
Abstract
The brainstem of the adult rat is relatively resistant to spreading depolarization (SD) but after enhancement of excitability SD can be evoked by local application of KCl. In the present experiments, we observed that the enhanced excitability even triggers prolonged periods of repetitive depolarizations (RDs), which elicit significant cardiovascular changes. In contrast to KCl-evoked SDs with amplitudes of ∼24 mV and spreading velocity of 4 mm/min, spontaneous RDs had amplitudes of 7 to 12 mV, propagated up to 30 times faster than KCl-evoked SDs, and depolarized larger brainstem areas including the contralateral side. Similarly as SD, RDs depended on glutamatergic neurotransmission and were blocked by MK-801 or by the calcium channel blocker agatoxin. They depended on sodium channels and were blocked by tetrodotoxin. Functionally, the invasion of RDs into the spinal trigeminal and other nuclei evoked bursts of action potentials, indicating that specific neuronal systems are affected. In fact, during episodes of RDs the blood pressure and the local blood flow at the surface of the brainstem and the cortex increased substantially. Brainstem RDs did not propagate into the cerebral cortex. We propose to consider brainstem RPs as a pathophysiological mechanism whose significance for brainstem disease states should be further explored.
Introduction
Cortical spreading depolarization (SD) is associated with a propagating negative direct current (DC) potential shift in the extracellular space of the cerebral cortex. The SD is caused by a massive depolarization of nerve cells, which release large amounts of potassium ions. The potassium ions induce an inward current across glial membranes, which lead to a depolarization of glial cells. The neuronal depolarization leads to a subsequent block of synaptic transmission producing the ‘spreading depression of electroencephalographic activity’. Leão (1944) for the first time described this transient flattening of the electroencephalogram in the cerebral cortex of the rabbit, and a few years later he identified the spreading depression as a propagating depolarization wave (Leão, 1947). The neuronal and glial depolarization is accompanied by redistribution of ions, by water influx into cells and by shrinkage of the extracellular volume (Somjen 2001, 2005; Nicholson, 1993; Gardner-Medwin, 1981; Kraig and Nicholson, 1978; Vyskočil et al, 1972). Depolarization phenomena in neurons are complex. Dendrites may be more important for SDs than somata because—as shown in hippocampal neurons—zonal depolarization and intense membrane shunts form longitudinal depolarization gradients (Canals et al, 2005). At present, nomenclature is changing from Leão's historical term ‘spreading depression’ to ‘spreading depolarization’ that is closer correlated with the electrophysiological mechanisms (Dreier, 2011), and we used the new term also with its abbreviation SD in this paper.
While SD easily can be elicited in cerebral cortex, the brainstem is more resistant to SD (Bureš et al, 1974). However, we showed that SD can be also evoked by microinjection of KCl in the immature rat brainstem up to postnatal day 14 after conditioning with acetate, but with maturation the brainstem becomes resistant to SD (Richter et al, 2003, 2005). In adult rat brainstem, KCl-evoked SD can be elicited by a stronger conditioning, i.e., with superfusion of a solution containing acetate, the potassium channel blocker tetraethylammonium chloride (TEA), and 10 mmol/L KCl. KCl-evoked SDs also depolarize centers of cardiovascular control in the brainstem, as indicated by transient increases in heart rate (HR), in systemic arterial blood pressure (ABP) and regional blood flow in the brainstem (Richter et al, 2008).
For a long time, SD in human cerebral cortex was related to the pathogenesis of migraine aura, because SD closely correlates with the propagation of scintillating scotoma across the visual field in the aura period (
All forms of SDs in the cortex and SDs in the brainstem appear as single events which are clearly separated by intervals and which occur only when a trigger, e.g., local application of KCl is applied. In studies on the SD in the brainstem, we noted occasionally that besides the SD evoked by KCl repetitive depolarizations (RDs) at shorter intervals may occur when the brainstem is conditioned by superfusion. Therefore, in the present study, we addressed specifically the occurrence, nature, and mechanisms of such RDs. We quantified their frequency, their amplitudes, and their mechanisms and asked whether they differ from SDs evoked by local KCl application. Furthermore, we investigated whether RDs evoke action potentials in brainstem nuclei and whether they induce time-coupled changes of blood pressure, HR, and local blood flow in brainstem and cortex. We found that RDs occur in about two thirds of rats after conditioning the brainstem, and that they induce cardiovascular changes similarly as SDs evoked by KCl. The mechanisms of RDs show similarities to SDs but the former have significantly smaller amplitudes, and ‘spreading’ occurs at a much higher velocity.
Materials and methods
Animal Preparation
The present study was performed according to the Protection of Animals Act of the Federal Republic of Germany (Tierschutzgesetz der Bundesrepublik Deutschland) and was approved by the Thuringian State Office for Food Safety and Consumer Protection (Thüringer Landesamt für Lebensmittelsicherheit und Verbraucherschutz) (Reg. No. 02-040/06). During the experiment, the animals were treated in accordance with the declaration of Helsinki and the guiding principles in the care and use of animals. Thirty-five male Wistar rats (body weight 300 to 450 g, aged older than 90 days) were anesthetized with 100 mg/kg sodium thiopentone (Trapanal; Nycomed Deutschland GmbH, Konstanz, Germany) intraperitoneally; supplemental doses (20 mg/kg intraperitoneally) were given as necessary to maintain areflexia. Through the cannulated trachea, the animals breathed spontaneously during surgery. A catheter (diameter 0.5 × 0.9 mm, Braun Melsungen, Melsungen, Germany) was inserted into the right femoral artery. Changes of systemic ABP were measured with the pressure transducer P23Db (Statham Instruments, Hato Rey, Puerto Rico). Body temperature was kept at 37°C using a feedback-controlled temperature constanter (L/M-80; List, Darmstadt, Germany). Another catheter was introduced into the femoral vein and allowed to supply the animals with 5% glucose or Tyrode solution. At the end of the experiments, the animals were killed by an overdose of the anesthetic intravenously.
The head of the animal was fixed in a stereotaxic frame. The apical and parietal parts of the skull were exposed after a median incision. Over the left hemisphere, the dura mater was exposed in an area of 4 × 4 mm (2 mm posterior to bregma, 1 to 2 mm laterally from the midline) using a mini-drill under cooling with artificial cerebrospinal fluid (ACSF; in mmol/L: NaCl 138.4, KCl 3.0, CaCl2 1.3, MgCl2 0.5, NaH2PO4 0.5, urea 2.2, glucose 3.4, warmed to 37°C and equilibrated with 5% CO2 in O2), and kept moist with ACSF throughout preparation. The dura mater was opened. Incising the neck muscles and the ligamentum atlantooccipitale exposed the brainstem from the occipital bone to the first cervical vertebra. The dura mater and arachnoidea underneath were removed.
After the surgical preparation, the animals were paralyzed with pancuronium bromide (Pancuronium Organon; Organon GmbH, Oberschleißheim, Germany; 2 to 4 mg/kg per hour intravenously) and artificially ventilated with room air that was enriched with O2 (oxygen content 30% to 35%). Anesthesia by sodium thiopentone was maintained to the end of the experiment.
Recording of Direct Current Potentials
An Ag/AgCl reference electrode containing 2 mol/L KCl was placed on the nasal bone. Intracortical DC potentials were recorded using a glass micropipette filled with 150 mmol/L NaCl (tip diameter ∼5
All electrodes were connected by Ag/AgCl wires to a custom built 4-channel-high impedance amplifier (Meyer, Munich, Germany). The signals were stored on PC. The animals were grounded through an Ag/AgCl electrode below the back skin.
Recording of Neuronal Activity in the Brainstem and Measuring of Calcitonin Gene-Related Peptide (CGRP) Release
In a group of six animals, action potentials from either single or multiple neurons were recorded extracellularly in the caudal trigeminal nucleus with carbon fiber microelectrodes (tip diameter 15 to 20
To measure in these six animals the calcitonin gene-realted peptide (CGRP) release from nociceptive nerve endings in the dura mater the opening of the skull was surrounded with dental acrylic, but the dura mater was kept intact (no DC electrode was inserted into the gray matter) and the pool above the dura mater was filled with 100
Superfusion of the Brainstem and Conditioning for Spreading Depolarization
The brainstem was superfused with modified ACSF in which 100 mmol/L of the chloride (75%) was replaced by 100 mmol/L acetate. In all, 10 mmol/L TEA (Sigma, Seelze, Germany) and 10 mmol/L KCl were added to the acetate-ACSF (acetate-ACSF+TEA+KCl). This superfusate had successfully conditioned the adult rat brainstem for SD in previous experiments (Richter et al, 2008). The superfusion lasted for at least 1 hour and was only interrupted when 1 mol/L KCl (see above) was injected into the brainstem to test the elicitation of SD by KCl during a period of RD waves.
In five animals, we injected intraperitoneally 3 mg/kg MK-801 (Sigma) during a period of RD waves. In another six animals, we applied 50
At the end of the experiment, the animals were killed by an overdose of sodium thiopentone (100 mg per rat, intravenously) and the terminal negative shift of the DC potentials in the cerebral cortex and the brainstem was monitored.
Measurement of Cardiovascular Parameters and Regional Blood Flow
Measurements of regional blood flow in the brainstem and the cerebral cortex, recording of systemic ABP, and HR were performed in all animals. An electrocardiogram was recorded from standard limb leads using stainless steel needle electrodes. In a subset of 10 rats, local brain cortical or brainstem blood flow (laser-Doppler flowmetry) were continuously measured using a Laser Doppler Flowmeter (MBF3D; Moor Instruments, Devone, Exminster, UK) with two single fiber probes (fiber separation was 225
Data Acquisition and Evaluation
All data except blood gas and acid-base balance values were recorded on PC by using custom signal acquisition programs. The DC shifts were analyzed regarding amplitudes at the first time of appearance and during full-blown periods of depolarization waves. Time differences between the peaks at different electrode positions were examined. Propagation velocity of the depolarization waves was recorded by dividing the time interval between two DC maxima by the spatial distance between the two electrodes, assuming a linear propagation. For HR calculations, the R-R intervals were estimated using the R wave peak as the trigger. The reciprocal of the R-R interval series represented the instantaneous HR (in b.p.m.). To evaluate the changes in local blood flow, we established a baseline over 3 seconds before the beginning of a depolarization and related the increase to the baseline (in the Results, the net increase is given). The laser-Doppler flowmetry monitor displays blood flow readings in arbitrary units that do not allow for measurement of blood flow in terms of absolute values, but the method is valid in determining relative changes of blood flow during moderate flow increases (Fabricius and Lauritzen, 1996).
Data are reported as mean values±standard deviations. In each animal, a mean value before and during the event was calculated, the data present mean values calculated from the individual means. The changes of values within groups (before conditioning versus during conditioning; before depolarization versus during depolarization) were tested for significance with the Wilcoxon matched pairs signed rank test. Comparisons between different groups were performed by nonparametric analysis of variance (Kruskal–Wallis test with Dunn's multiple comparison test) or by a one-way analysis of variance. When required, a Bonferroni adjustment was performed to evaluate significant differences. Significance was accepted at
Results
Electrophysiological Characterization of the Repetitive Depolarization Waves
The conditioning of the brainstem with a special superfusate (75% of the chloride ions were replaced by sodium acetate+10 mmol/L KCl+10 mmol/L TEA) evoked in 24 out of 35 animals (68.6%) RD waves in the brainstem (Figure 1). The average time between starting the superfusion and the observation of the first depolarization wave was 43.3±29.9 minutes. In most of the animals, these depolarization waves occurred first as single events, and after another 15 to 30 minutes they occurred more regularly, and finally they appeared as series of RDs. Within such a series of RDs 6 to 74 DC shifts occurred, and a series of RDs had a mean duration of 30.9±20.6 minutes (minimum 9.4 minutes; maximum 106.7 minutes). The time interval between two subsequent RDs progressively shortened within a series of RDs (Figure 2). In contrast to the shortened time intervals, the temporal delay between RDs at the different electrodes did not significantly change. It was 2.9±5.4 seconds for the first one and 2.5±2.3 seconds within the series of depolarizations. In 15 out of the 24 animals, there was no temporal delay at all between the two recording electrodes (electrode distance in 12 rats 1 mm, in the other three rats 1.5, 3.0, or 5.0 mm), and only in 9 rats the temporal difference was longer than 1 second.
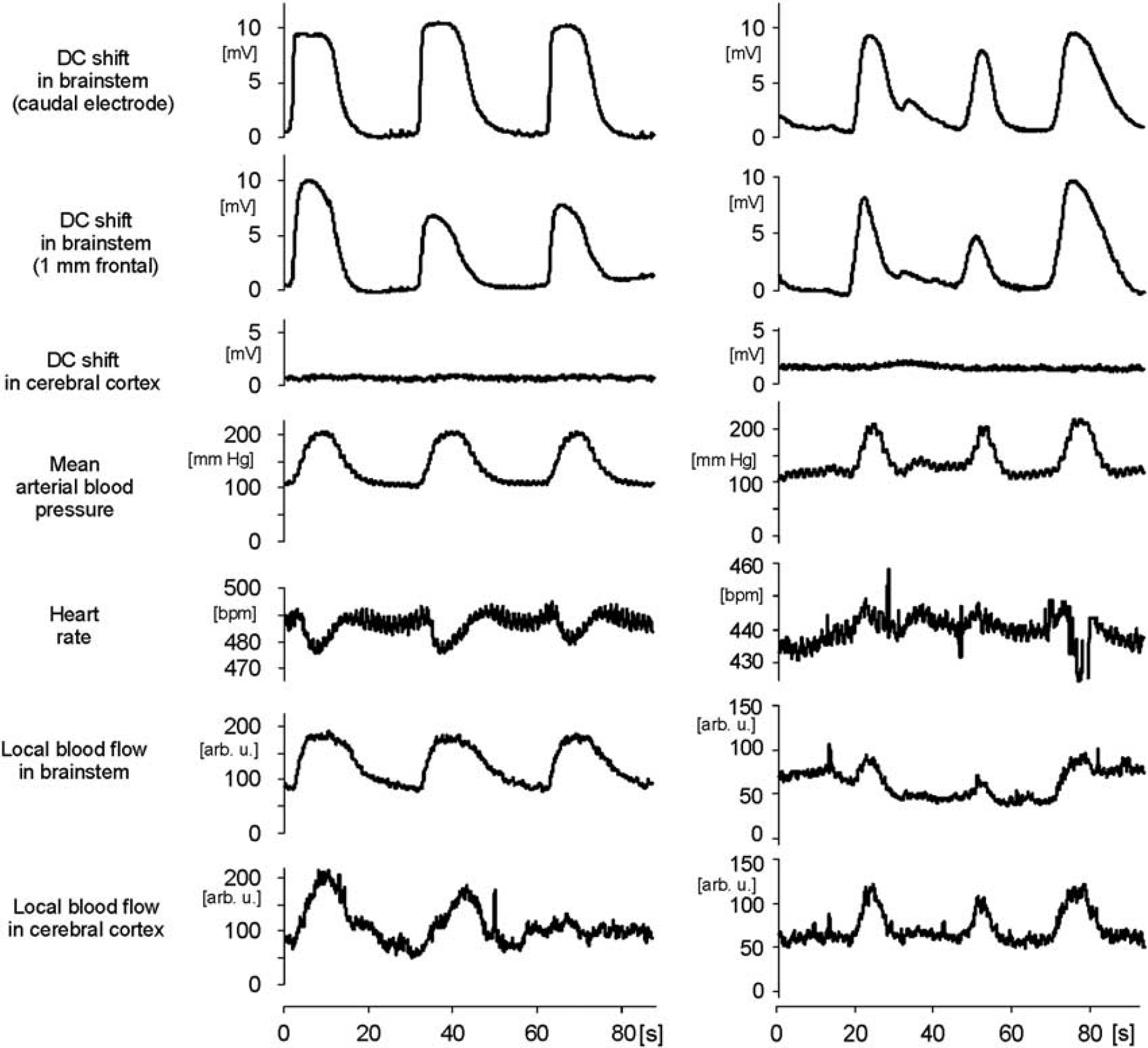
Repetitive depolarizations (RDs) in the brainstem and changes of cardiovascular parameters. Specimens show simultaneous recordings of direct current (DC) potentials in brainstem at two sites separated by 1 mm and in cerebral cortex, heart rate, systemic arterial blood pressure (ABP), and local blood flow in brainstem and cerebral cortex. The two columns depict rapidly recurring depolarizations in two different rats. Note the arrhythmia induced by the third depolarization in the right panel (vertical lines). Depolarizations in the brainstem never induced spreading depolarization (SD) in the cerebral cortex.
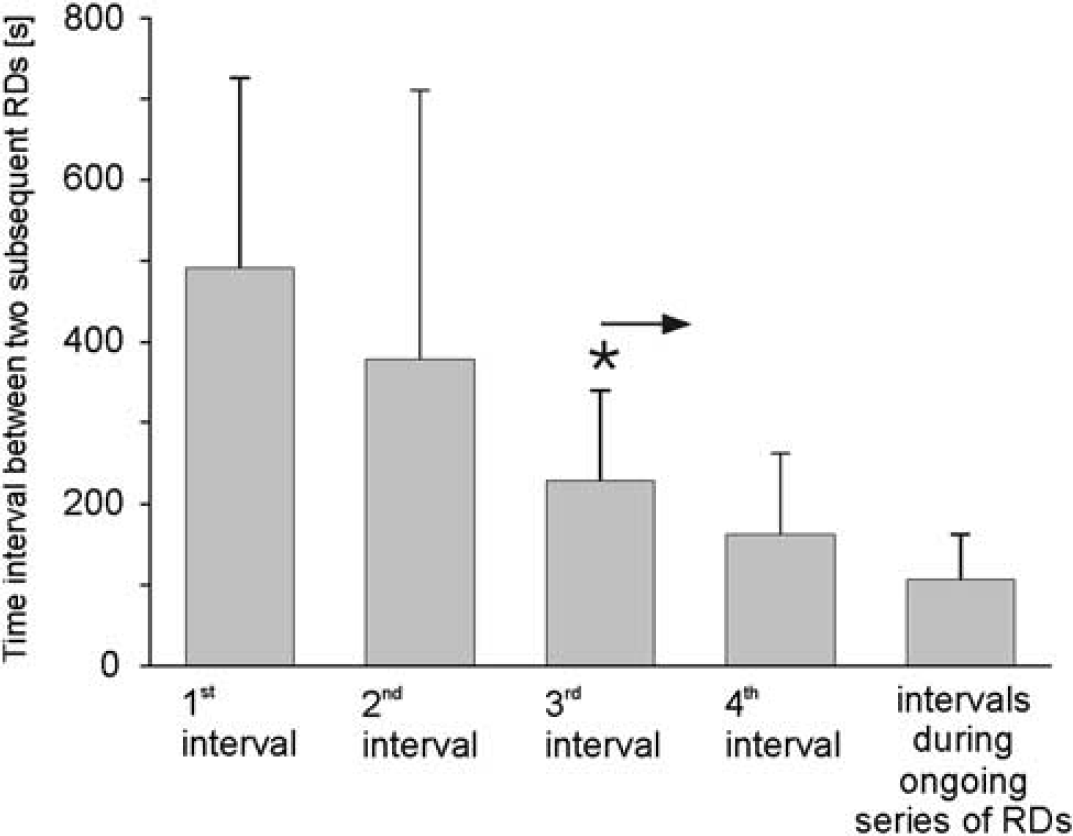
Shortening of time intervals between two subsequent repetitive depolarizations (RDs) in a sequence from the first, single depolarization to an ongoing series of RDs. Data show mean values from 24 animals with RDs. The asterisk with arrow indicates statistical significant differences to the first interval from the third depolarization on (Wilcoxon matched pairs signed rank test,
The mean DC amplitude was 7.7±4.0 mV (Figure 1), but in some cases DC amplitudes larger than 25 mV were observed (see also Figure 3B). The time interval of ∼2 seconds between the onsets of depolarization waves at two electrodes with a distance of 1 mm indicated a propagation velocity of ∼0.5 mm/s corresponding to 30 mm/min which is 10 times faster than in SD evoked by KCl. Overall, we observed 34 episodes with altogether 774 depolarizations in the trigeminal nucleus (Sp. V) area. In three of the rats, the RDs ended spontaneously; in all other animals either a washout of the conditioning superfusate or the application of a substance inferring with glutamatergic neurotransmission (see below) stopped the depolarizations.
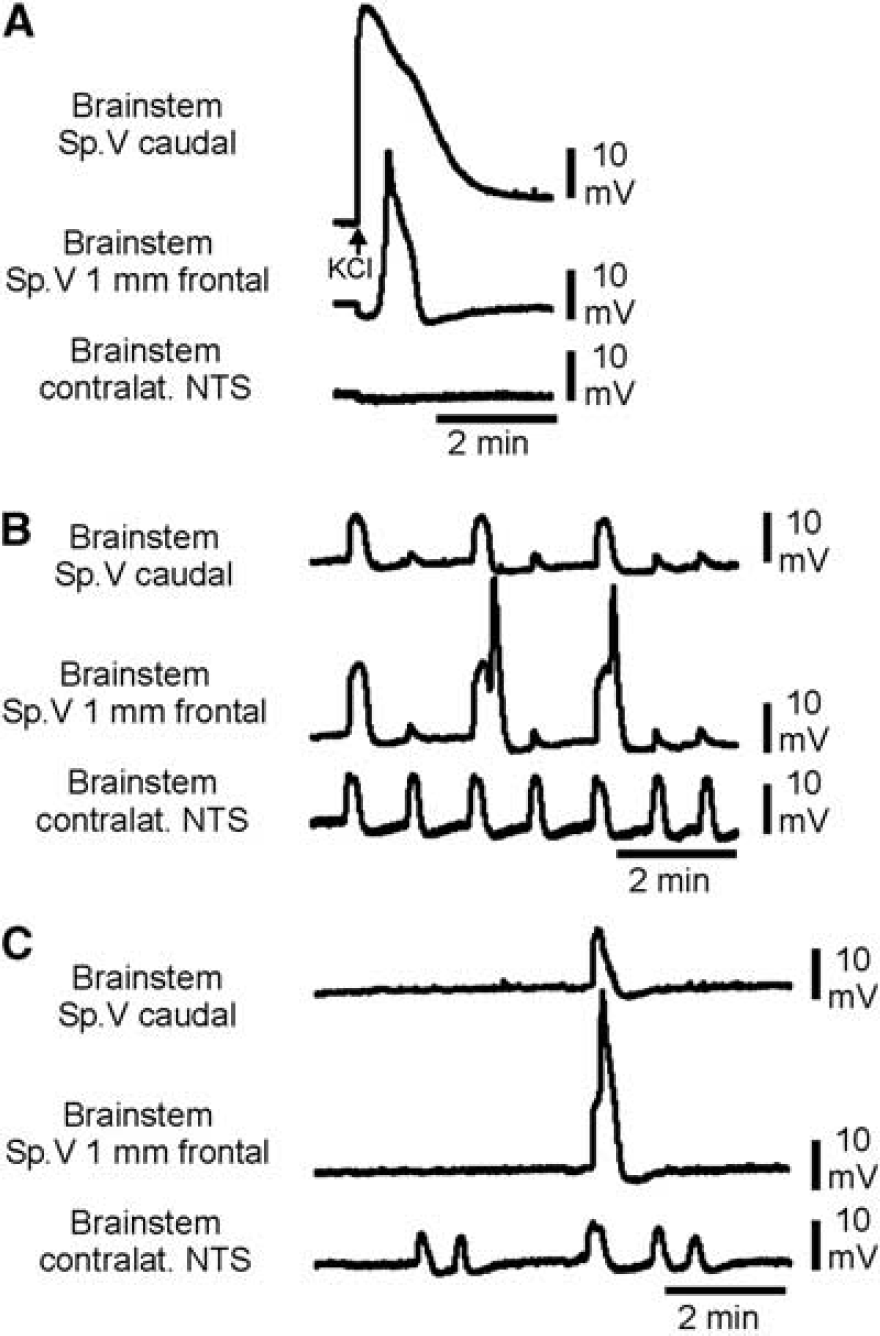
Direct current (DC) recordings showing single spreading depolarization (SD) evoked by KCl and repetitive depolarizations (RDs) in the brainstem. (
In adult rat brainstem, slice preparations (Funke et al, 2009) were able to induce hypoxic SD in the nucleus tractus solitarii (NTS) area and supposed that the adult NTS area should be more susceptible to SD than other parts of the brainstem. Therefore, we recorded DC shifts both in the Sp. V area and in the contralateral NTS in six rats. In five out the six rats, we observed altogether 178 RDs. The almost simultaneous appearance of DC shifts in the whole brainstem (Figure 3B) differed from SD evoked by KCl, which migrated only ipsilaterally and did not invade into the contralateral side of the brainstem (Figure 3A). A typical feature of the depolarization waves in the NTS area was their very uniform shape. Depolarizations in the NTS started 2.2±2.1 seconds earlier than those in the contralateral Sp. V area of the brainstem (Figure 3B). They occurred more regularly than in the Sp. V area, i.e., depolarizations in the NTS were not always accompanied by depolarizations in the Sp. V area (Figure 3C). In Table 1, the main features of the RDs in rat brainstem are summarized.
Parameters of RDs in adult rat brainstem in trigeminal nucleus (Sp. V) area and nucleus tractus solitarii (NTS) area
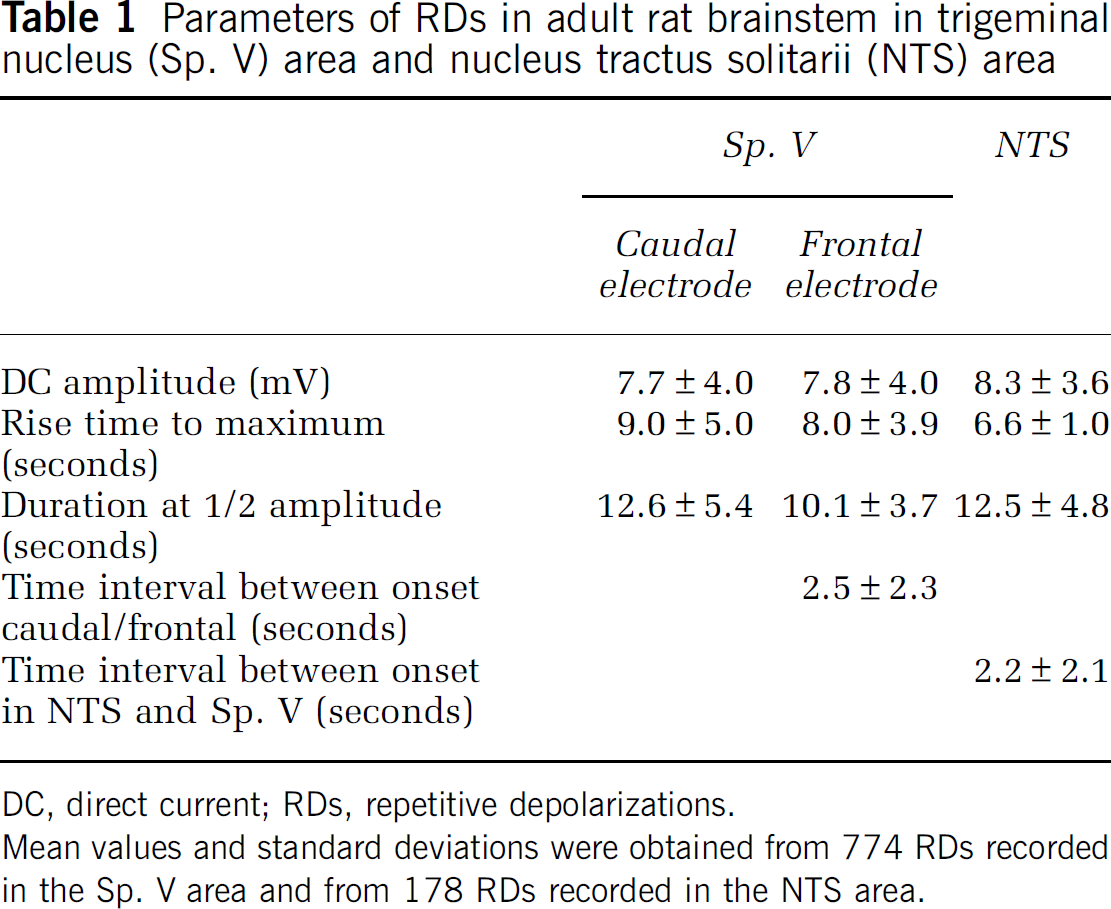
DC, direct current; RDs, repetitive depolarizations.
Mean values and standard deviations were obtained from 774 RDs recorded in the Sp. V area and from 178 RDs recorded in the NTS area.
Influence on Cardiovascular Parameters
To investigate whether RDs influence HR, systemic ABP, and local blood flow at the surface of the brainstem or at the cerebral cortex, we measured these parameters together with DC recordings in a group of 10 rats. In all, 5 out of 10 rats developed RDs, and typical cardiovascular effects associated with DC shifts are depicted in Figure 1.
The RDs were accompanied by significant increases of the mean ABP by 80.6±19.7%. During SD evoked by KCl mean ABP increased significantly by 53.5±26.1%, but this increase was significantly smaller than the former one. No change in mean ABP occurred during epochs without depolarizations (Figure 4A). In parallel, the HR changed but the pattern of change differed between the animals: in one animal HR increased by only 3.9 b.p.m.; in two rats HR showed a slight and short acceleration followed by a deceleration of HR by 19.4 or 21.2 b.p.m. (Figure 1, left column), and in the other two rats HR accelerated by 15.9 or 16.3 b.p.m. (Figure 1, right column), respectively. Some HR changes were accompanied by arrhythmia for some seconds, but both the arrhythmia and the frequency change returned to normal after the depolarization shifts.
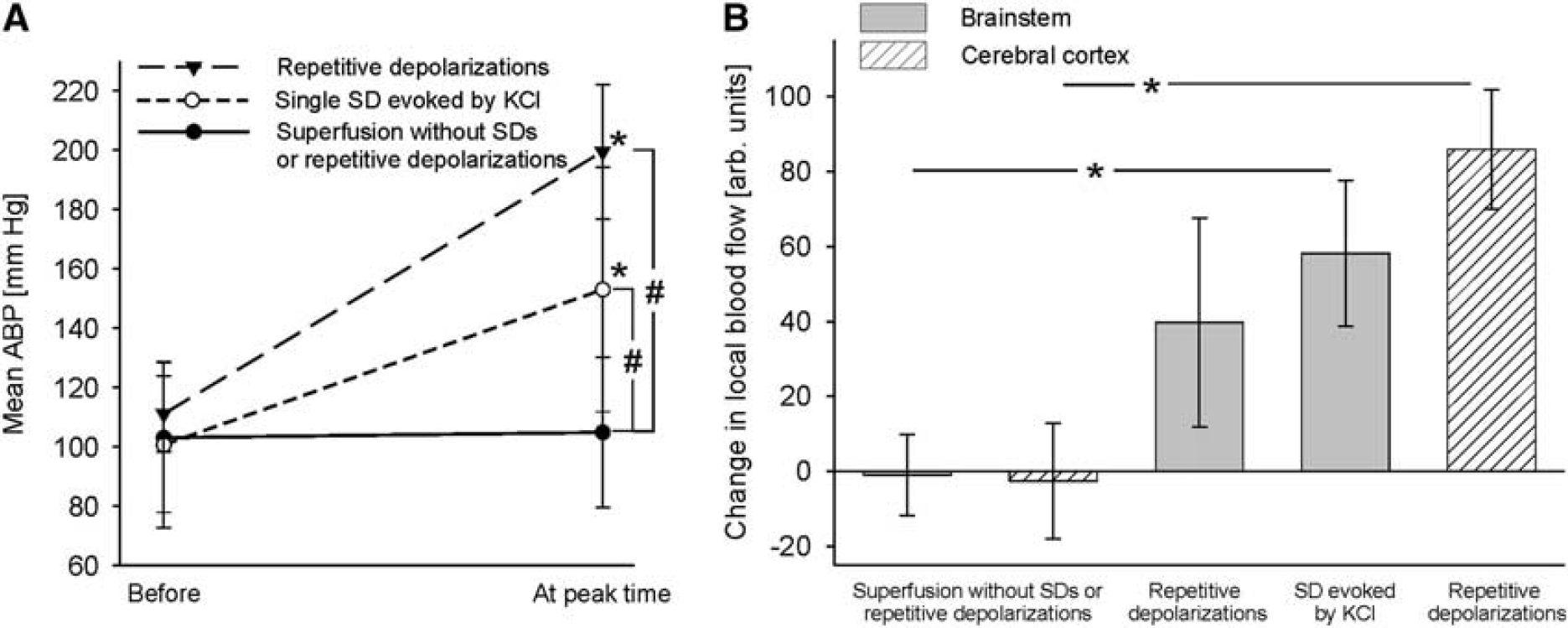
Changes of mean arterial blood pressure (ABP) and local blood flow during single spreading depolarizations (SDs) evoked by KCl and repetitive depolarizations (RDs). (
Each transient increase in ABP was accompanied by a significant but transient increase in blood flow in brainstem and cerebral cortex that was in a close correlation with the onset of the ABP increase but outlasted the normalization of ABP. The increase in local blood flow at the cerebral cortex was larger than at the brainstem. In the brainstem, however, the increase in blood flow did not differ significantly between RDs or SD elicited by microinjection of KCl. If no depolarization occurred, local blood flow did not change either (Figure 4B).
Depolarization and Neuronal Activity in the Brainstem
To further explore the nature of RDs, we recorded extracellularly action potentials from neurons in the caudal trigeminal nucleus that received sensory input from the dura mater. In a close distance to the carbon fiber electrode for recordings of action potentials, the electrode array for KCl microinjection and DC recording was inserted. We specifically asked whether KCl-evoked SDs evoked by KCl and RDs evoke similar changes of neuronal activity.
In six rats, we recorded altogether 40 SD evoked by KCl after conditioning of the brainstem with acetate-ACSF+TEA+KCl. These SD had amplitudes of 24.5±7.2 mV, a mean duration of 19.4±5.1 seconds at 50% of maximal amplitude, and propagated with 4.0±1.5 mm/min across the brainstem. It was noted that during conditioning ongoing neuronal activity at the recording site increased (Figure 5A). During the depolarizing phase of an SD, a burst of action potentials was observed in the brainstem that ended when the negative DC shift reached its maximum. The bursts were followed by a short period of complete neuronal silencing that lasted as long as the DC potential repolarized and reached its lowest amplitude. Neuronal activity reappeared after the SD (Figure 5B). When the KCl microinjection failed to ignite an SD, no burst of action potentials was observed.
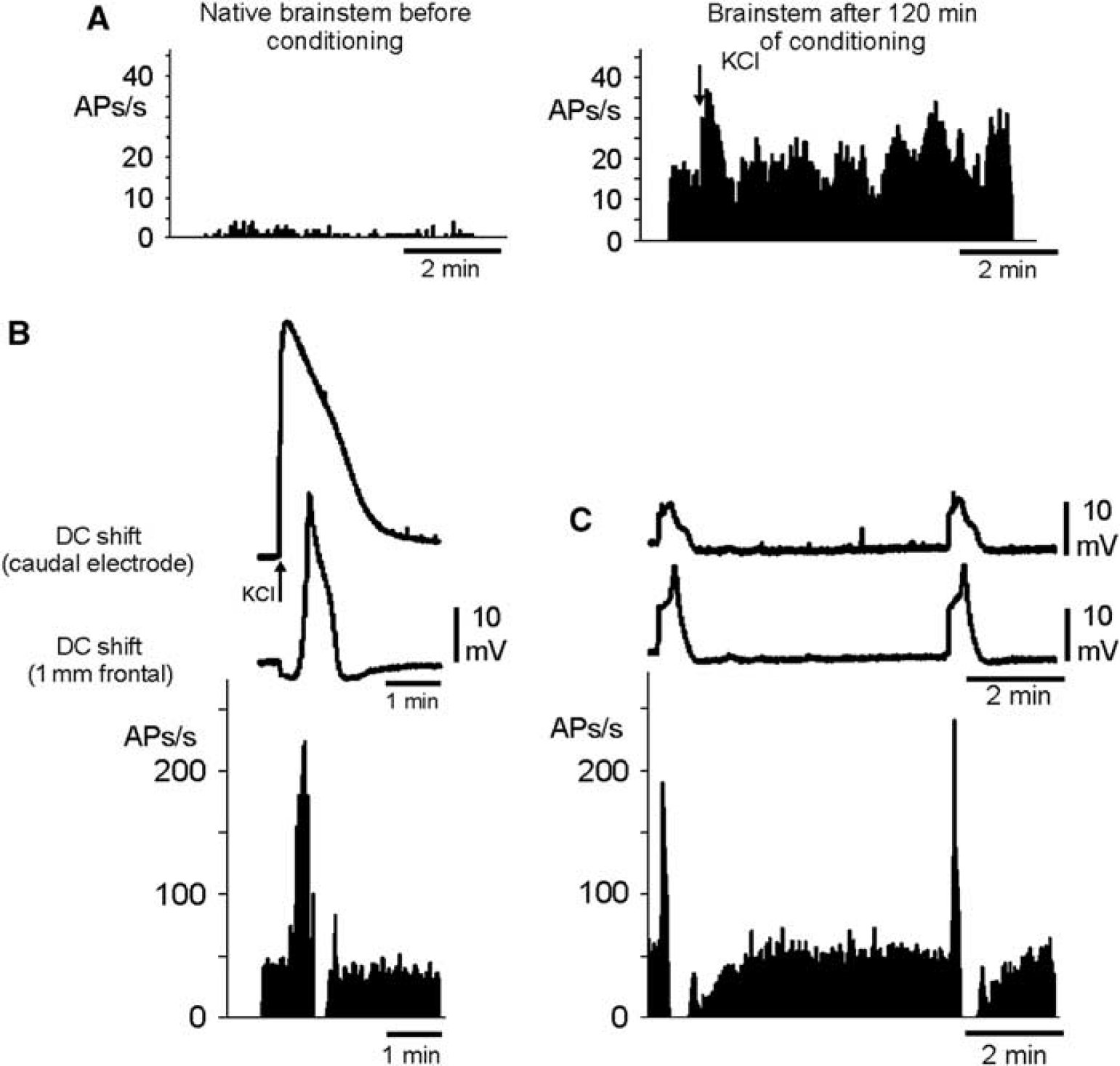
Elicitation of bursts of action potentials of brainstem neurons by repetitive depolarizations (RDs). (
A similar pattern of neuronal activity was observed during RDs in the conditioned brainstem (five out of six rats tested). During each depolarizing phase, a strong burst of action potentials was observed that lasted to the maximum of the DC shift, and after neuronal silencing activity reappeared with return of the DC potential to baseline (Figure 5C). The bursts of action potentials occurred as long as RDs were observed, and successful blockade of the depolarizations stopped the bursts of action potentials as well.
Repetitive Depolarizations Do Neither Elicit Cortical SD Nor Activate Nerve Endings in the Dura Mater
An intriguing question in the context of migraine is whether the brainstem triggers cortical changes and whether brainstem activation may cause an activation of the sensory fibers and thereby produce a neurogenic inflammation in the dura (Williamson and Hargreaves, 2001). In most rats, at least one DC microelectrode was inserted in the ipsilateral parieto-occipital cerebral cortex at a depth of 1,200
Furthermore, we did not observe changes of CGRP release at the intact outer surface of the dura mater in the six animals used for action potential recording. The basal release of CGRP was 4.9±0.8 pg/ml (
Repetitive Depolarizations Depend on Glutamatergic Signaling
First, we tested whether TTX influenced the occurrence of RD. Within 1 to 2 minutes after the application of TTX to the brainstem, depolarizing waves were abolished and did not recover within 30 to 45 minutes of washout with acetate-ACSF+TEA+KCl (Figure 6). The effect of TTX indicates a neuronal origin of RDs. It was noted that TTX also prevented the generation of SD usually evoked by microinjection of KCl, which is different from the TTX effect in the cortex (Sugaya et al, 1975).
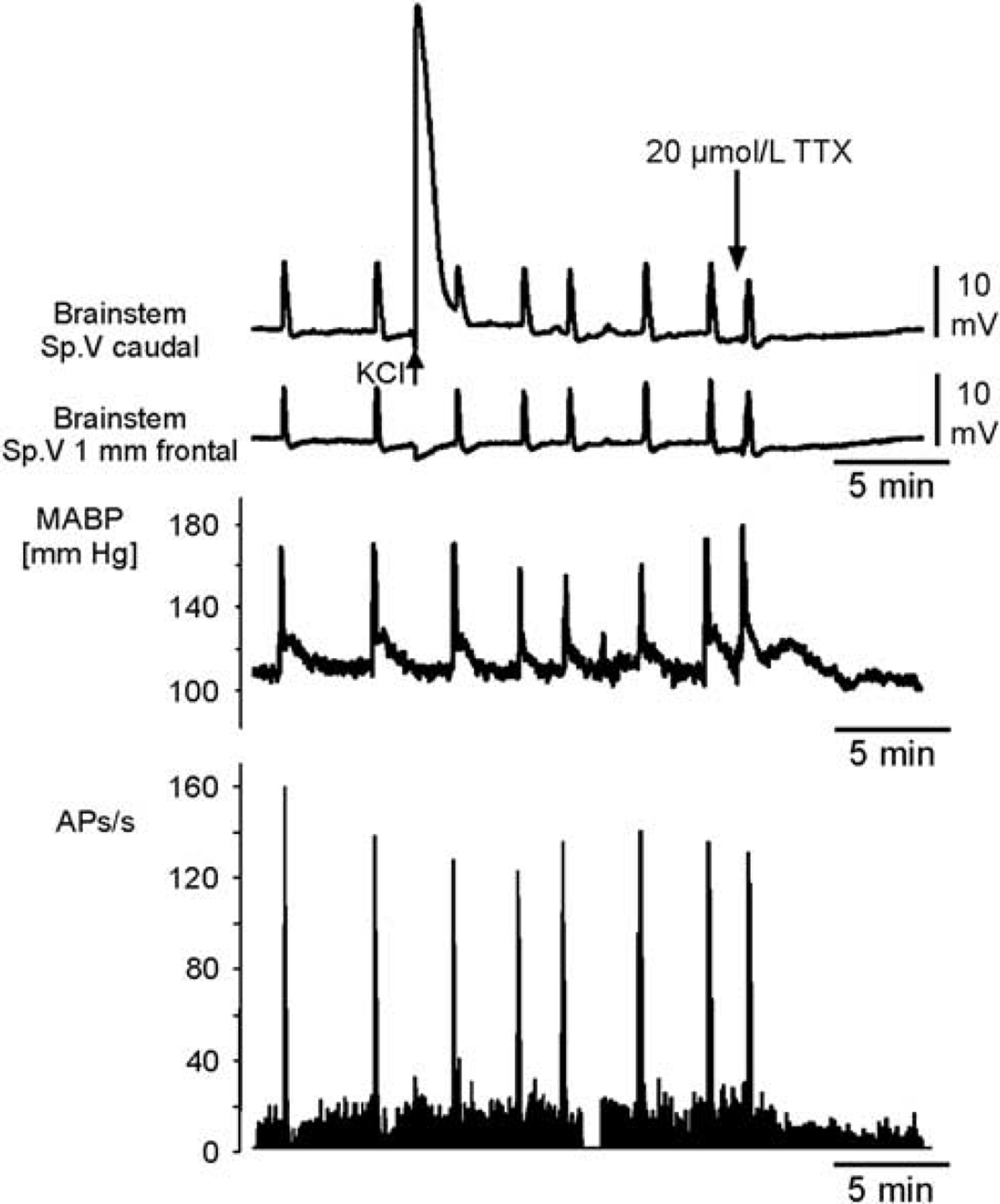
Simultaneous recording of direct current (DC) potentials, mean arterial blood pressure (ABP), and neuronal activity in the brainstem during repetitive depolarizations (RDs). Comparison of DC shifts in the brainstem at two recording sites in the trigeminal nucleus area (Sp. V), of simultaneously recorded changes in mean ABP (MABP), and of neuronal activity (APs) in the Sp. V area close to the site of DC electrodes during RDs after conditioning with acetate-ACSF+TEA+KCl. Note the bursts of action potentials during the depolarizing phases of the DC shifts. A microinjection of KCl (arrow in the top trace, large DC shift) did not induce a propagating spreading depolarization (SD). Topical application of 20
To explore whether glutamatergic mechanisms are involved in RDs, we administered the
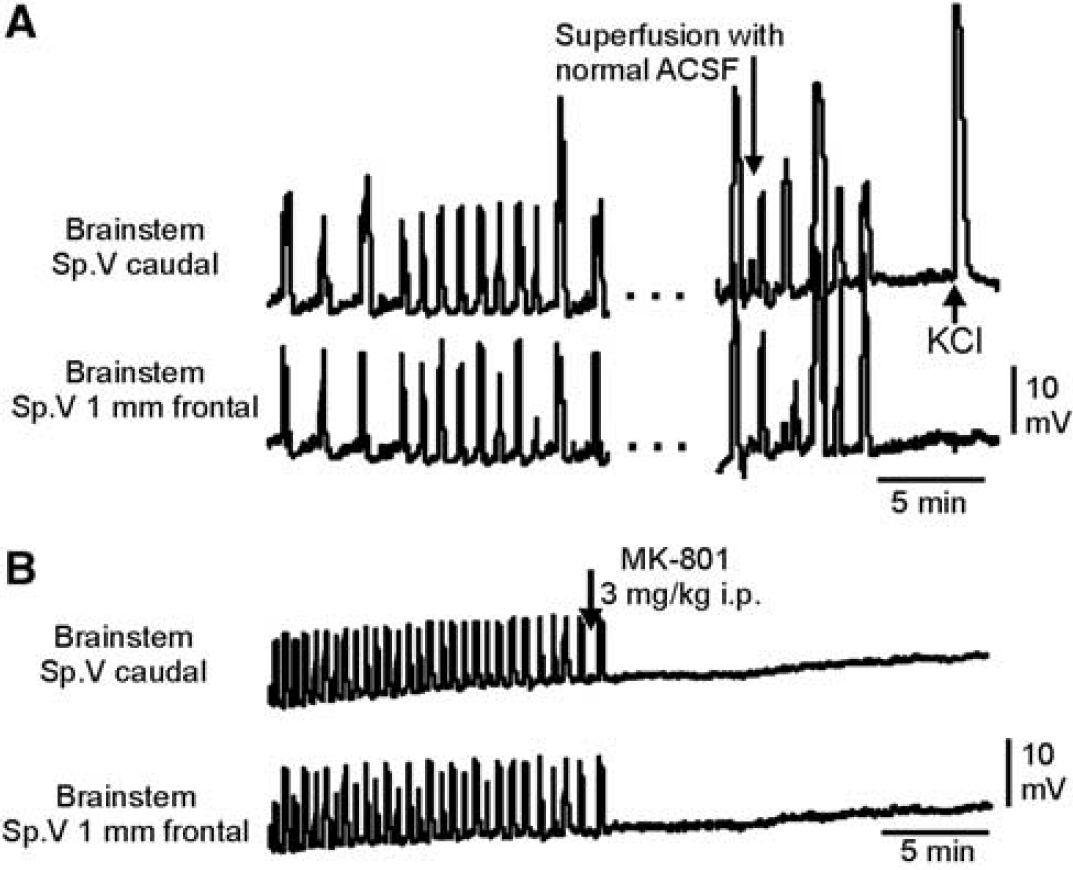
Termination of repetitive depolarizations (RDs) by either normal ACSF or NMDA receptor antagonist. (
Discussion
In the present study, we detected for the first time that the brainstem of the adult rat has the potential to generate RDs and we explored the nature of such depolarizations. We found that RDs occur after an ample period of enhanced neuronal excitability in the brainstem. The RDs are of neuronal origin. They induce bursts of action potentials, then they propagate into centers of cardiovascular control similarly as SDs evoked by KCl. The RDs depend on the functioning of TTX-sensitive sodium channels and on glutamatergic neurotransmission, and they influence a much larger area of the brainstem than KCl-evoked SDs.
The RDs did not result from deterioration of brain tissue because they were closely linked to the conditioning with acetate-ACSF+TEA+KCl and abolished by a simple washout of the brainstem tissue with normal ACSF, which restored previous excitability. We did not find factors for the nonoccurrence of RDs in one third of the animals. Doses of anesthesia, surgical procedures, or durations of the preparations did not differ between rats with or without RDs.
The RDs fulfilled some of the major criteria of ‘spreading depolarizations’ as stated by Somjen (2001), since (1) they depolarized the brainstem tissue and (2) they had amplitudes larger 5 mV and a steep depolarizing phase. However, their DC amplitudes were only 7 to 12 mV whereas KCl-evoked SDs have amplitudes of 15 to 33 mV, and they did not increase with the duration of conditioning. Furthermore, RDs spread 20 to 30 times faster than in KCl-induced SDs and reached the whole brainstem including the contralateral NTS area. Time intervals between the depolarizations shortened with ongoing repetitions. It is possible that electric junctions among neurons (Herreras et al, 1994) or fast propagating Ca++ waves as described for the hippocampus (Kunkler and Kraig, 1998) contribute to this high propagation velocity.
Similarly as SDs in the cerebral cortex (Marrannes et al, 1988; Bureš et al, 1974) or in the brainstem, RDs involved glutamatergic neurotransmission, in particular NMDA receptors. Systemic application of the NMDA receptor antagonist MK-801 quickly stopped RDs. Activation of NMDA receptors during SD is thought to be evoked by an initial increase of [K+]e, which produces an initial depolarization and a consequent release of glutamate, and further increases of [K+]e then cause the spreading of depolarization (Somjen, 2001). Because the conditioning solution contained K+, the elevation of K+ might be also an important factor in the generation of RDs. This is in line with observations in rat hippocampus, where microdialysis with high K+ solution ignited series of recurrent SDs (Herreras and Somjen, 1993), and in rat cerebral cortex, where series of SD were observed during spreading ischemia (Windmuller et al, 2005). The relation to an elevated [K+]e is furthermore underscored by the lack of occurrence of RDs in the immature rat brainstem, where the conditioning superfusate did not contain higher K+ concentrations (Richter et al, 2003). However, two observations are noteworthy that suggest differences between mechanisms of KCl-evoked SD and RDs. The KCl-evoked SD depends on synaptic transmission (i.e., functioning of glutamatergic receptors) and glial functioning but not on the functioning of voltage-gated sodium channels. Sugaya et al (1975) showed that 10
An important aspect of the present study is the change of cardiovascular functions during RDs. The systemic blood pressure (ABP) showed always an increase which was time locked to each depolarization, suggesting that DC shifts affected also blood pressure control centers. By contrast, the patterns of HR changes differed. The reduction of heart frequency may be a reflex response to the increase of blood pressure. The increase of HR may be a response of sympathetic activation. Increases in local blood flow both at the surface of the brainstem and cerebral cortex were also tightly linked to the RDs and the changes in ABP. We could not explore possible influences of RDs on respiration because the animals had to be ventilated to avoid muscle contractions.
To further elucidate possible clinical consequences of RDs, we addressed the question of whether they may trigger SD in the cerebral cortex and an activation of trigeminal afferent fibers in the dura. This question was based on the consideration that neuronal events in the brainstem are involved in the triggering of migraine attacks (Akerman et al, 2011). In none of the rats, any SD-related DC deflections in the cortical gray matter occurred time locked to brainstem depolarizations or KCl-evoked SDs in the brainstem. Furthermore, based on the release of CGRP from the dura, we did not observe an activation of trigeminal afferents during RDs. Thus, it is unlikely that RDs can trigger peripheral or cortical events outside the brainstem. However, the massive activation of neurons in the spinal trigeminal nucleus may indicate the potency of brainstem DC potentials to generate activity in the spinal trigeminal system (see Weiller et al, 1995). In basilar-type migraine, patients exhibit typical brainstem symptoms in the aura phase (Kirchmann et al, 2006). Furthermore, ‘hyperintensities’ in the brainstem of migraineurs were seen in magnetic resonance imaging investigations more often than in healthy control subjects (Kruit et al, 2006). The neurologic basis for such activations is unclear yet, but depolarizations as shown in this study might be a possible candidate.
In summary, we report that the adult brainstem has the potential to produce RDs after a proper conditioning. Though the native brainstem is relatively resistant against SD or other forms of depolarization, it can develop episodes of depolarizations that are primarily not life-threatening in a ventilated animal and can last >1 hour. The RDs share many features with SD evoked by KCl, which clearly indicates that they belong to the SD continuum: they are characterized by steeply rising negative DC shifts, they induce bursts of action potentials, and they depend on glutamatergic neurotransmission. At the mechanistic level, these depolarizations seem to involve the same receptors and ion channels as KCl-evoked SDs. In contrast to the latter, RDs depend of TTX-sensitive sodium channels, they affect a much larger area (or probably the whole brainstem) and they propagate much faster. Thus, RDs represent a spatially more extensive form of hyperexcitability than locally arising and spreading depolarizations. Whether this form of depolarization might be of importance in brainstem pathophysiology will be investigated in future experiments.
Footnotes
Acknowledgements
The authors thank Ms Konstanze Ernst and Ms Wibke Lütz for excellent technical assistance.
Disclosure/conflict of interest
The authors declare no conflict of interest.