
Abstract
Damage of oligodendrocytes after ischemia has negative impact on white matter integrity and neuronal function. In this work, we explore whether Netrin-1 (NT-1) overexpression facilitates white matter repairing and remodeling. Adult CD-1 mice received stereotactic injection of adeno-associated virus carrying
INTRODUCTION
Oligodendrocytes produce the main components of myelin in the brain, mainly derived from the maturation of oligodendrocyte progenitor cells (OPCs) during development or after brain injury in adulthood. 1 Myelins are essential for axon integrity and functional electrical impulse transmission. Damage to oligodendrocyte leads to disruption of myelin structure and function, exacerbating motor function and other neurobehavioral performances. 2 For example, neurobehavioral outcomes are greatly destroyed when myelins are damaged in multiple sclerosis. 3 Cerebral ischemia not only damages gray matter neurons but also white matter glial cells including oligodendrocytes and myelin. Because of relatively low blood supply during ischemia, oligodendrocytes are particularly susceptible to ischemic damage due to excitotoxicity, oxidative stress, and other cellular mechanisms. 4
In the past decade, neuroprotective drugs were proved effective in experimental stroke but failed in human clinical trials. One reason may lie in that these drugs are targeted at gray matter. However, human brain consists of two major components with almost the same volume. 5 Protection and remodeling on white matter are often neglected, especially in the postischemic recovery period. Therefore, additional attention should be paid to white matter protection in acute ischemic stroke and repairing during recovery phase. White matter repairing refers to increasing endogenous OPCs, which occupy 5% to 8% glial cells in the brain. Oligodendrocyte progenitor cells can differentiate into mature oligodendrocytes and participate in remyelination during central nervous system injuries including cerebral ischemia.
During the spontaneous recovery period after ischemic stroke, angiogenesis, neurogenesis, and oligodendrogenesis occur to promote tissue repairing and remodeling. 6 However, the endogenous recovery process is usually insufficient. New approaches should be considered to promote more aspects of recovery progress after ischemic brain injury.
Netrin-1 (NT-1), a lamina-related protein, has a pivotal role in axon guidance. During central nervous system development, NT-1 either attracts or repels axon outgrowth depending on its binding receptors. 7 Netrin-1 also influences small blood vessel development and tissue morphogenesis.8, 9 Another important function of NT-1 in development is to affect white matter development by influencing OPC proliferation, differentiation, and migration.10, 11 Under pathologic conditions, NT-1 has many roles in various systems. It inhibits inflammation and apoptosis and promotes repairing after ischemic stroke by increasing angiogenesis.12, 13, 14, 15 However, the effect of NT-1 on white matter repairing after ischemic brain injury has not been explored yet. In the present study, we aim to investigate whether NT-1 overexpression could improve white matter repairing and remodeling after ischemic stroke, and if so, the possible role of NT-1 will be further studied.
MATERIALS AND METHODS
Animals and Experimental Design
This study was carried out in accordance with ARRIVE guidelines. Animal experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC), Shanghai Jiao Tong University, Shanghai, China. Mice were housed under standard laboratory conditions. Adult male CD-1 mice(
Adeno-Associated Virus-Netrin-1 Virus Production
Adeno-associated virus was packaged and titered as described previously.
16
Briefly,
Adeno-Associated Virus-Netrin-1 Gene Transfer into Mouse Brain
Adult CD-1 mice were anesthetized with ketamine/xylazine (100 mg/10 mg per kg, Sigma, San Louis, MO, USA) intraperitoneally. The mice were fixed on a stereotaxic plate (RWD, Shenzhen, China). A 0.5 mm bone hole was drilled by a hand drill (Fine Science Tool, foster City, CA, USA). Five microliters of AAV-NT-1 (containing 1 × 1011 particles) was slowly injected into the left striatum AP=−0.02 mm, ML=−2.5 mm, DV=3 mm relative to the bregma using a mini pump (WPI, Sarasota, FL, USA). The needle was kept in the brain for additional 5 minutes and after injection, the needle was withdrawn. The bone hole was sealed, the wound was sutured, and animals were returned to their cages after awake. 19
Transient Middle Cerebral Artery Occlusion in Mice
After 1 week of
Immunofluorescent Staining
After sacrificing animals, the brains were removed and immersed in 4% paraformaldehyde fixation solution at 4°C overnight. After fixation, brains were dehydrated in 30% sucrose solution and sectioned in 30-
BrdU Staining and Cell Counting
BrdU powder (Sigma) was dissolved in normal saline in a concentration of 65 mmol/L. BrdU solution was injected intraperitoneally at 50 mg/kg twice a day for two time phases for 3 consecutive days: 4 to 6 days after MCAO surgery and 3 days before killing animals. For BrdU double staining, sections were first treated with 2 mol/L HCl for 30 minutes at room temperature and then with sodium borate twice each for 10 minutes. Sections were then treated with 0.3% Triton–PBS for 30 minutes, blocked by 5% normal donkey serum, incubated with anti- BrdU (1:200 dilution, Santa Cruz), CD140
Western Blot Analysis
Protein used in western blot experiments was isolated from the striatum. Mice were anesthetized by ketamine/xylazine intraperitoneally. After anesthesia, brains were quickly removed to a cooled brain mold, and then cut into four sections by five blades that are 2 mm apart; the second rostral section including ischemic core was collected. The section was divided into contralateral cortex, striatum and ipsilateral cortex, striatum using micro forceps under microscope. The protein extracted from the ipsilateral striatum was used for further western blot analysis. For protein extraction, tissues were first homogenized by ultrasonic on ice and then centrifuged. Equal amount of total protein (40
Neurobehavioral Tests
Mice were trained for 3 consecutive days before tMCAO surgery. Neurobehavioral tests were performed before MCAO and at 1, 2, and 4 weeks after tMCAO by an investigator who was masked to the experimental groups. 20
Rotarod Test
An accelerating rotarod (Zhenghua, Anhui, China) provides an index of motor coordination and balance with its velocity slowly increasing from 5 to 40 r.p.m. The duration that mice maintained on the accelerating rotarod was measured. Each animal was given three trials and the longest time animals spent on the rod was recorded.
Beam Walk
For beam walk test, mice were trained to traverse a horizontally elevated square beam with 7 mm side length to reach an escape platform placed 1 m away. Mice were placed on one end of the beam and the latency to traverse 80% of the beam towards the escape platform was recorded. Data from motor tests were analyzed as mean latency to cross the beam from three trials.
Neurologic Score
Modified neurologic severity scores of the animals were graded on a scale of 0 to 14, which is a composite of motor, reflex, and balance tests. 21
Statistical Analysis
Data were presented as mean±s.d. Bonferroni
RESULTS
Netrin-1 Overexpression in the Mouse Brain
Netrin-1 expression was examined 2 weeks after
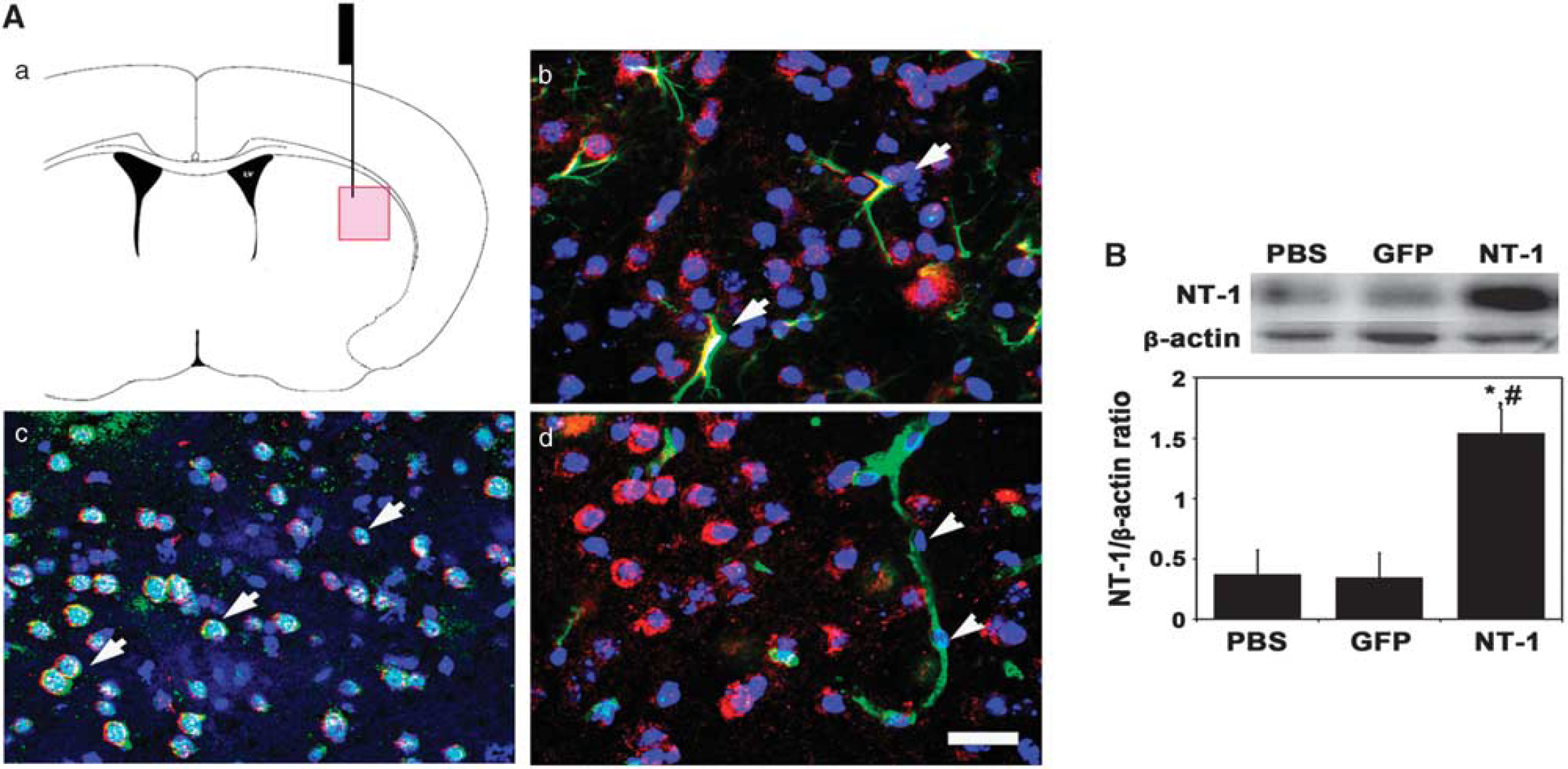
Netrin-1 (NT-1) overexpression in the mouse brain after adeno-associated virus
Netrin-1 Overexpression Improved Neurobehavioral Outcomes
As indexes of motor function, the beam walk test, and rotarod test in
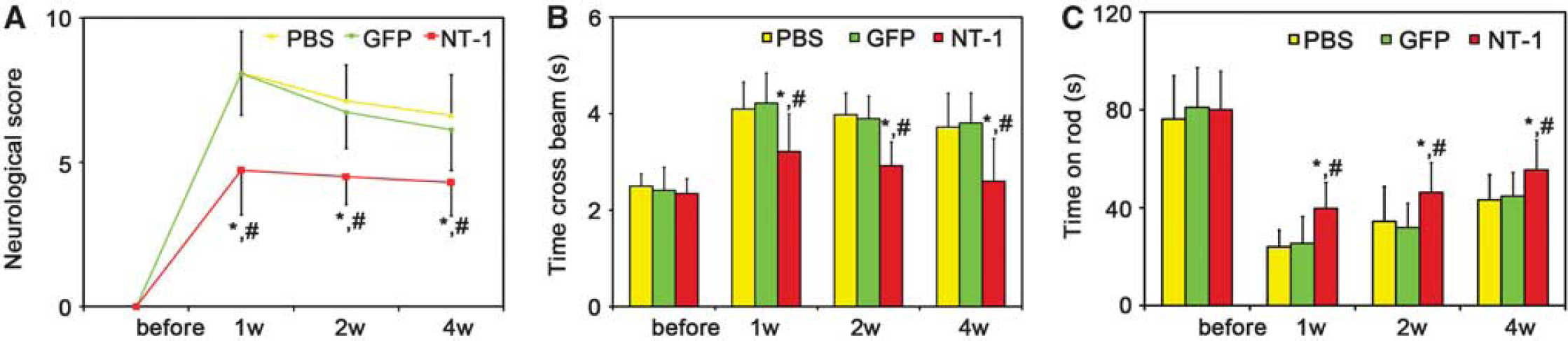
Netrin-1 (NT-1) overexpression improved the behavioral outcomes. The neurologic score (
Netrin-1 Overexpression Improved Oligodendrocyte Progenitor Cell Proliferation and Maturation
Brain sections were double labeled with CD140
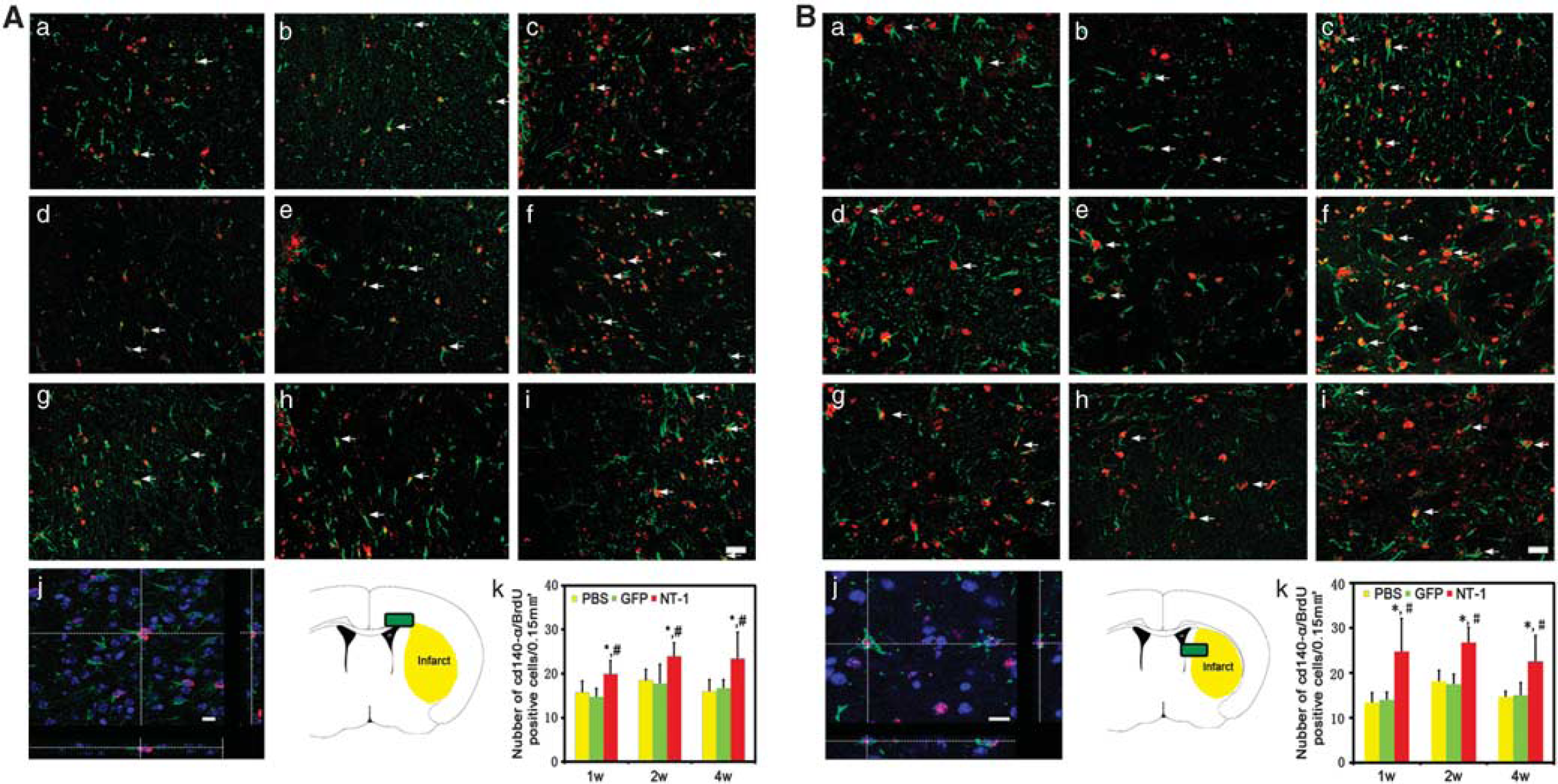
Proliferated oligodendrocyte progenitor cells (OPCs) in corpus callosum and striatum. Double-labeled BrdU and CD140
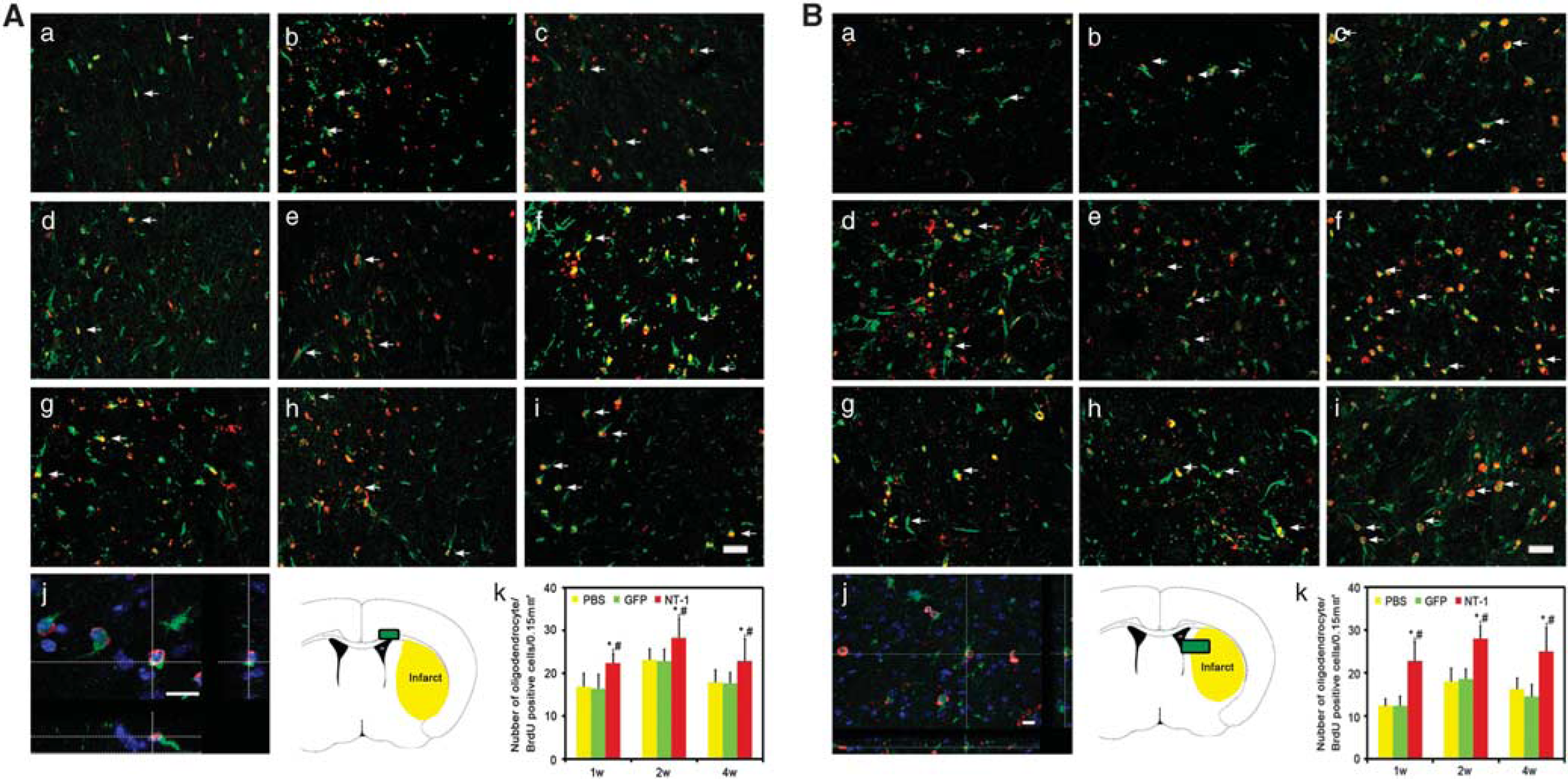
New matured oligodendrocytes in corpus callosum and striatum. Double-labeled BrdU and oligodendrocyte-specific protein (OSP) immunostaining was performed to demonstrate the newly matured oligodendrocyte. Photomicrographs showed BrdU (red)- and OSP (green)-positive staining in the corpus callosum (
Netrin-1 Overexpression Promoted White Matter Integrity
Myelin basic protein immunostaining was used to detect the white matter morphologic changes. After tMCAO, many small cavities in the ipsilateral corpus callosum were detected, whereas white matter maintained intact morphology in the contralateral hemisphere (Figure 5Aj). In the striatum, the injured axons collapsed and neurofilaments broke down in the ipsilateral hemisphere, whereas axons and fibers presented integrity structure in contralateral hemisphere (Figure 5Bj). As an indicator of white matter integrity, the ratio of MBP intensity of the ipsilateral over the contralateral hemispheres in
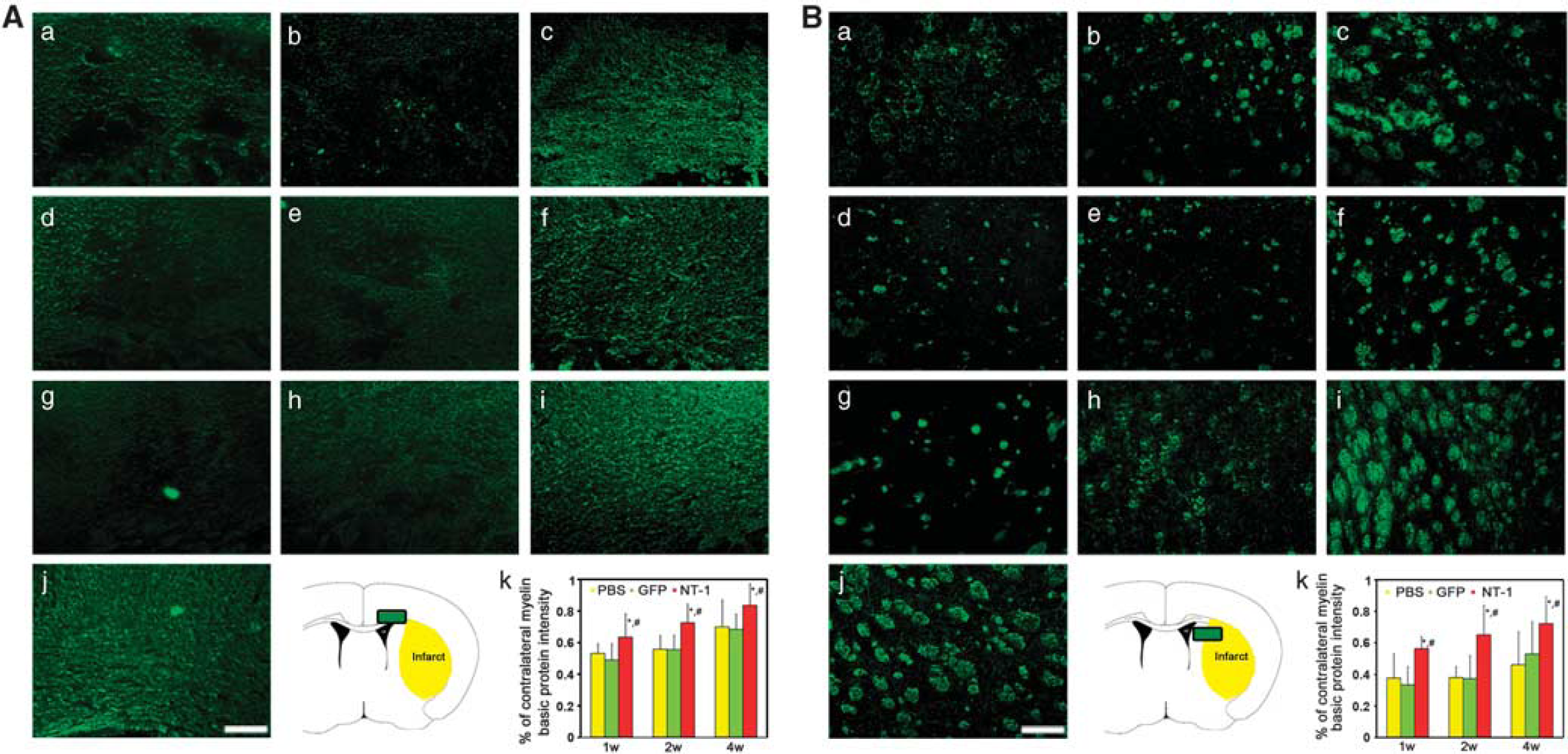
The morphology of myelined fibers in corpus callosum and striatum. The myelin basic protein (MBP) was stained in corpus callosum and striatum to detect the white matter injuries. Photomicrographs showed MBP-positive fibers in the corpus callosum (
Effects of Netrin-1 on White Matter were Dependent on DCC and UNC5H2
The effects of NT-1 on white matter were dependent on its binding receptors. Immunostaining results revealed that both DCC and UNC5H2 were expressed in the cytoplasm of OPCs (Figures 6A and 6B). In contrast, UNC5H2 was expressed on the MBP-positive fibers, whereas DCC receptor was absent (Figures 6C and 6D). These results implied that NT-1 had different roles in oligodendrogenesis.
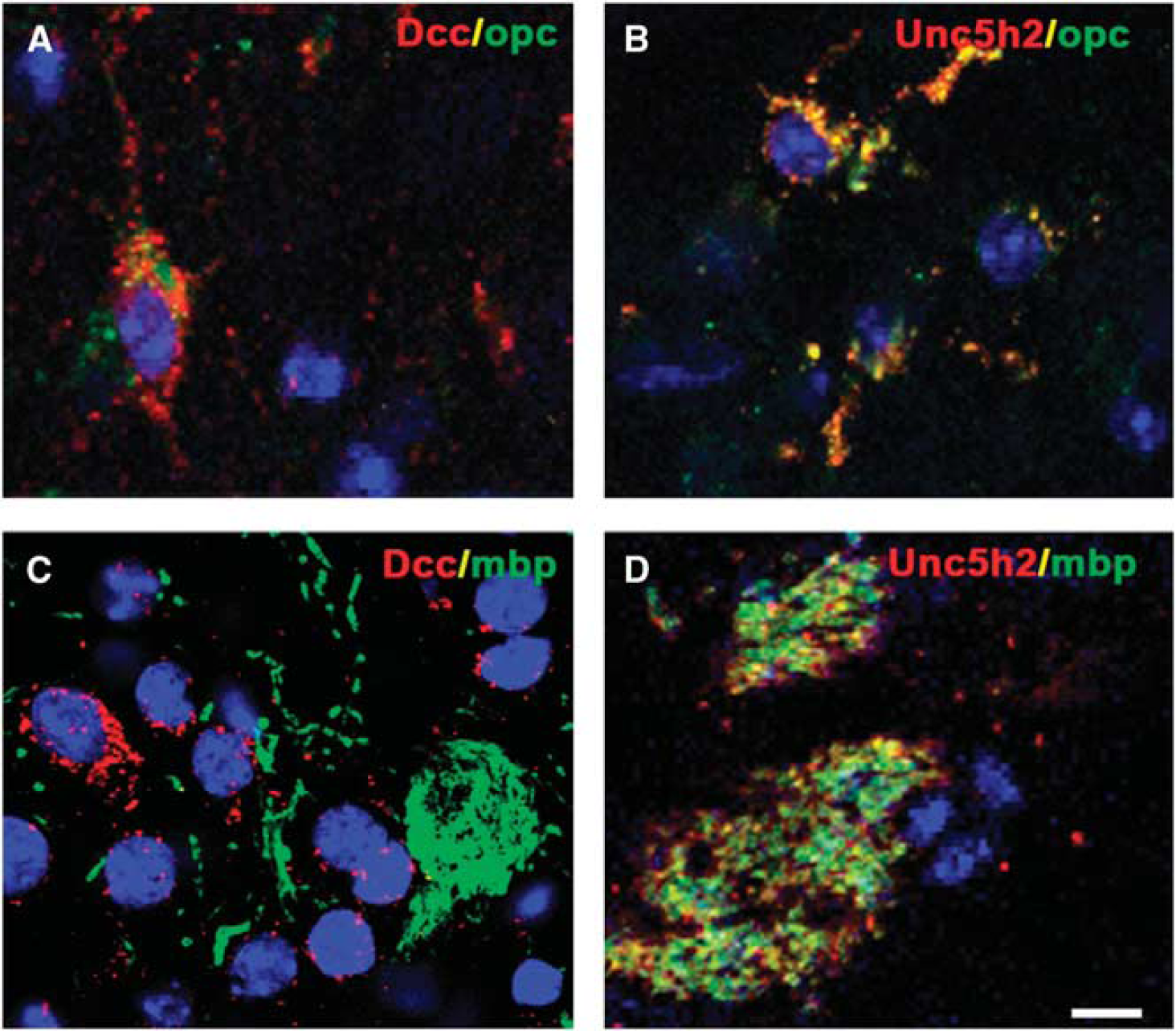
Expression of Netrin-1 (NT-1) receptors in white matter. Two receptors of NT-1 were detected by immunohistochemistry, both of deleted in colorectal carcinoma (DCC) (
DISCUSSION
Oligodendrogenesis is involved in brain tissue repairing and remodeling after ischemic brain injury.22, 23 Promoting white matter remodeling via pharmacological intervention or cell-based therapy improved motor function recovery.24, 25 In the present work, we studied the effect of NT-1 on white matter remodeling in tMCAO mice. We found that ischemia-induced motor function damage was greatly attenuated in
The effects of NT-1 on white matter development have been widely investigated. Netrin-1 is secreted by neurons and oligodendrocytes in the central canal, which conduces to generating an appropriate amount of oligodendrocytes by promoting OPC proliferation. 10 Netrin-1 also supports oligodendrocytes development by promoting OPC migration. 11 On the axon myelination process, NT-1 increases the branching of oligodendrocyte process. 27 In the peripheral nervous system, NT-1 promotes proliferation of Schwann cells, which possess the same function as oligodendrocytes in the central nervous system. Under ischemic conditions, NT-1 could be re-activated, suggesting that it may be a target for ischemic stroke treatment. 28 In the acute phase of ischemia, NT-1 reduces the infarct volume by inhibiting apoptosis, and improves synapse plasticity by changing the long-term potential.29, 30 Our previous study showed that NT-1 overexpression improved neurobehavioral performance by promoting focal angiogenesis and reduced infarct volume in a long-term study. Our current data suggest that NT-1 overexpression also restores myelination by increasing the number of proliferated OPCs in both the corpus callosum and striatum. This effect is consistent with the role of NT-1 in the developmental progress. Although NT-1 mainly acts as a guidance factor during both development and adulthood, it also increases the proliferation of renal proximal tubular epithelial cells as well as endothelial cells.31, 32 Consistent with NT-1 on white matter development, we demonstrated that NT-1 also preserves the similar function during ischemia by enhancing OPC proliferation and participating in white matter repairing and remodeling, which may contribute to improvement in neurobehavioral outcomes after ischemic brain injury.
Function of NT-1 relies on the combined receptors in various systems. Most broadly studied receptors of NT-1 are DCC and UNC5H2, but their functions in brain injury are still unknown, especially in the white matter system. 7 Both DCC and UNC5H2 are involved in the development of oligodendrocyte. 33 Netrin-1 binding with DCC displays both attraction and repelling effect whereas NT-1 binding with UNC5H2 mainly showed repelling role. 34 The proliferation of Schwann cells also requires UNC5H2 participation. 35 We found that both DCC and UNC5H2 were expressed on the cytoplasm of OPCs, implying that these two receptors were probably associated with the proliferative effects of NT-1 on OPCs after ischemic stroke. In contrast, immunohistochemistry showed that only UNC5H2 was detected on the MBP-positive fibers, indicating that UNC5H2 may be relevant with forming the sheath of axons and maintaining white matter integrity. Currently, DCC or UNC5H2 antagonists are commercially unavailable. Antibodies can be used to block DCC or UNC5H2;36, 12 however, the BBB limits accessibility of antibody into the brain. Although we demonstrated the neurobehavioral outcome improved and oligodendrocytes were increased in AAV-NT-1 transduced mice; future study is needed when proper blockers are developed for these receptors.
Our results demonstrated that NT-1 overexpression also promoted long-time recovery after ischemic stroke by improving oligodendrogenesis and white matter repairing. As
Footnotes
The authors declare no conflict of interest.