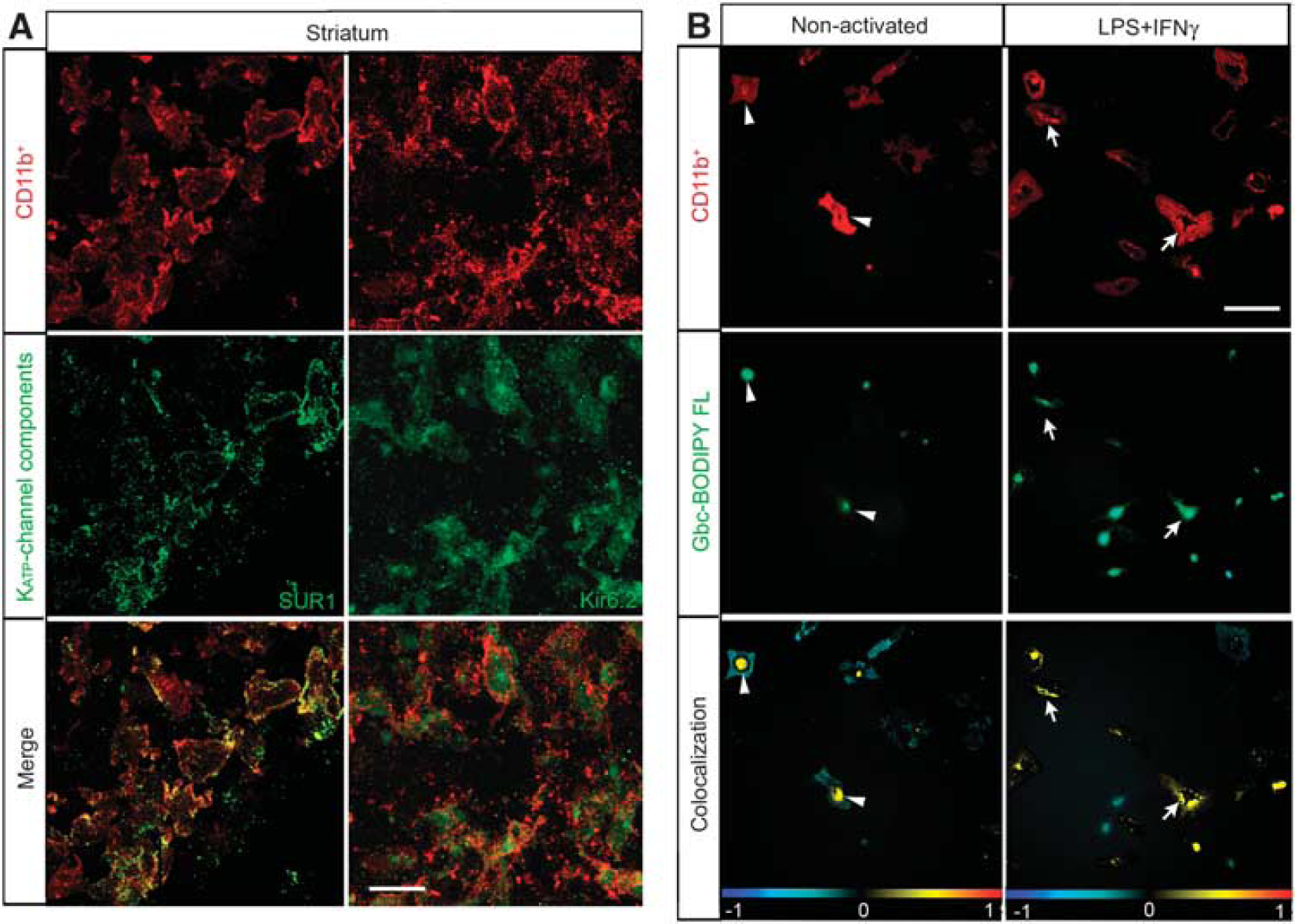
The recent review article ‘Sulfonylurea receptor 1 in central nervous system injury: a focused review'1 is an extensive summary of the current knowledge of the role of the sulfonylurea receptor 1 (SUR-1) and SUR-1-regulated NCCa_ATP channels in acute brain injuries. The review also highlights the potential translational applicability of the use of glibenclamide in treating brain pathologies such as cerebral ischemia or traumatic brain injury. It is proposed that the cellular targets of glibenclamide are neurons, endothelial cells, oligodendrocytes, and astrocytes, and that glibenclamide resolves the cytotoxic edema after cerebral ischemia by blockade of the astroglial NCCa-ATP channel. However, independent studies have shown that the SUR-1-formed ATP-dependent potassium (KATP) channels are expressed by microglia,2–6 which reinforces the idea that glibenclamide may also target microglia and modulate their inflammatory phenotype in various brain pathologies. More important, we recently showed that microglia after cerebral ischemia increase the expression of Kir6.2 and SUR-1 components of the KATP channel in the lesion core3 and also in the medial striatum of the ischemic hemisphere4 (Figure 1A).
Reactive microglia express and translocate sulfonylurea receptor 1 (SUR-1) to the cell surface. (A) Confocal photomicrographs of SUR-1 and Kir6.2 (green) in reactive microglia (CD11b-positive; red) localized to the medial striatum in middle cerebral artery occlusion rats. Yellow in the merge image denotes colocalization, whereby reactive CD11b-positive cells expressed SUR-1 or Kir6.2 72 hours after ischemia. (B) Localization of glibenclamide (Gbc) (Gbc BODIPY FL; green fluorescence) in rat microglial primary culture. Non-activated or culture-activated lipopolysaccharide (LPS) + interferon gamma (IFNγ) for 48 hours are shown in the upper row. Microglial cells were labeled with an anti-CD11b (red) antibody and Hoechst (blue) to stain the nuclei. Lower row shows respective colocalization of the red and green channels, where the yellow denotes the presence of the Gbc binding in microglial cells. Arrowheads denote perinuclear colocalization and arrows show surface labeling. The data shown are representative of four experiments each. Scale bar in (A) is 15 μm and in (B) is 20 μm (from Ortega et al.4).
KATP channel subunits possess an endoplasmic reticulum-retention motif, which prevents trafficking of mismatched subunits to the membrane. Our studies in rat primary microglial cultures suggested that microglial activation involves translocation of SUR-1 from its internal reservoir toward the cell surface (Figure 1B). Furthermore, in vitro studies using BV2 microglia cells and primary microglial cultures have demonstrated that reactive microglia are sensitive to different KATP channel drugs regulating the phagocytic activity and the release of cytokines and chemokines.2,3,5, Our findings of the enhancement of microglial in vitro phagocytosis were correlated with in vivo experiments, where increased clearance of cell debris and calcium was found in the infarcted hemisphere, and consequently provided an optimal neuroprotection in the surrounding tissue.3 Although glibenclamide also blocks other ion channels, of which some belong to the ATP-binding cassette proteins (e.g., the cystic fibrosis transmembrane conductance regulator) expressed by microglial cells, it is unlikely that glibenclamide will bind to these channels because they present much lower affinity to the drug than that used in our studies.3,4 Thus, these findings shed a new light on the putative role of the microglial KATP channel in mediating at least, in part, the neuroprotective and neurorestorative effects of glibenclamide after stroke.
Simard et al7 have described that the activation of NCCa-ATP channels in astrocytes causes cell-blebbing characteristic of cytotoxic edema. The glibenclamide-induced beneficial effect in middle cerebral artery occluded (MCAO) animals was only linked with the blockade of these channels, whereas the involvement of KATP channels in the process has been ignored.7,8 Interestingly, Simard et al7 only used inside-out patches of large neuron-like cells isolated from the core 2 hours and 6 hours after MCAO or isolated native reactive astrocytes type 1.9 The KATP channel biophysical properties in other cell types isolated from the core or the periinfarct area after brain ischemia, which could be also expressing SUR-1 regulated channels, were not assessed. Intriguingly, despite the massive neuronal loss observed, immunoblots revealed no concentration changes of Kir6.1 and Kir6.2 proteins in the ischemic core.7,10 Our findings are consistent with these results and argue for a contribution of the microglial KATP channels to the neurorestorative effects of glibenclamide by reducing the severity of ischemic damage. We observed that reactive microglia enhances SUR-1, Kir6.1, and Kir6.2 protein expression, and ameboid microglia express KATP channels in the necrotic core of the lesion.3,4 Therefore, this upregulation is certainly contributing to the enhancement of SUR-1 found by Simard et al,7 and may help to compensate for a putative decrease in Kir6.1 and Kir6.2 subunits owing to the massive neuronal loss in the lesion core.
However, we also observed that glibenclamide increased the number of migrating neuroblasts toward the ischemic core 72 hours after reperfusion, thereby indicating that glibenclamide modifies the cell lineage choice or enhances progenitor cell proliferation and migration.4 As adult neural precursor cells do not express the Kir6.1 nor the Kir6.2 subunits, it is unlikely that they will express functional KATP channels or present sensitivity to the glibenclamide treatment. However, microglia, which are the primary immune effector cells in the brain and are a component of the neurogenic niche, participate in promoting the proliferation, migration, and differentiation of neural precursors cells by the release of a wide variety of bioactive molecules, including neurotransmitters, purines, cytokines, and growth factors. Interestingly, reactive microglia in the medial striatum expressed the KATP channel components SUR-1 and Kir6.2 at 72 hours after ischemia (Figure 1A), suggesting participation over longer time intervals in postischemic regeneration and neurogenesis. Complementary in vitro studies showed that the specific blockade of the microglial KATP channel cause the release of soluble factors that enhance the activation of neural precursor cells from the subventricular zone (unpublished data). Therefore, ongoing studies will help us to elucidate the complex interplay of ischemic and inflammatory signals responsible for the role of the microglial KATP channel in endowing microglia to a new distinct phenotype promoting brain repair after injury.
Taken together, the involvement of the KATP channel expressed by microglia can contribute to the beneficial effects of glibenclamide on stroke models. Our data on the glibenclamide-mediated control of the microglia activity through KATP channel blockade argues for a multifunctional neuroprotective effect of SUR targeting after brain injuries. This is consistent with the idea of cross-talk between multiple cell types and death mechanisms after cerebral ischemia. By briefly summarizing the current state of knowledge in this area, this commentary hopefully provides a new and complementary insight on the neuroprotective and neurorestorative effects of glibenclamide after stroke.
ETHICS STATEMENT
All of the experiments reviewed were approved by the Ethics Committee of the Universitat de Barcelona, in accordance with the regulations from the Catalan government (Generalitat de Catalunya) or the Animal Ethics Committee (Hämeenlinna, Finland). Animals were handled following European legislation (86/609/EEC) and all efforts were made to minimize the number used and animal suffering, in accordance with the ARRIVE guidelines.
DISCLOSURE/CONFLICT OF INTEREST
MJR holds an EU patent (No. WO2006/000608). The remaining authors declare no conflict of interest.
References
1.
SimardJMWooSKSchwartzbauerGTGerzanichV. Sulfonylurea receptor 1 in central nervous system injury: a focused review. J Cereb Blood Flow Metab2012; 32: 1699–1717.
2.
LiuXWuJ-YZhouFSunX-LYaoH-HYangYThe regulation of rotenone-induced inflammatory factor production by ATP-sensitive potassium channel expressed in BV-2 cells. Neurosci Lett2006; 394: 131–135.
3.
OrtegaFJGimeno-BayonJEspinosa-ParrillaJFCarrascoJLBatlleMPuglieseMATP-dependent potassium channel blockade strengthens microglial neuroprotection after hypoxia-ischemia in rats. Exp Neurol2012; 235: 282–296.
VirgiliNEspinosa-ParrillaJFManceraPPastén-ZamoranoAGimeno-BayonJRodríguezMJOral administration of the KATP channel opener diazoxide ameliorates disease progression in a murine model of multiple sclerosis. J Neuroinflammation2011; 8: 149.
SimardJMYurovskyVTsymbalyukNMelnichenkoLIvanovaSGerzanichV. Protective effect of delayed treatment with low-dose glibenclamide in three models of ischemic stroke. Stroke2009; 40: 604–609.
9.
ChenMDongYSimardJM. Functional coupling between sulfonylurea receptor type 1 and a nonselective cation channel in reactive astrocytes from adult rat brain. J Neurosci2003; 23: 8568–8577.
10.
SimardJMTsymbalyukNTsymbalyukOIvanovaSYurovskyVGerzanichV. Glibenclamide is superior to decompressive craniectomy in a rat model of malignant stroke. Stroke2010; 41: 531–537.