of autologous blood (bICH). In mice, we compared the effects of these two models on initial and delayed tissue damage, motor system connections, and behavioral recovery. There is no difference in lesion size between models. Injection of autologous blood causes greater mass effect and early mortality. However, cICH produces greater edema, inflammation, and cell death. Injection of the enzyme collagenase causes greater loss of cortical connections and secondary shrinkage of the striatum. Intracerebral hemorrhage occurs within the motor system connections of the striatum. Mapping of the projections of the forelimb motor area shows a significant sprouting in motor cortex projections only in cICH. Both models of ICH produce deficits in forelimb motor control. Behavioral recovery occurs by 5 weeks in cICH and 9 weeks in bICH. In summary, cICH and bICH differ in almost every facet of initial and delayed stroke pathophysiology, with cICH producing greater initial and secondary tissue damage and greater motor system axonal sprouting than bICH. Motor recovery occurs in both models, suggesting that motor system axonal sprouting in cICH is not causally associated with recovery.
INTRODUCTION
Stroke is a leading cause of death and disability across the world. Intracerebral hemorrhage (ICH) constitutes up to 15% of all stroke cases, and has clinical outcomes that are often worse than ischemic stroke.1,2 Mortality rates are as high as 50% 1 month after stroke, and survivors typically suffer life-long functional deficits. 3 Two well-established animal models of ICH are used to better understand the pathophysiology of the disease and the endogenous mechanisms of repair. These vary considerably in their mechanism. The infusion of the bacterial enzyme collagenase disrupts local blood vessels and produces endogenous hemorrhage into the site of the injection 4 but also substantial secondary inflammation. 5 Direct injection of autologous blood produces an immediate focal hemorrhage but is not derived from the native vasculature.5,6 There have been comparative studies of these models in the rat, but no comparative studies in the mouse and no studies of the mechanisms of axonal sprouting and neural repair in experimental ICH. Axonal sprouting is a prominent feature of tissue repair and recovery in non-hemorrhagic stroke7,8 and is causally associated with motor recovery in preclinical stroke models. 9 The present studies compare the immediate and long-term tissue damage from collagenase and autologous blood infusion models of ICH in the mouse, and the process of axonal sprouting and behavioral recovery of forelimb motor systems in this stroke subtype.
MATERIALS AND METHODS
Animals
All experiments were approved by the UCLA Chancellor's Animal Research Committee and performed in accordance with the National Institutes of Health animal protection guidelines. Two-month-old male C57/BL6 mice (Jackson Laboratories, Bar Harbor, ME, USA) were used in cohorts of 10 to 12 per group with a survival of 1 week, 5 weeks, and 9 weeks after stroke or sham procedure.
Models of Intracerebral Hemorrhage
Intracerebral hemorrhage was induced by the stereotactic injection of either autologous tail blood or of bacterial collagenase type VII (Sigma-Aldrich, St Louis, MO, USA) into the striatum. Mice were anesthetized with isoflurane and placed in a stereotaxic apparatus with constant anesthesia (1.5% to 2%) and core temperature maintained with a rectal probe (37° ± 0.5°). Correlation between brain and core temperature was established by taking corresponding temperature readings during bICH, cICH, or sham operation (
In the autologous tail blood injection (bICH), 20
Collagenase was diluted to a stock solution 0.115 U/
Mapping of Motor Cortex Connections
Changes in axonal density after stroke were assessed using intracortical injection of the neuroanatomical tracer biotinylated dextran amine (BDA 10000 MW, Life Technologies, Grand Island, NY, USA). Each treatment group (
All brains were fixed, frozen, and coronally sectioned 50
The same sections were subsequently cover slipped with Aqua-Poly/Mount (Polysciences, Warrington, PA, USA) and imaged with a Nikon C2 confocal microscope (Nikon Instruments, Melville, NY, USA) at × 10 for high-resolution BDA injection site images. Background MGV was measured for each section in the contralesional ventral cortex, which did not have axonal BDA label and reflected only background fluorescence. Individual injection volumes were determined by measuring the MGV of the × 10 injection images for the contiguous region of dense extracellular BDA, taking the sum of section means, and multiplying by 200
Higher MGV means more axonal fluorescence, and indicates more axonal projections. To validate this statement from results of the microarray scans, one section from each animal was stained with Cy-3-conjugated Streptavidin (Jackson ImmunoResearch), imaged, and measured with microarray scanning as described. Then axonal projections in the contralesional striatum were then directly quantified t × 40 magnification 9 (StereoInvestigator MBF Bioscience, Williston, VT, USA) on a Leica DMLB fluorescent microscope (Leica Microsystems, Wetzlar, Germany). The microarray fluorescence intensity was correlated with the direct axonal counts (GraphPad Prism 6, La Jolla, CA, USA).
Behavior
Paw-preference was determined by the percentage of single-handed rears in the cylinder test10,11 before ICH. Mice were categorized with preferences of left and right (
Histology and Immunohistochemistry
Brains were fixed, frozen, and coronally sectioned at 50
Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay was performed as described previously 12 on series of three sections using ApopTag In Situ Apoptosis Detection Kit (S7110, EMD Millipore). Briefly, sections were incubated with terminal deoxynucleotidyl transferase, which catalyzes the addition of nucleotide triphosphates labeled with digoxigenin to the 3-OH ends of double-stranded or single-stranded DNA (which localize in apoptotic bodies in high concentrations). The digoxigenin was then detected with an antibody conjugated to fluorescein. Nuclear stains were performed by incubating the sections for 5 minutes in phosphate-buffered saline containing propidium iodide before cover slipping. Cell nuclei tagged by TUNEL were manually quantified using neuroanatomical quantification software (StereoInvestigator, MBF Bioscience).
For mapping of Cholera Toxin Subunit B-injected brains, at the time of ICH induction, 0.5
Lesion or infarct size was measured in tissue sections stained with Cresyl violet.
14
Images at × 4 magnification were acquired (SPOT 5.0, Diagnostic Instruments, Sterling Heights, MI, USA) in six 50-
Brain edema was measured using the wet/dry weight method as previously described.
15
Briefly, animals (
The hemoglobin content of brains subjected to bICH, cICH, or sham (
Statistical Analysis
All tests were analyzed blindly to experimental condition. Animals were randomly assigned to treatment condition. Sample size was determined by a power analysis (Statistical Solutions, Cottage Grove, WI, USA), with the expected variance and change in performance in the grid-walking and cylinder rearing tasks predicted based on published studies.8–10,13 A group size of
RESULTS
Infarct Volume and Tissue Damage
There is no difference in lesion size between ICH models 24 hours after stroke (
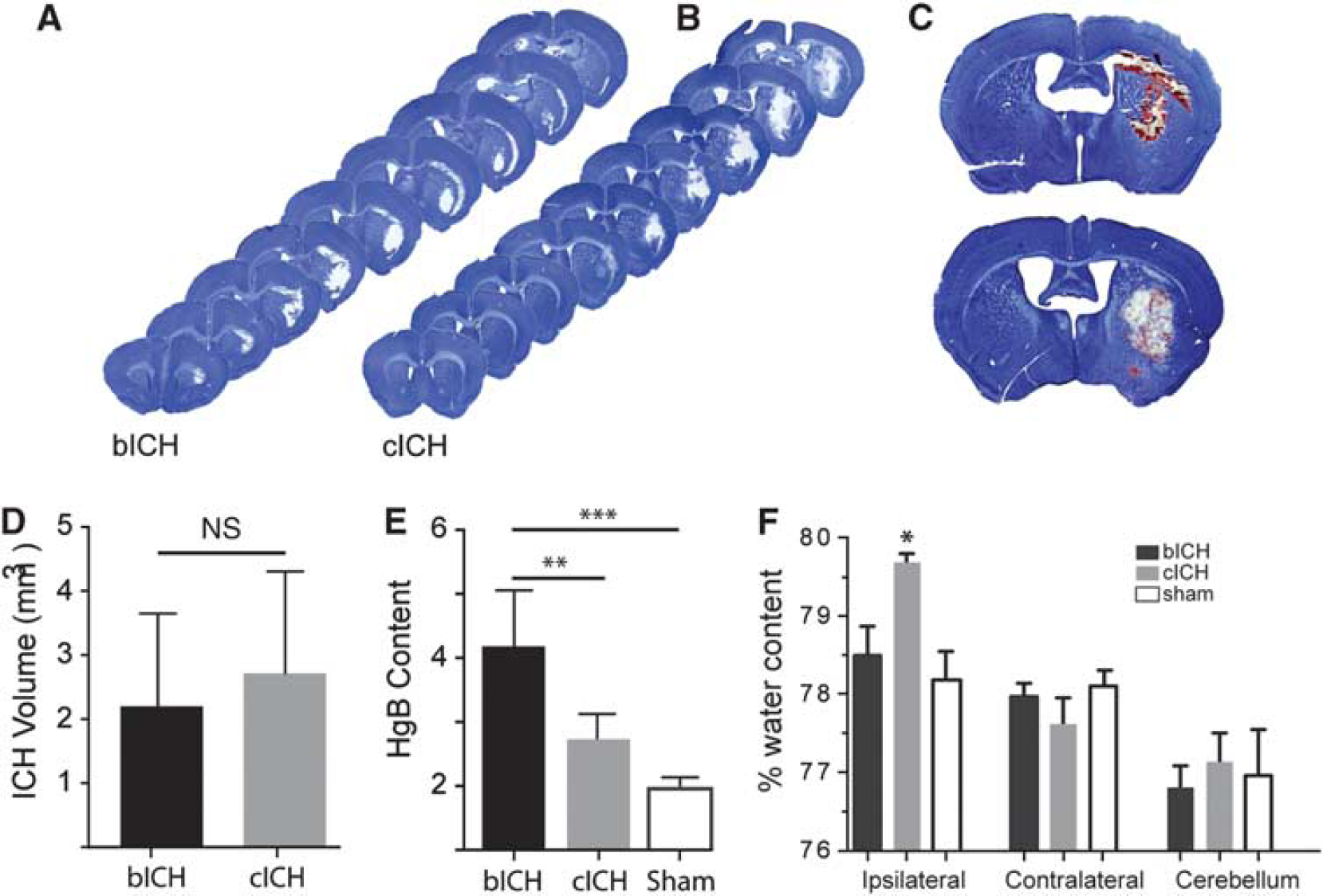
Initial tissue damage after intracerebral hemorrhage (ICH). (
Progressing damage was measured in both models. Hemispheric enlargement was greater in bICH, peaking at 3 days (Supplementary Figure 1E,
Inflammation, Gliosis Cell Death, and Axonal Loss after Intracerebral Hemorrhage
The inflammatory response to ICH was assessed at multiple time points after stroke. Iba-1 staining for microglia/macrophages after ICH showed an increase in staining in both models at 3 and 7 days, that is significantly increased compared with sham injection at 7 days (Figures 2A and 2B,
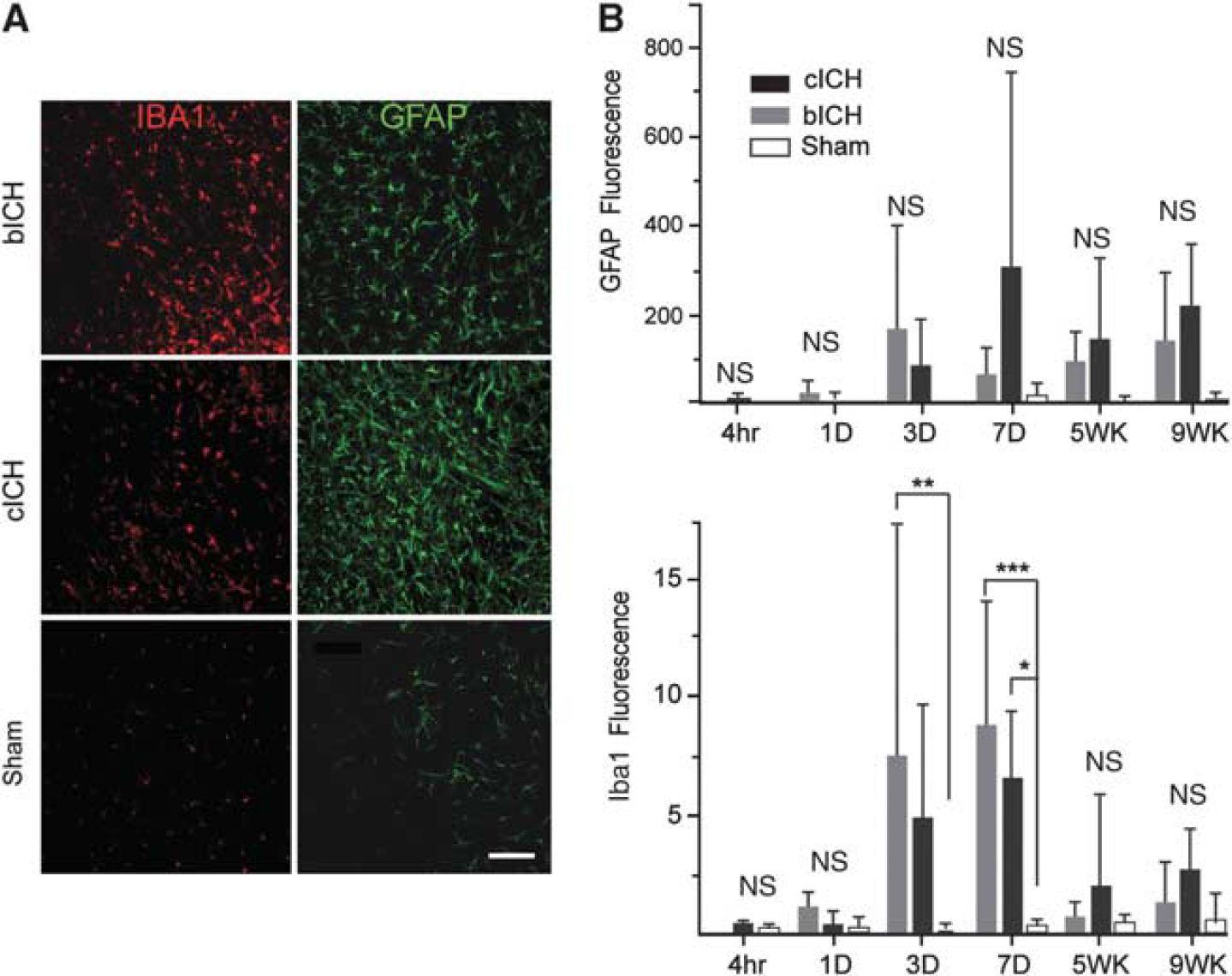
Inflammatory and astrocyte responses after intracerebral hemorrhage (ICH). (
Terminal deoxynucleotidyl transferase dUTP nick-end labeling staining 24 hours after stroke revealed extensive cell death throughout the infarct core, especially in cICH (Figures 3A and 3B). The number of TUNEL + cells did not differ between bICH and sham groups at the 24-hour time point (Figures 3A and 3C) but was significantly higher in cICH (**
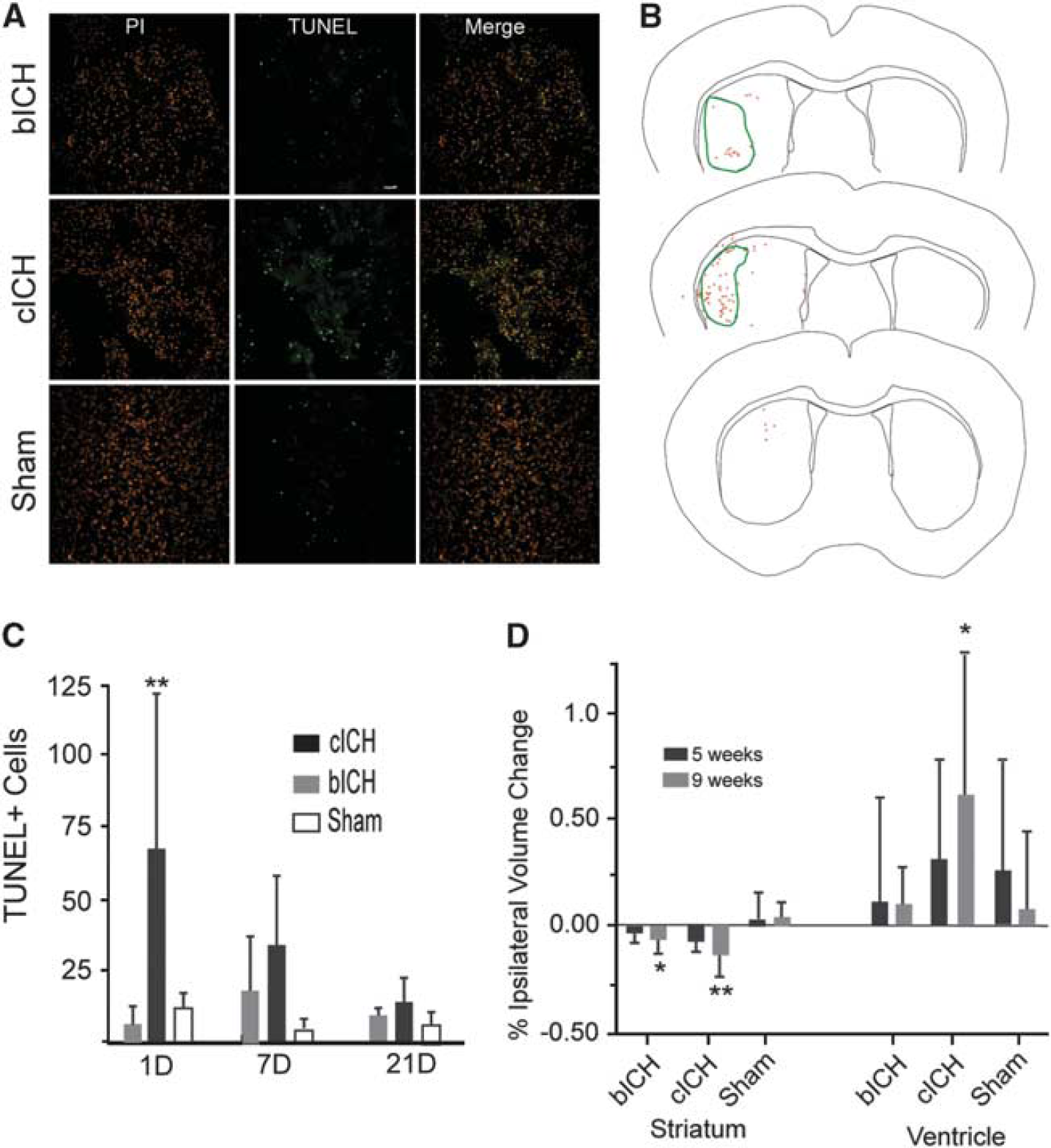
Cell death after intracerebral hemorrhage (ICH). (
In human and rodent ICH, the striatum is the predominant location of damage.
19
It is also a site of termination of axonal projections from cortex and a passage point where cortical projections pass through to thalamus, brainstem, and spinal cord.20,21 Intracranial hemorrhage damages these connections.
22
The degree of loss of cortical projections to or through the striatum was measured by back-labeling all of the cortical axons that project to the site of the ICH with the axonal tracer CTb, within the collagenase or blood injection, and then measuring the retrogradely labeled cell bodies. This approach labels cells that project to or through the hemorrhagic lesion,
23
allowing a quantification of the effect of the ICH on network connectivity of the overlying cortex. A large number of neurons in cingulate, frontal, and parietal cortex of both hemispheres project to the site of the ICH (Figures 4A and 4B). These include primary motor and somatosensory areas and secondary somatosensory areas bilaterally. There is a significant loss of cortical projections after ICH, particularly cICH. Collagenase-induced ICH causes loss of cortical projections from cortex ipsilateral and contralateral to the hemorrhage (Figure 4, *
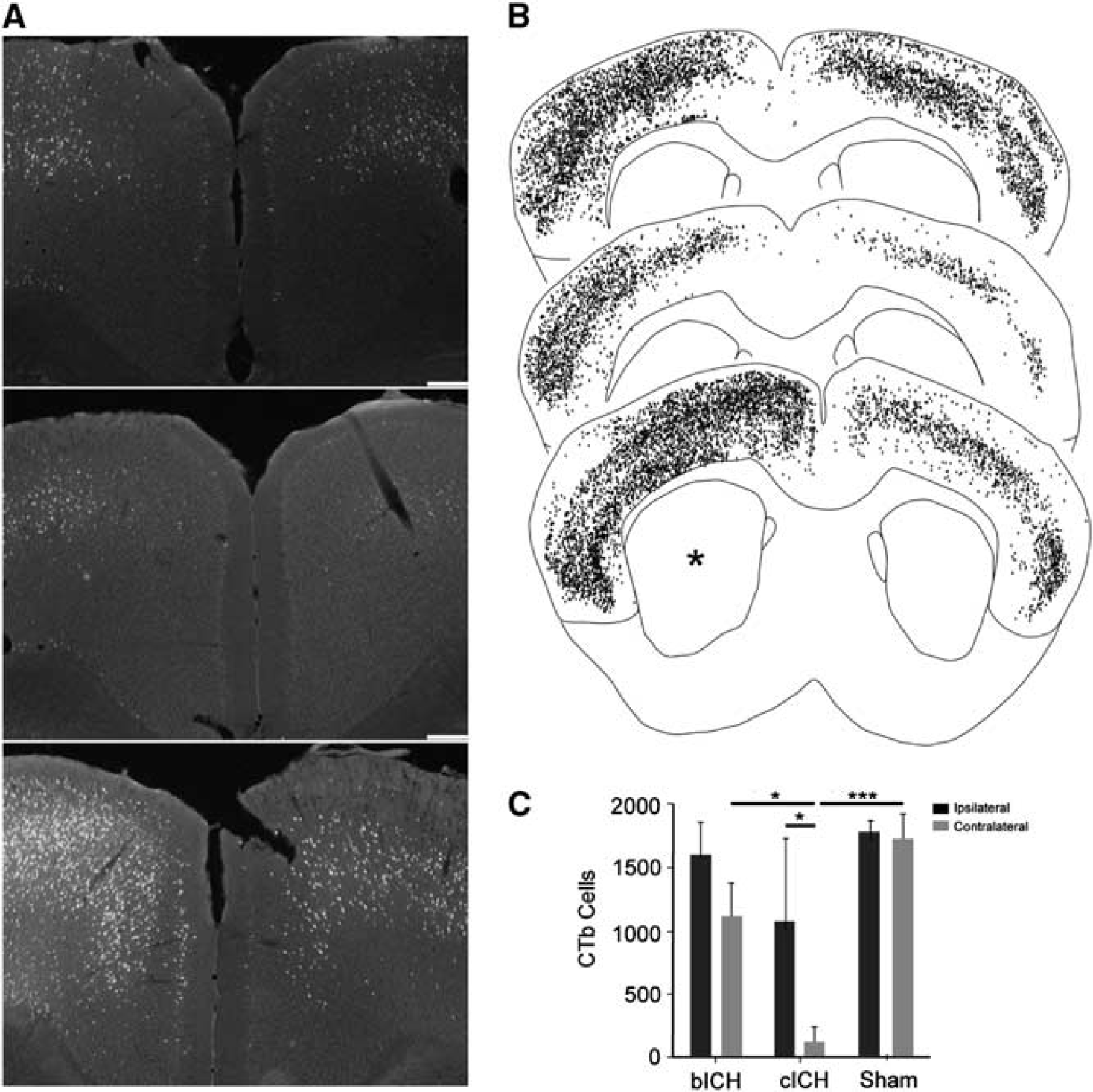
Axonal connections with striatum after intracerebral hemorrhage (ICH). (
Motor Impairments after Intracerebral Hemorrhage
Forelimb motor performance was assessed in both models. Animals performed similarly in a prestroke assessment of both grid-walking and cylinder rearing tasks. One week after stroke, bICH and cICH showed a significant motor deficit indicated by an increase in forelimb foot faults in the grid-walking task compared with baseline levels (Figure 5, **
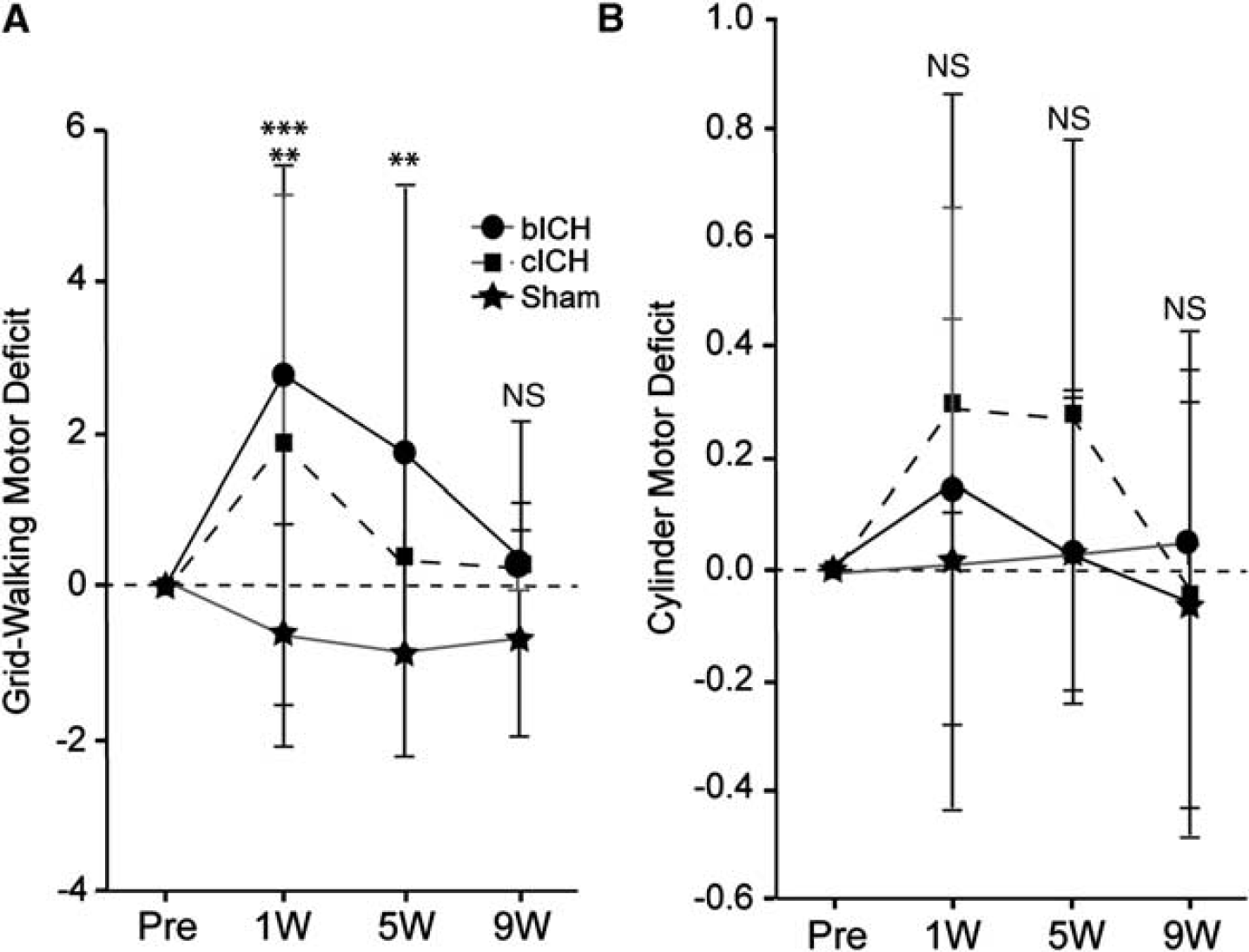
Motor impairments after intracerebral hemorrhage (ICH). (
Motor System Connections after Intracerebral Hemorrhage
Motor cortical projections have an important role in motor function and movement. In human and rodent ICH, the hemorrhage site is in the region of motor cortex projections within the ipsilateral hemisphere and from the ipsilateral hemisphere to contralateral cortex and striatum,
21
. The anatomical tracer BDA was injected into forelimb motor cortex
24
8 weeks after ICH. The axons were visualized with fluorescent labeling using a linear fluorescent high-resolution microarray scanner. To first determine that this measurement accurately and linearly measures axonal projections, BDA-labeled axons were counted and regressed with fluorescent measurements in the same area for all animals in this study (Supplementary Figure 2A). A statistically significant, linear correlation was shown between microarray scanner fluorescent intensity and number of axons, validating the accuracy of the microarray data (
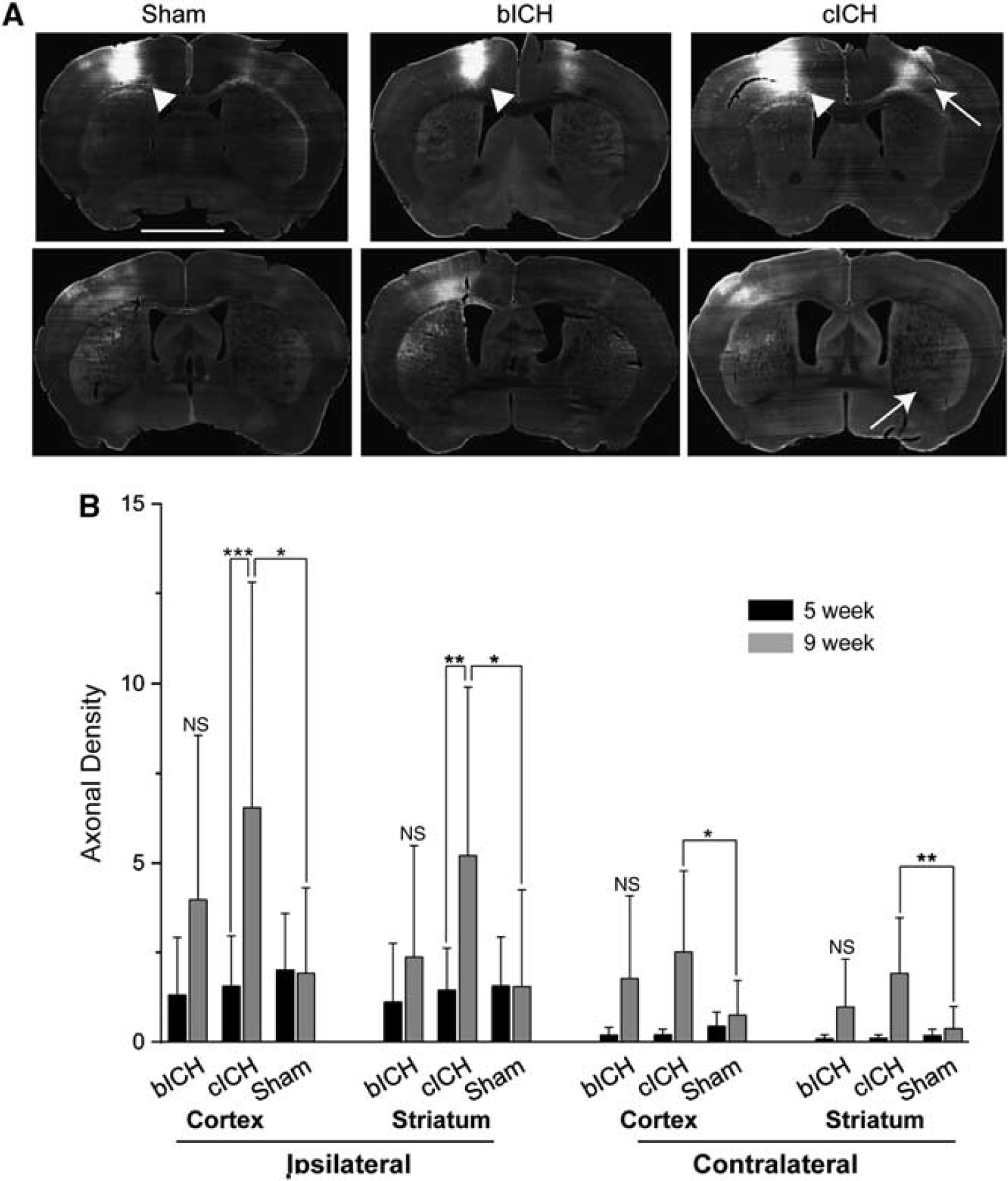
Motor cortex connections after intracerebral hemorrhage (ICH). (
At 9 weeks, corticostriatal projections are significantly increased in cICH compared with 5 weeks. Axonal density increased approximately fourfold in both the ipsilesional cortex and striatum, and over 10-fold in the contralesional cortex and striatum (Figures 6A and 6B). In the sham model, axonal labeling was similar between time points. While the bICH model showed the same increasing trends for motor system axonal connection as cICH, the changes in axonal density did not reach significance (Figure 6, CCX: bICH = 1.775 ± 2.304, cICH = 2.514 ± 0.2.259, sham = 0.753 ± 0.968, contralesional striatum: bICH = 0.982 ± 1.333, cICH = 1.916 ± 1.554, sham = 0.377 ± 0.617, ICX: bICH = 3.974 ± 4.581, cICH = 6.535 ± 6.269, sham = 1.926 ± 2.386, ipsilesional striatum: bICH = 2.375 ± 3.105, cICH = 5.199 ± 4.701, sham = 1.547 ± 2.701). Thus, cICH produces a significant axonal sprouting response from the ipsilateral cortex (above the ICH) to both the contralateral and ipsilateral cortex and striatum.
DISCUSSION
The development of novel therapeutics to support repair after stroke and ICH is dependent on accurate animal modeling of the disease. This study performed a direct comparison of tissue damage and long-term neurologic function in the two most widely used models of experimental ICH in the mouse, collagenase ICH (cICH) and autologous blood ICH (bICH). The two ICH methods produce equivalent lesion sizes. Collagenase intracerebral hemorrhage produces a lesion that more closely matches a hemorrhagic stroke, with bland tissue and less blood in the same-sized lesion cavity. Despite this equivalency in lesion size, cICH produces greater cell death, edema, inflammation, and delayed ventricular dilation, and bICH produces greater initial mass effect from the blood and higher mortality. Both types of hemorrhage destroy axonal projections to the striatum, but this loss was dramatic in cICH, with loss of over 95% of the projections from the cortex contralateral to the striatal hemorrhage site. Both models produced forelimb motor control deficits, with those in cICH recovering at 5 weeks and those in bICH recovering by 9 weeks. New motor projections form between 5 and 9 weeks in cICH from forelimb motor cortex to ipsilateral and contralateral cortical and striatal areas. This difference in tissue damage and long-term axonal sprouting patterns indicates that the two models used in ICH differ in almost every facet of initial and delayed effects of stroke and of reactive plasticity in the post-stroke brain.
In the present study, we modified injection of autologous blood and collagenase for the ICH. Initially, these ICH models were developed so the blood or the collagenase was injected directly down into the targeted brain area, the striatum. 25 However, these coordinates cause the needle to penetrate the mouse forelimb motor cortex, causing damage to this primary motor region and complicating the interpretation of a motor deficit, as the motor deficit may be due to the hemorrhage in the striatum or to the damage in the motor cortex from the needle en route to the striatum. To resolve this complication, the injection was done at a 10° angle, with the needle tract not passing through the forelimb motor cortex. We further improved the previous models of ICH by administering blood or collagenase via a smaller, 33-gauge needle. This modification prevented backflow commonly seen, especially in bICH, and further minimized damage from the technique itself outside of the hemorrhage site. This revised method in both models of ICH produced lesions that did not differ in size at any anterior/posterior level. The use of heparin in bICH did not induce additional bleeding when added as a test to the cICH model, but it may be associated with other effects not studied here. Both lesions targeted the dorsolateral striatum, the site of sensorimotor system function.20,21 However, although these lesions overlap across animals in their location within the dorsolateral striatum of the mouse, their precise anatomic positions were not matched.
Previous studies have compared collagenase and blood ICH models in the rat. The differences in the rat between these two models parallel many of those in the mouse. As in the present findings in the mouse, rat cICH produces greater edema, local neuronal loss in the striatum, and greater tissue atrophy; inflammation is more pronounced in the rat cICH model.26,27 As in the present study, the cylinder test of forelimb function shows a very modest cylinder effect in cICH in rats 28 or a short-term effect in cICH in mice. 29 Some of the effects of bICH and cICH in the mouse are distinct to that seen in the rat. Behavioral deficits persist for a greater period in rat cICH26,30 than in the mouse. Unlike in rat, sensorimotor deficits as measured by gait in the mouse resolve sooner in cICH than bICH. These two lesions models also have very distinct patterns of axonal loss and later axonal sprouting.
One of the main findings in this study is that there is a difference in axonal loss, long-term atrophy, and secondary axonal sprouting between cICH and bICH despite similarly sized lesions. Collagenase intracerebral hemorrhage produces greater shrinkage of the striatum and ventricular dilation than bICH. In fact, there is no difference in bICH in the striatal or ventricular size compared with sham. Collagenase intracerebral hemorrhage causes significantly greater loss of cortical projections, particularly in the projections from the somatosensory and motor cortical areas contralateral to the striatum, which are nearly completely lost after cICH. In rat cICH, there is loss of connections to the striatum with a modest reduction in neurons that project from the substantia nigra to the striatum, 31 a decrease in cortical projections from the ipsilateral motor cortex 32 and thinning of ipsilateral cortex. 26 There are no reports of axonal loss in the rat in bICH. This greater loss of axonal projections to the striatum in cICH in the mouse could be due to greater local tissue damage after the hemorrhage. Although the lesion size did not differ between the two models, there is greater cell death in hemorrhage tissue in cICH, edema, and inflammation. These may be measures of incomplete damage in apparently intact tissue adjacent to the hemorrhage after collagenase injection, rather than in autologous blood injection. This greater incomplete damage in perihematomal tissue in cICH is likely because damage in cICH reflects both hemorrhage and also bland (non-hemorrhagic) stroke (Figures 1C and 1E). In rat cICH, myelin is damaged and myelin debris is present 1 to 3 days after ICH in the margins of the hematoma and adjacent perihematomal tissue. 14 Axonal damage, as assessed by axonal accumulation of beta-amyloid precursor protein, retraction bulbs, and disintegrating axonal profiles, is seen in the first 3 days after cICH in the rat adjacent to the hemorrhage. 14 The present and published data suggest that cICH produces greater tissue injury in perihematomal striatum and that this leads to greater secondary tissue atrophy and loss of projections to the striatum from cortex.
This is the first report of spontaneous axonal sprouting after ICH. While there is an increase in axonal projections from the forelimb motor cortex ipsilateral to the lesion in both models, this was only significant for cICH. In fact, cICH produces dramatic axonal sprouting from the forelimb motor cortex above the striatal hemorrhage, such that there is a three to fivefold increase in axonal projections from motor cortex. Axonal sprouting occurs in non-hemorrhagic stroke, from cortex opposite to the stroke to striatum, brainstem, and spinal cord and in the cortex adjacent to the stroke. 7 In the cortex adjacent to the stroke, axonal sprouting forms new projections that link motor cortex to premotor and somatosensory areas. 8 When the formation of these new projections from motor adjacent to the stroke to premotor cortex is blocked, mice do not experience motor recovery. 9 These data establish a causal role for post-stroke axonal sprouting in behavioral recovery in some mouse stroke models. In cICH, post-hemorrhage axonal sprouting occurred between 5 and 9 weeks after the hemorrhage was induced. This means that axonal sprouting occurred after mice had recovered motor function, which occurred at 5 weeks after hemorrhage. Further, mice with cICH and bICH recovered to the same behavioral performance despite the very different degrees of axonal sprouting between these two ICH models. The inference here is that axonal sprouting in cICH, unlike that seen in non-hemorrhagic stroke in the mouse, is not associated with functional recovery.
Human ICH results in disconnection syndromes in overlying cortex, and recovery is associated with the strength of functional connectivity in sensorimotor areas. Patients with ICH in the striatum show lower measures of connectivity from the ipsilateral hand motor cortex to contralateral motor, striatal/basal ganglial and subcortical sites.33,34 This parallels cICH in the present study, where connections from the mouse forelimb motor cortex ipsilateral to the lesion drop out after the hemorrhage. These parallels between connectivity patterns in human ICH and mouse cICH raise the important issue of which mouse ICH model might be considered to most closely model the human condition. Damage after ICH in humans can occur with local displacement of brain structures from blood, mass effect from edema, and toxicity of blood breakdown products. 2 Edema after ICH produces a high early mortality. 2 The initial mass effect and higher mortality in mouse bICH models these early events in human ICH. However, in mouse cICH, there is greater overall tissue damage and a component of non-hemorrhagic stroke. Thus, the pattern of atrophy and loss of connections would suggest that cICH produces a pattern of injury seen in more severe human ICH and bICH produces a pattern of damage seen in less severe ICH. However, functional recovery occurs in both models. Thus, there are four conclusions from detailed studies of mouse ICH models: cICH produces a pattern of damage and disconnection that models moderate to severe human ICH; bICH produces a pattern of initial lesion progression that models human ICH; the patterns of behavioral recovery in the mouse are disassociated from the underlying brain damage; and reactive axonal sprouting in cICH is not linked to functional recovery.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.