
Abstract
We present here an initial characterization of ATP binding cassette (ABC) transporter function and regulation at the blood–spinal cord barrier. We isolated capillaries from rat spinal cords and studied transport function using a confocal microscopy-based assay and protein expression using western blots. These capillaries exhibited transport function and protein expression of P-glycoprotein (Abcb1), multidrug resistance protein 2 (Mrp2, Abcc2), and breast cancer-related protein (Bcrp, Abcg2). Exposing isolated capillaries to dioxin (activates aryl hydrocarbon receptor) increased transport mediated by all three transporters. Brain and spinal cord capillaries from dioxin-dosed rats exhibited increased P-glycoprotein-mediated transport and increased protein expression for all three ABC transporters. These findings indicate similar ABC transporter expression, function, and regulation at the blood–spinal cord and blood–brain barriers.
Introduction
Multiple barriers separate the central nervous system from the periphery. Of these, the blood–brain barrier, which resides within the capillary endothelium, is the most studied and thus the most fully characterized (Abbott et al, 2010). This barrier is characterized by low paracellular permeability and the expression of specialized plasma membrane transport proteins and receptors, which together limit and regulate the exchange of solutes between the brain and the periphery. Certain members of the ATP binding cassette (ABC) family of transporters, e.g., P-glycoprotein (Abcb1), breast cancer-related protein (Bcrp, Abcg2), and multidrug resistance-associated protein (Mrp2, Abcc2), are substantial contributors to barrier selectivity. These plasma membrane proteins are ATP-driven efflux pumps that face the vascular space and limit brain entry of foreign chemicals (xenobiotics), including neurotoxicants and therapeutic drugs (Miller, 2010).
A morphologically similar, capillary-based barrier, the blood–spinal cord barrier, separates the spinal cord from the periphery (Bartanusz et al, 2011). As in brain capillaries, spinal cord capillary endothelial cells express multiple tight junctional proteins and adherens junction-associated proteins (Ge and Pachter, 2006). Low tight junctional permeability of the blood–spinal cord barrier presents a substantial physical barrier to macromolecules (Bartanusz et al, 2011). In contrast, little is known about ABC transporter function and regulation at the blood–spinal cord barrier. As in brain capillaries, P-glycoprotein is localized to the luminal plasma membrane of the spinal cord capillary endothelial cells (Sugawara et al, 1990); inhibiting P-glycoprotein
Here, we present an initial characterization of ABC transporter function and regulation at the rat blood–spinal cord barrier. In isolated capillaries, we show transport activity and protein expression of P-glycoprotein, Bcrp, and Mrp2. We show that activity of these transporters increased when capillaries were exposed to subnanomolar concentrations of 3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), a persistent environmental toxicant and high affinity ligand for the arylhydrocarbon receptor (AhR). P-glycoprotein activity and P-glycoprotein, Bcrp and Mrp2 protein expression also increased in TCDD-dosed rats. These findings define one mechanism by which expression of three ABC transporters is regulated at the blood–spinal cord barrier.
Materials and methods
Materials
[
Animals
All experiments were performed in compliance with NIH animal care and use guidelines (Guide for the Care and Use of Laboratory Animals, National Research Council) and approved by the NIEHS Animal Care and Use Committee (ARRIVE Guidelines). Male, 6 to 9 months old, retired breeder Sprague-Dawley rats and 12-week-old male, wild-type (FVB) and P-glycoprotein-null (FVB.129P2-
For
Capillary Isolation and Transport Assay
A detailed procedure for brain capillary isolation was described previously (Hartz et al, 2004). Briefly, for each preparation, white matter, meninges, midbrain, choroid plexus, blood vessels and olfactory lobes were removed from the 5 to 10 brains under a dissecting microscope and brain tissue homogenized. Tissue was kept in cold phosphate-buffered saline (PBS) (2.7 mmol/L KCl, 1.5 mmol/L KH2PO4, 136.9 mmol/L NaCl, 8.1 mmol/L Na2HPO4, 1 mmol/L CaCl2, 0.5 mmol/L MgCl2, 5 mmol/L
Confocal microscopy-based transport assays with isolated brain capillaries have been described previously (Hartz et al, 2004); an identical procedure was used for spinal cord capillaries. All experiments were performed at room temperature in coverslip-bottomed imaging chambers filled with PBS. In general, capillaries were exposed for 3 hours to nuclear receptor ligands without or with additional inhibitors. Fluorescent substrates, NBD-CSA for P-glycoprotein, TR for Mrp2, and BODIPY-prazosin for Bcrp, were added to each chamber and luminal substrate accumulation was assessed 1 hour later. In some experiments, specific transport inhibitors were included in the incubation medium. To acquire images, the chamber containing the capillaries was mounted on the stage of a Zeiss Model 510 inverted confocal laser scanning microscope (Thornwood, NY, USA) and imaged through a × 40 water-immersion objective (numeric aperture=1.2) using a 488-nm laser line for NBD-CSA and BODIPY-prazosin or a 543-nm laser line for TR. Images were saved to disk and luminal fluorescence was subsequently quantitated by Image J software (NIH, Bethesda, MD, USA). Data are presented as arbitrary fluorescence units from 8-bit images.
Western Blots
Membranes were isolated from control and ligand-exposed capillaries as described previously (Hartz et al, 2004). An aliquot of the membrane protein was mixed with NuPAGE 4 × sample buffer (Invitrogen), loaded onto 4% to 12% Bis-Tris NuPAGE gel, electrophoresed, and then transferred onto an Immobilon-FL membrane (Millipore, Bedford, MA, USA). The membrane was blocked with Odyssey Blocking Buffer (Li-Cor Biosciences, Lincoln, NE, USA) at room temperature for 1 hour, and then immunoblotted with antibodies against P-glycoprotein, Mrp2, or Bcrp. The membrane was stained with corresponding goat anti-rabbit or goat anti-mouse fluorescence dyes IRDye 680 (or IRDye 800) in PBS with 0.1% Tween-20 at room temperature for 45 minutes and then imaged using an Li-Cor Biosciences Odyssey Infrared Imaging System.
Electrophoretic Mobility Shift Assay
Nuclear protein extract was isolated from control and TCDD-induced spinal cord capillaries using an NE-PER kit (Pierce, Rockford, IL, USA), following manufacturer's instructions. Complementary DNA oligonucleotides 5′-GAT CCG GCT CTT CTC ACG CAA CTC CGA GCT CA-3′ and 5′-TGA GCT CGG AGT TGC GTG AGA AGA GCC GGA TC-3′ (dioxin response element recognition sequence highlighted) were end labeled with IRDye700 fluorescent dyes by IDT (Coralville, IA, USA) and annealed at 100°C for 5 minutes. The EMSA (Electrophoretic Mobility Shift Assay) samples were prepared using Odyssey Infrared EMSA Buffer Kit (Li-Cor Biosciences), with 5 μg of nuclear protein extract used in each AhR/dioxin response element binding reaction. The binding reaction was incubated at the room temperature for 20 minutes and the DNA-protein complexes were resolved on a precast, 6% native polyacrylamide gel in 0.5 × TBE buffer. The gel was removed from the electrophoresis unit and directly imaged on the Odyssey Infrared Imaging System.
Statistical Analyses
Data are expressed as mean±standard error of the mean. Differences between treatment means were considered significant when
Results
We established previously an assay to measure efflux transporter activity in living, isolated brain capillaries (Hartz et al, 2004). It is based on the use of confocal microscopy and digital image analysis to measure the specific accumulation of fluorescent substrates (NBD-CSA for P-glycoprotein, BODIPY-prazosin for Bcrp, and TR for Mrp2) within capillary lumens (Bauer et al, 2008; Hartz et al, 2004; Wang et al, 2010). In those studies, we also validated selective inhibitors for each transporter: PSC833 for P-glycoprotein, Ko143 for Bcrp, and MK571 for Mrp2. In initial experiments with spinal cord capillaries, we found rapid luminal accumulation of all three fluorescent substrates, with steady state established within 45 minutes (not shown). Figure 1A shows representative confocal images of rat spinal cord capillaries after 60 minutes incubation in medium containing 2
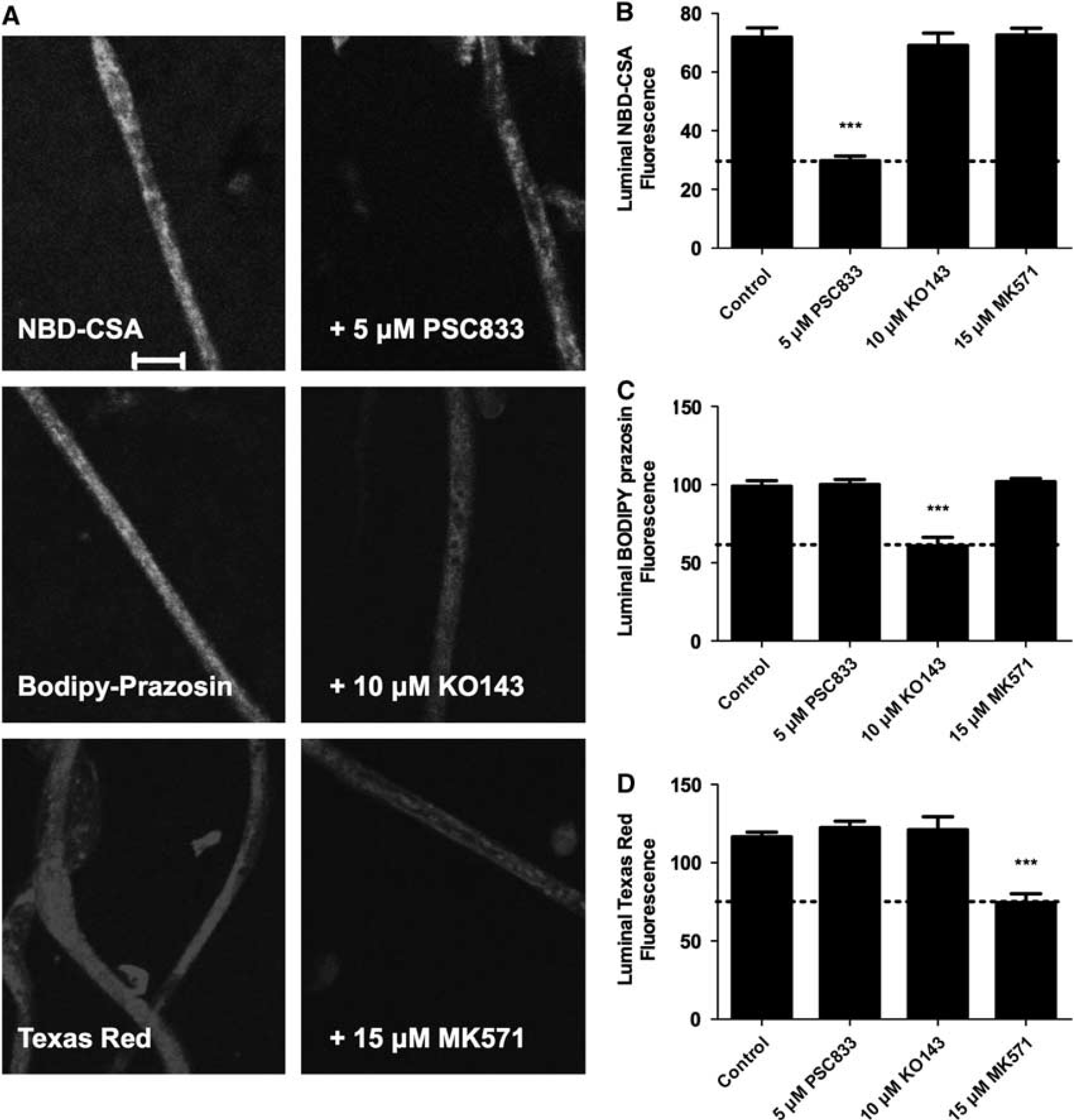
ATP binding cassette (ABC) transporter function at the rat blood–spinal cord barrier. (
To further validate the assay, we measured luminal accumulation of NBD-CSA in brain and spinal cord capillaries from wild-type and P-glycoprotein-null mice. For both tissues, luminal NBD-CSA accumulation in capillaries from wild-type mice was over twice that in capillaries from P-glycoprotein-null mice (Figure 2). Moreover, in both tissues, luminal NBD-CSA accumulation in capillaries from P-glycoprotein-null mice was same as in capillaries from wild-type mice that had been exposed to PSC833 (Figure 2). Note that previous studies indicated that luminal fluorescence remaining after PSC833 inhibition of transport represents nonspecific accumulation, likely from diffusive entry plus binding to cellular elements (Hartz et al, 2004). Together, the experiments in Figures 1 and 2 define assays that measure transport activity of three ABC transporters in rat spinal cord capillaries. For the following experiments concerned with transporter regulation, we use steady-state, specific (inhibitable) luminal accumulation of the fluorescent substrates as a measure of transporter activity in the isolated capillaries.
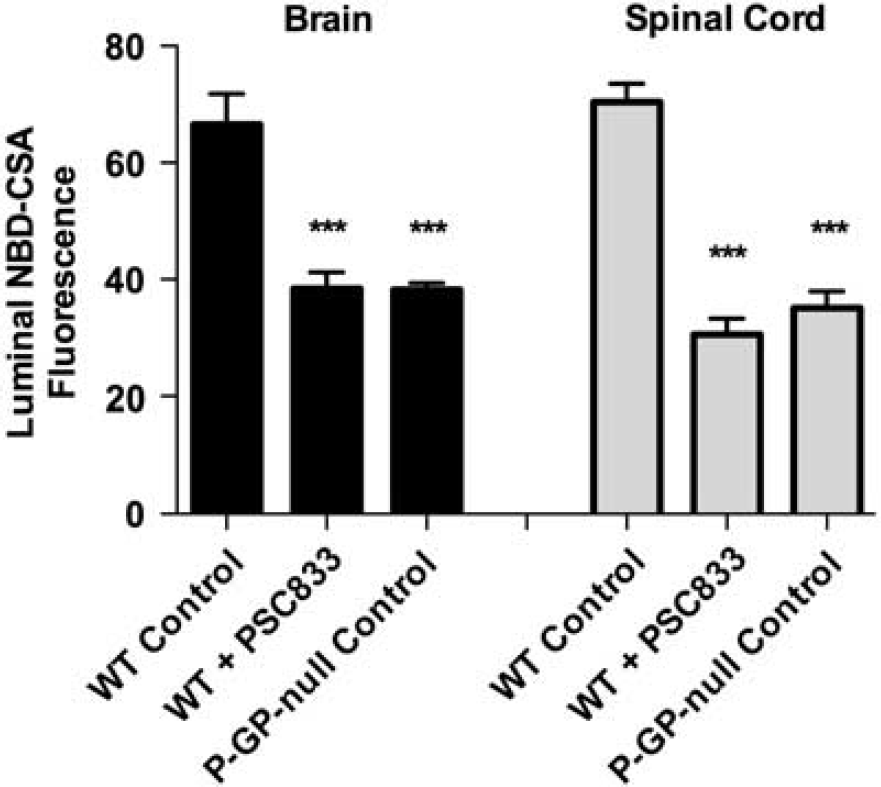
NBD-CSA transport in brain and spinal cord capillaries from wild-type (WT) and P-glycoprotein (P-GP)-null mice. Capillaries were incubated for 60 minutes in medium with 2
We recently reported that transport activity and protein expression of ABC transporters at the blood–brain barrier are upregulated by ligands that activate the AhR, e.g., the dioxin, TCDD (Wang et al, 2011). Exposing spinal cord capillaries to 0.05 to 0.5 nmol/L TCDD increased specific P-glycoprotein transport activity in a concentration-dependent manner (Figure 3A). Pretreating the capillaries with actinomycin D or cycloheximide abolished the increase in transport activity caused by TCDD exposure (Figure 3B). Exposing the capillaries to AhR antagonists, resveratrol or
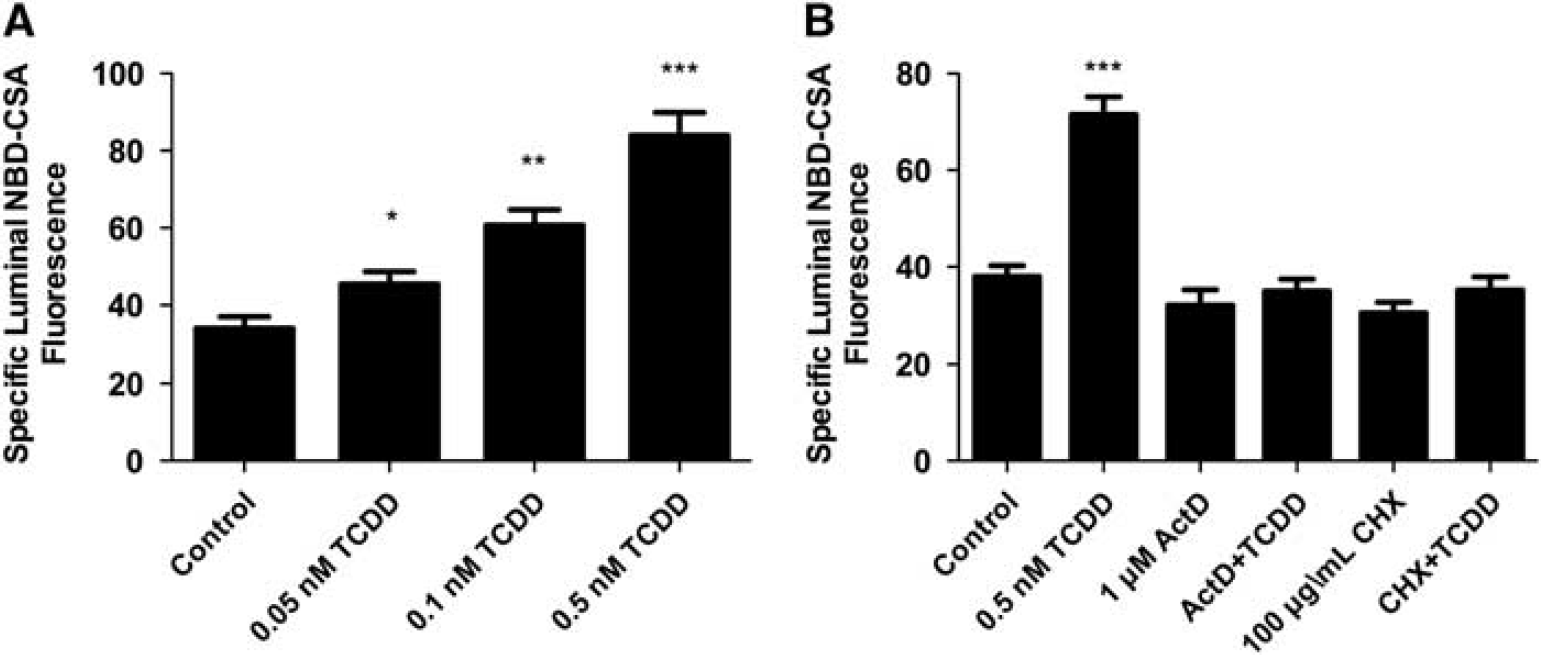
TCDD exposure increases P-glycoprotein-mediated transport. (
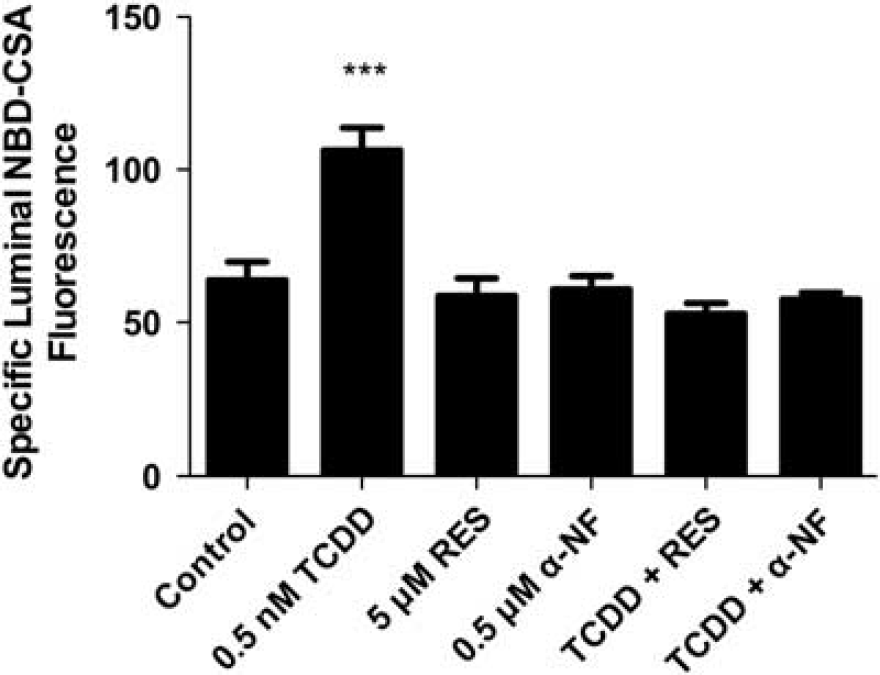
TCDD effects on P-glycoprotein-mediated transport are blocked by arylhydrocarbon receptor (AhR) antagonists, 0.5
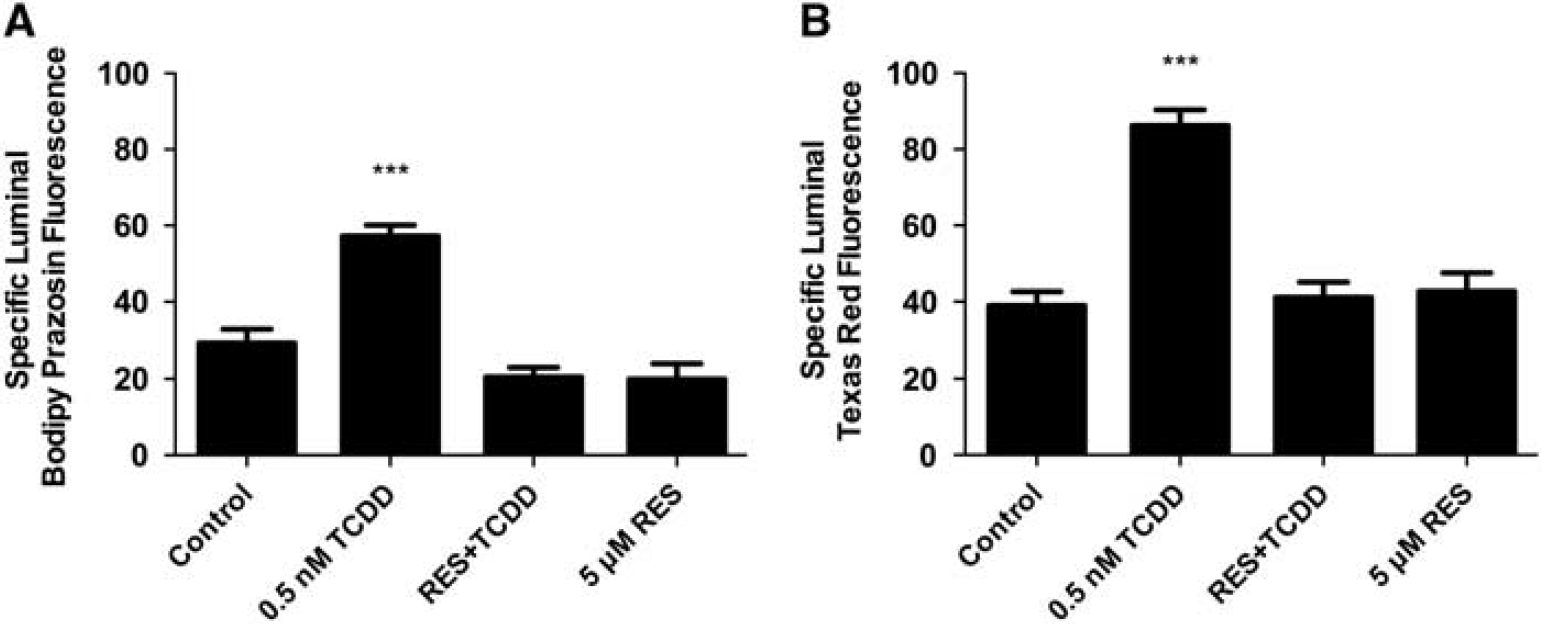
TCDD exposure increases Bcrp- (
In rats and mice, activation of the pregnane-X-receptor (PXR) and the constitutive adrostane receptor (CAR) increases ABC transporter expression at the blood–brain barrier (Miller, 2010). In initial experiments we exposed spinal cord capillaries to pregnenolone 16
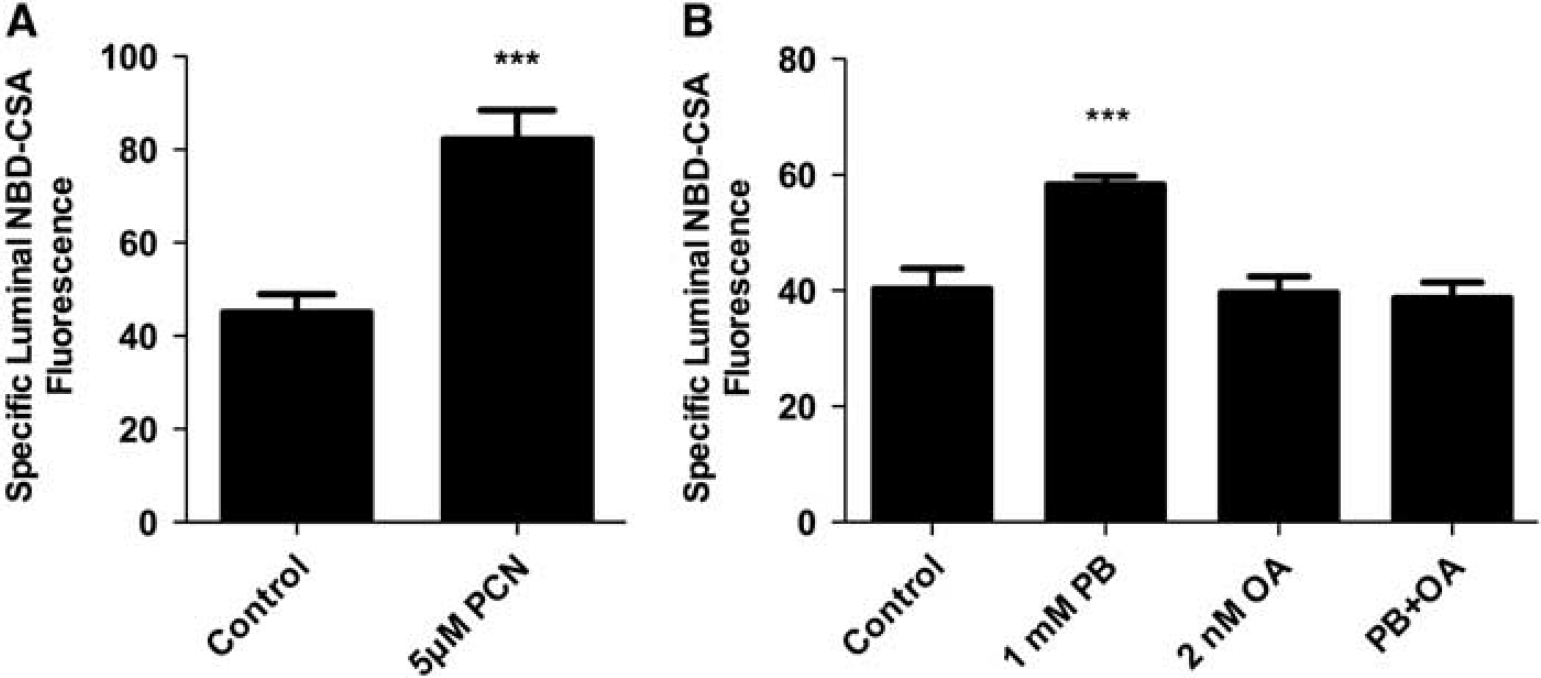
Exposing rat spinal cord capillaries to 5
To determine the effects of TCDD exposure
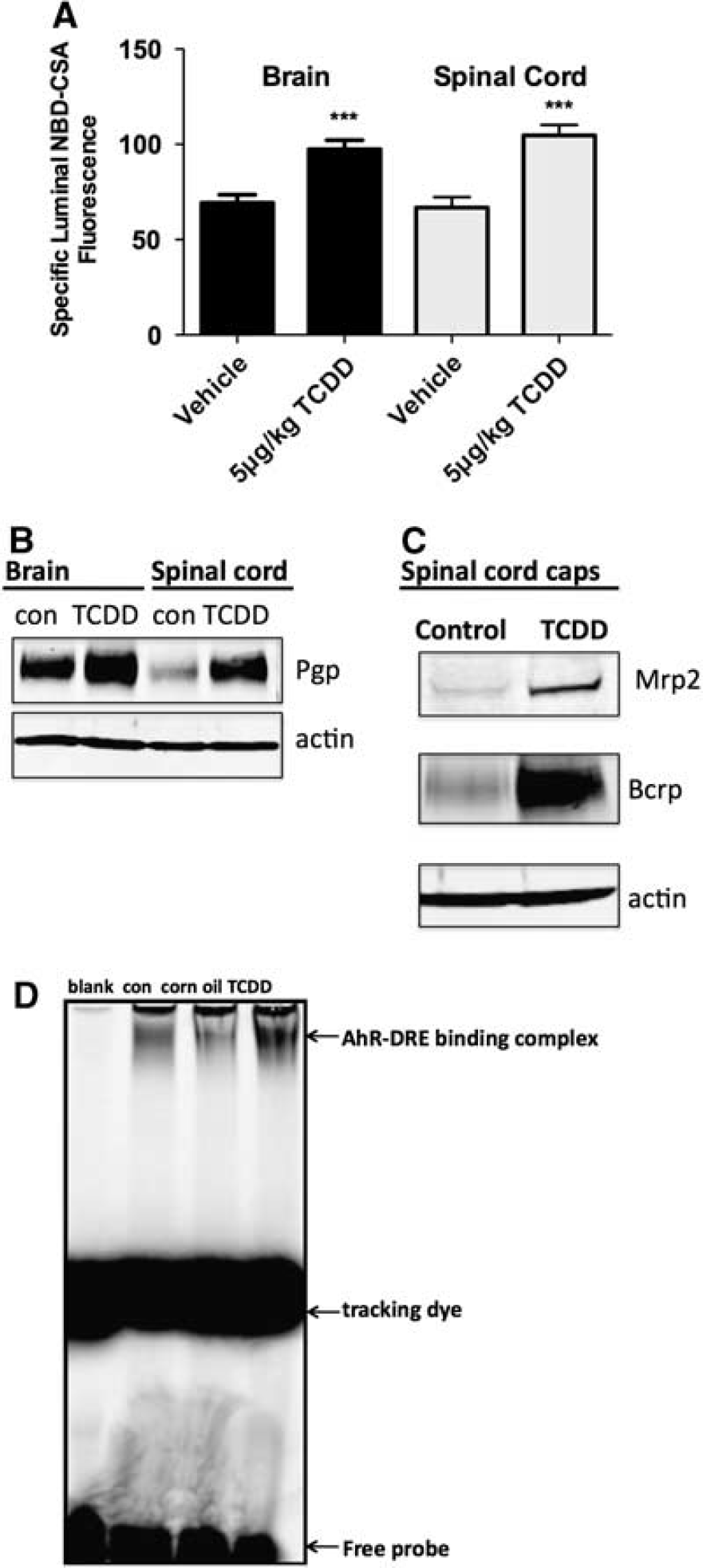
ATP binding cassette (ABC) transporter function and expression in spinal cord and brain capillaries from control and TCDD-dosed rats (5
Discussion
Our understanding of the molecular basis of blood–spinal cord barrier function and dysfunction lags substantially behind that of the blood–brain barrier. This is especially true for the ABC transporters that provide an important, selective element of the barrier. In the present study, we show that specific ABC transporter activity can be measured in isolated spinal cord capillaries using the same confocal microscopy-based assays developed previously for brain capillaries (Miller et al, 2000). We confirm the presence of P-glycoprotein on capillary membranes and show its transport function in the isolated capillaries from rats and mice. As shown previously in mouse (Ge and Pachter, 2006), at the protein level, P-glycoprotein expression appears to be decrease in rat spinal cord capillaries than in rat brain capillaries. Importantly, we show for the first time at the blood–spinal cord barrier transport function and protein expression for Bcrp and Mrp2. Although we have not yet immunostained spinal cord capillaries for Mrp2 and Bcrp, the present transport data are entirely consistent with luminal localization of both transporters. Our unpublished experiments with spinal cord capillaries isolated from multiple mouse strains show similar results for ABC transporter function.
Certain ligand-activated nuclear receptors upregulate genes that code for proteins involved in phase 1 and phase 2 xenobiotic metabolism and xenobiotic excretion. Although most work in this area has been focused on peripheral barrier and excretory tissues, e.g., liver, recent studies show similar responses at the blood–brain barrier for PXR, CAR, and AhR (Miller, 2010; Wang et al, 2011). Consider AhR, which is activated by environmental toxicants, e.g., TCDD, and drugs.
Finally, blood–brain barrier, P-glycoprotein and Bcrp, substantially determine the extent to which many small molecule drugs enter the brain (Hartz et al, 2010). Of these transporters, P-glycoprotein handles a wider range of drugs, and is thus considered as a major obstacle to drug delivery to the brain. Available data indicate that blood–brain barrier P-glycoprotein expression is altered in central nervous system disease, e.g., increased in epilepsy and stroke and reduced in Alzheimer's disease (Zlokovic, 2008, 2011).
In this regard, altered blood–spinal cord barrier function is increasingly recognized as one consequence of traumatic injury, e.g., spinal cord and peripheral nerve injury, as well as neurodegenerative disease, e.g., amyotrophic lateral sclerosis. Both tight junction permeability and transporter expression/function are implicated. On the one hand, altered barrier function can disturb homeostatic mechanisms that depend on the composition of spinal cord interstitial fluid. On the other hand, altered expression of drug efflux pumps can affect pharmacotherapy. Consider amyotrophic lateral sclerosis, recent studies show altered blood–spinal cord barrier function before symptoms become apparent both in an animal model of the disease (transgenic SOD1 mouse) and in patients with sporadic and familial amyotrophic lateral sclerosis (Zlokovic, 2011). Moreover, the SOD1 mouse exhibits increased P-glycoprotein expression at the blood–spinal cord barrier (Boston-Howes et al, 2008; Milane et al, 2010), suggesting reduced ability to deliver certain drugs to the spinal cord. How this disease leads to increased transporter expression is not known. We posit that understanding the molecular basis for drug transport at the blood–spinal cord barrier and its regulation will provide strategies to improve delivery of therapeutic drugs.
Footnotes
Disclosure/conflict of interest
The authors declare no conflict of interest.