Hypoxic preconditioning is thought to rely on gene products regulated by hypoxia-inducible factor (HIF)-1. Here, we show that the HIF-1 target gene cyclin-dependent kinase inhibitor 1, p21WAF1/CIP1, is essential for neuroprotection by hypoxic/aglycemic or erythropoietin preconditioning using wild-type and p21WAF1/CIP1-deficient neurons. Furthermore, overexpression of wild-type p21WAF1/CIP1 or phospho-mutants significantly increased cell death after hypoxia/aglycemia. Moreover, deferoxamine-induced endogenous tolerance did not involve p21WAF1/CIP1 expression in cortical neurons. Our data suggest that balanced expression and potentially posttranslational regulation of p21WAF1/CIP1 is required for hypoxic preconditioning.
Neuroprotective effects of hypoxic preconditioning are well described and complex mechanisms have been proposed to mediate protection observed in various experimental models.1 We have previously shown that neuroprotection can be afforded by different preconditioning paradigms such as erythropoietin (EPO), deferoxamine (DFO), and sublethal periods of combined hypoxia/aglycemia (H/A).2,3 Here, hypoxia-inducible factor (HIF)-1-dependent gene expression is an important mediator of neuroprotection.4
The cyclin-dependent kinase inhibitor 1, p21WAF1/CIP1, is a HIF-1 target gene and increased expression has been observed after global brain ischemia in the hippocampus5 as well as by pharmacological stabilization of HIF-1 by DFO in undifferentiated embryonic neurons.6 Neuroprotection by p21WAF1/CIP1 involves the inhibition of apoptosis signal-regulating kinase, an upstream SAPK/JNK pathway regulator,7 as well as inactivation of caspase 3.8 Furthermore, cytoplasmic localization and under certain circumstances neuroprotection of p21WAF1/CIP1 are regulated by phosphorylation on T145 and S146 by Akt-kinase,9,10 one of the major protein kinases important for neuronal preconditioning.2
Although potential functions of p21WAF1/CIP1 have been delineated for neuroprotection under different conditions, its role in preconditioning has not been studied. Thus, we aimed at investigating the contribution of p21WAF1/CIP1 to neuroprotection in established preconditioning models and to study the effects of p21WAF1/CIP1 phosphorylation on neuronal viability after in vitro ischemia.
MATERIALS AND METHODS
Cortical neuronal cell cultures
All experiments were carried out with notice to the regional authorities (Landesamt für Arbeitsschutz, Gesundheitsschutz und technische Sicher-heit Berlin). Primary cultures of cortical neurons were prepared from mouse (E16–E18; B6;129-Cdkn1atm1Tyi or B6/129SF2/J; The Jackson Laboratory, Bar Harbor, ME, USA) or rat (E18; Forschungseinreichtung für Experimentelle Medizin, Charité) embryos as described.2 In brief, cortices were dissected, the meninges were removed and the tissue was incubated in trypsin/EDTA (0.05/0.02% w/v in PBS (phosphate-buffered saline)) at 37 °C for 15 minutes. After rinsing with PBS and dissociation medium (modified Eagle's medium supplemented with 10% fetal calf serum, 10mM HEPES, 44mM glucose, 100 U/mol/L penicillin/streptomycin, 2mM L-glutamine, 100 IE insulin/l), tissue was dissociated and plated in starter medium (Neurobasal medium™ supplemented with B27™ (further referred as NBM, Invitrogen, Paisley, UK, 100 U/mL penicillin/streptomycin, 0.5 mM L-glutamine, 25 μM glutamate). The cultures were used for experiments after 8 days in vitro.
Preconditioning Protocols
Hypoxic preconditioning was performed in a gas tight anaerobic chamber (Concept 400, Ruskinn Technologies, Pencoed, UK) for 90 minutes 48 hours before hypoxia/aglycemia (H/A) as described previously.2 Treatment with EPO (final concentration 100 U/L, Sigma-Aldrich, Deisenhofen, Germany) was initiated 48 hours before H/A. Concentration has been chosen based on previous studies showing a neuroprotective effect after H/A2. Alternatively, 150 μM DFO (Sigma-Aldrich) in PBS or PBS alone was applied to neuronal cultures on day 8 in culture as described.11
Hypoxia/Aglycemia
Combined hypoxia/aglycemia was performed as described previously.2 Duration of H/A has been evaluated in preliminary experiments in which p21WAF1/CIP1+/+ and p21WAF1/CIP1–/– neurons had been exposed to 120, 150, and 180 minutes of H/A. For the present study, 120 minutes were chosen. Residual oxygen was removed by a palladium catalyzer. Hypoxia/aglycemia (120 minutes) in the overexpression experiments was performed in an inVivo2–400 (Ruskinn) chamber essentially as described previously.11 Oxygen concentration in these experiments was 0.3%.
Plasmids, cloning, and transfection
Cloning and transfection (see Supplementary Material) were performed as described.11
Quantitative Real-Time PCR
Preparation of total mRNA and qPCR measurements (see Supplementary Material) were performed as described in Mergenthaler et al11 on selected samples (N = 6) from the preconditioning series described therein.
Cell Death Assessment
Cell death was assessed by measurement of lactate dehydrogenase (LDH) activity in the supernatant medium. In neuronal cultures, LDH release into the medium reliably correlates with the number of damaged cells.2 Cell death in the overexpression experiments was analyzed by the cotransfection/cocultivation approach described previously11 (see Supplementary Material).
Statistics
Preconditioning experiments were performed in triplicate (three independent cell culture preparations) with 16 to 48 cell cultures per experimental condition. Data are presented as mean values ± s.d. Box plots were generated in SPSS Statistics 20.0 (IBM SPSS GmbH, Ehningen, Germany) and display median, 25th and 75th percentiles within boxes and 5th and 95th percentiles with whiskers. Statistical analyses and number of independent experiments (N) are indicated in the figure legends (Figures 1 and 2).
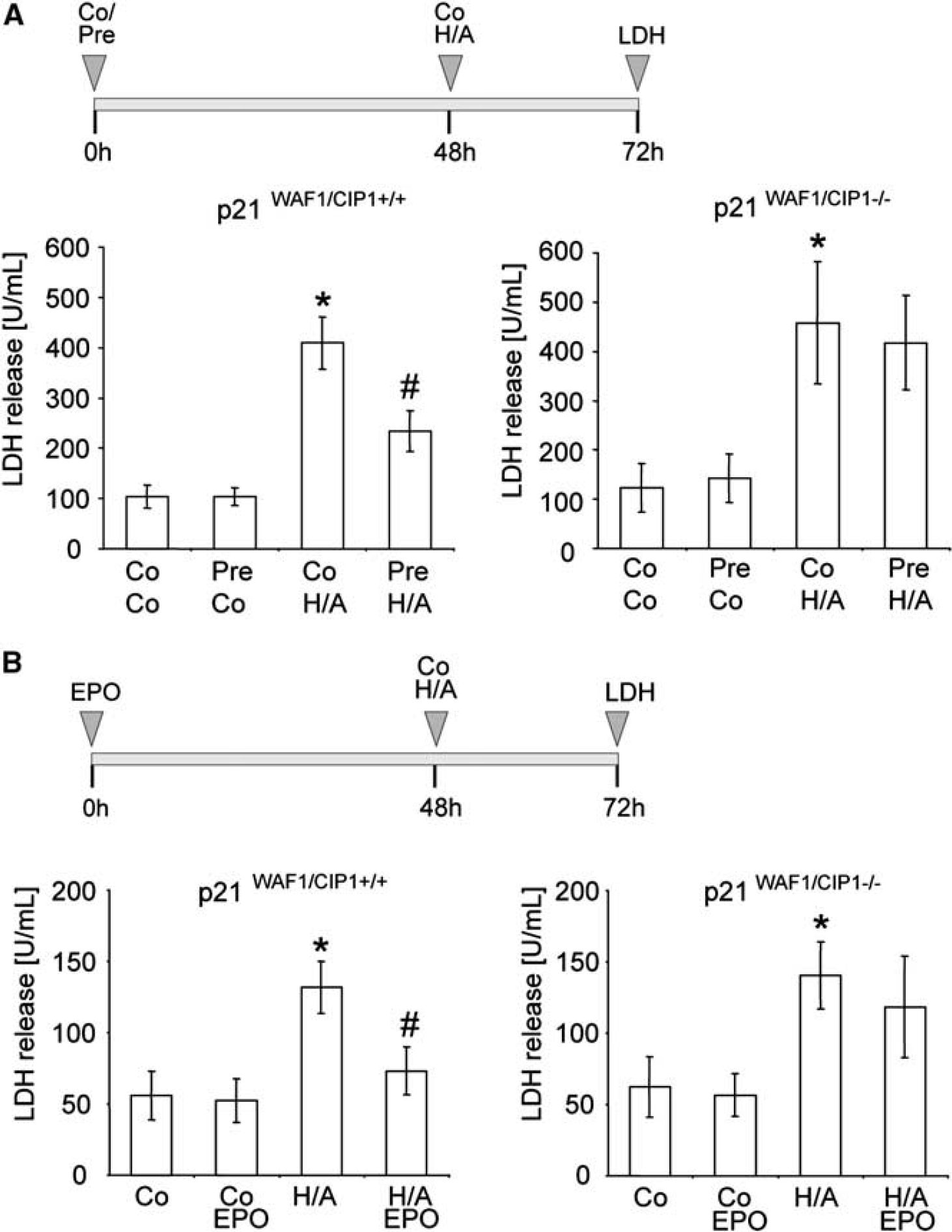
P21WAF1/CIP1 is required for hypoxic preconditioning and erythropoietin (EPO)-mediated neuroprotection. (A) Cultures of cortical neurons from p21WAF1/CIP1+/+ and p21WAF1/CIP1–/– mice were exposed to a sublethal preconditioning hypoxia/aglycemia for 90 minutes. Twenty-four hours after a lethal H/A (120 minutes), neuronal survival was determined by LDH activities in the supernatant medium. (B) Effect of preconditioning by recombinant human EPO (EPO, 100 U/L) on p21WAF1/CIP1+/+ and p21WAF1/CIP1–/– neurons 24 hours after H/A measured by LDH activities in the supernatant medium. Data are presented as mean values ± s.d., *P < 0.05, one-way analysis of variance (ANOVA) followed by Bonferroni correction. Co, normoxic control; Pre, sublethal H/A (preconditioning); H/A, hypoxia/aglycemia.
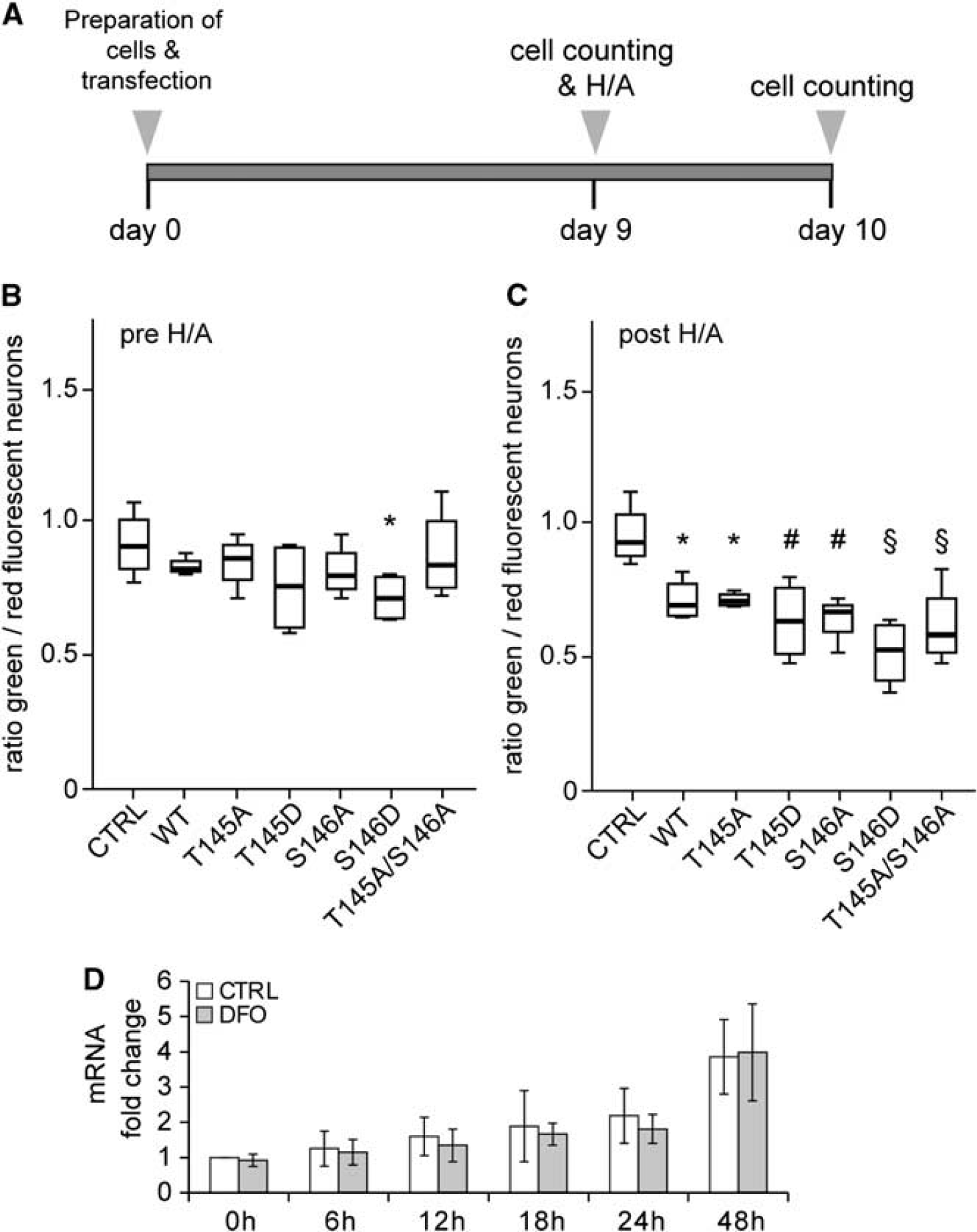
Expression of p21WAF1/CIP1 after preconditioning and influence of p21WAF1/CIP1 overexpression on neuronal survival after H/A. (A) Experimental outline of p21WAF1/CIP1 overexpression experiments. Results obtained from counting cells before H/A are shown in (B), 24 hours after in (C). (B, C) Analysis of neuronal survival expressed as ratio of green/red fluorescent neurons before (pre H/A) and 24 hours after (post H/A) H/A. P values (compared with control, CTRL, transfection and cocultivation of H2b–GFP and H2b-mOrange only): (A) * = 0.031 and (B) * = 0.006; # = 0.001; § < 0.001; analysis of variance (ANOVA) with unprotected Fisher's LSD posttest. N = 4. WT: p21WTWAF1/CIP1; T145A: p21T145AWAF1/CIP1; T145D: p21T145DWAF1/CIP1; S146A: p21S146AWAF1/CIP1; S146D: p21S146DWAF1/CIP1; T145A/S146A: p21T145A/S146A.WAF1/CIP1. (D) Analysis of p21WAF1/CIP1 mRNA expression after hypoxia-mimicking preconditioning with deferoxamine (DFO) (150 μM), indicating that p21WAF1/CIP1 mRNA expression is not increased after activation of hypoxia-inducible factor (HIF)-1 in primary neurons within the first 48 hours compared with 0 hours control (CTRL, treated with vehicle only). N = 6. H/A, hypoxia/aglycemia.
RESULTS
p21Waf1/Cip1 Deficiency Abolishes Hypoxic Preconditioning- or Erythropoietin-Preconditioning-Induced Neuroprotection To investigate whether p21WAF1/CIP1 has a functional role in hypoxic preconditioning-induced neuroprotection against otherwise lethal H/A, neurons from p21WAF1/CIP1-deficient mice and wild-type neurons (p21WAF1/CIP1+/+) were exposed to a sublethal H/A for 90 minutes and incubated for additional 48 hours before an otherwise lethal H/A (120 minutes; Figure 1A). Preliminary studies were performed to test different preconditioning times (15, 30, 60, and 90 minutes) to afford a maximum time of preconditioning without significant cell death determined by LDH release into the supernatant medium (data not shown). As shown in Figure 1A, p21WAF1/CIP1+/+ neurons were protected by hypoxic preconditioning (LDH activity in the supernatant medium: 234.3 ± 40.1 U/mL) in contrast to p21WAF1/CIP1+/+ neurons exposed to a 90-minute incubation under normoxic conditions 48 hours before H/A (LDH: 409.4 ± 51.8 U/mL). Likewise, significant cell death was found in nonpreconditioned p21WAF1/CIP1–/– neurons (LDH: 458.2 ± 123.6 U/mL), and hypoxic preconditioning did not protect these neurons after H/A (LDH: 417.8 ± 95.8 U/mL). Similar results were obtained in the EPO-preconditioning model. While p21WAF1/CIP1+/+ neurons treated with EPO for 48 hours were protected against H/A (LDH: 73.1 ± 16.8 U/mL; H/A alone LDH: 131.9 ± 18.1 U/mL; Figure 1B), EPO preconditioning of p21WAF1/CIP1–/– failed to mediate neuroprotection after H/A for 120 minutes (LDH: 118.4 ± 35.5 U/mL; H/A alone LDH: 140.6 ± 23.2 U/mL). We conclude that p21WAF1/CIP1 is required for H/A preconditioning- or EPO preconditioning-induced neuroprotection against in vitro ischemia.
Neuroprotection Is Not Related to Increased Expression of p21WAF1/CIP1
We therefore tested whether overexpression of wild-type p21WAF1/CIP1 (p21WAF1/CIP1WT) resulted in increased neuroprotection after H/A (Figure 2A). However, overexpression of p21WTWAF1/CIP1 demonstrated no beneficial effect on survival of rat cortical neurons during the period of cultivation (Figure 2B) and showed no protection after H/A (Figure 2C). Indeed, expression of mutant p21WAF1/CIP1 with locked phosphorylation state (phospho-deficient or phospho-mimetic) resulted in increased cell death in transfected neurons before (Figure 2B) and after (Figure 2C) H/A, albeit with undetectable differences in the global phosphorylation state of nuclear p21T145WAF1/CIP1 (Supplementary Figure 1). While mutants of the primary Akt phosphorylation site (T145) did not alter neuronal survival before H/A, the phospho-mimetic mutant of the weaker Akt-kinase binding site (p21S146D)9 increased neuronal cell death before H/A. However, phospho-deficient, double phospho-deficient, and phospho-mimetic mutants increased neuronal cell death after H/A. Furthermore, pharmacologic preconditioning of cultured primary embryonic rat brain cortical neurons (p21WAF1/CIP1+/+) with the HIF-1 stabilizing iron chelator DFO, a well-established HIF-1-activating and neuroprotective treatment,3,11 did not result in an increased p21WAF1/CIP1 mRNA expression compared with control-treated neurons within 48 hours (Figure 2D). In summary, these results indicate that p21WAF1/CIP1 is necessary for neuronal preconditioning. However, our data further suggest that balanced expression levels and potentially posttranslational modification rather than transcriptional activation of p21WAF1/CIP1 via HIF-1 is important for the regulation of its neuroprotective properties.
DISCUSSION
In the present study, we investigated the role of p21WAF1/CIP1 in in vitro models of neuronal preconditioning. We found that p21WAF1/CIP1 plays a crucial role for hypoxic and EPO-mediated preconditioning after combined hypoxia/aglycemia. Furthermore, we demonstrate that neuroprotection in this model relies on balanced expression levels of p21WAF1/CIP1.
Upregulation of p21WAF1/CIP1 has been demonstrated upon ischemia,5 and the present study supports that p21WAF1/CIP1 is involved in neuronal preconditioning. We conclude that neuroprotection mediated by preconditioning requires a well-balanced expression of p21WAF1/CIP1. Our data further indicate that under certain conditions altered phosphorylation of p21WAF1/CIP1 rather than changes in expression levels may regulate the function of p21WAF1/CIP1 in neuroprotection. Importantly, neuroprotection was abolished after hypoxic and EPO preconditioning in p21WAF1/CIP1–/– neurons, further suggesting a common mechanism involving p21WAF1/CIP1 in both preconditioning models. This includes common downstream targets such as caspase 3 inhibition via protein kinase B phosphorylation2,8 or the inhibition of stress activated protein kinases/Jun kinases.12 Furthermore, both hypoxic and EPO preconditioning activate protein kinases such as protein kinase B2, further supporting that phosphorylation of p21WAF1/CIP1 is important for its stability and functional regulation.9
Thus, we studied the impact of p21WAF1/CIP1 phosphorylation on neuronal survival after combined hypoxia/aglycemia. Previous studies have shown that phosphorylation of p21WAF1/CIP1 on T145 and S146 abrogates binding to the PCNA (proliferating cell nuclear antigen).9 Release of PCNA from phosphorylated p21WAF1/CIP1, which is complete from phospho-mimetic p21WAF1/CIP1T145D and incomplete from phospho-mimetic p21WAF1/CIP1S146D supports the function of PCNA in DNA replication and repair,9 which is also important for neuronal recovery after hypoxia or ischemia.13 In accordance with these data, we found that the phospho-deficient p21WAF1/CIP1 mutants (p21T145A, p21S146A, p21T145A/S146A) resulted in increased cell death after H/A. However, overexpression of phospho-mimetic p21WAF1/CIP1 mutants (p21T145D, p21S146D) also resulted in increased neuronal injury after H/A. Given that the detrimental effect of phospho-mutants of p21WAF1/CIP1 on neuronal survival was more pronounced after H/A and given the HIF-1-dependent upregulation of p21WAF1/CIP1 at earlier time points of neuronal differentiation,6 DNA repair may be more relevant for stable maintenance of neuronal DNA during neuronal differentiation, development and long-term cultivation (9 days in our model) than after ischemia.
Hypoxic preconditioning activates HIF-1,4 and HIF-1 target genes are important mediators of preconditioning.1 Upregulation of p21WAF1/CIP1 has been demonstrated for up to 2 hours after hypoxia in the retina14 and in undifferentiated primary cortical neurons after DFO treatment.6 Together with our results from primary cultures of differentiated neurons, these data suggest a tightly regulated transcriptional response and posttranslational modification of p21WAF1/CIP1 after preconditioning. These data indicate a complex role for p21WAF1/CIP1-dependent neuroprotection. In addition to HIF-1 activation, increased expression of p21WAF1/CIP1 may also depend on other transcription factors such as p53.15 Costimulatory molecules such as glucocorticoid or mineralocorticoid receptors may further play an important role in regulating p21WAF1/CIP1 function.10
In conclusion, our data indicate that a balanced expression of p21WAF1/CIP1 is essential for neuronal preconditioning in vitro and could be of relevance for survival of neurons after cerebral ischemia. Moreover, our results suggest that p21WAF1/CIP1-mediated effects may depend on its phosphorylation status rather than on transcriptional upregulation.
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflict of interest.
Footnotes
ACKNOWLEDGEMENTS
The authors thank D Freyer, R Gusinda, K Stohlmann, and M Thielke for experimental support and S Dimmeler for plasmids.
Notes
Supplementary Information accompanies the paper on the Journal of Cerebral Blood Flow & Metabolism website ()
References
1.
MergenthalerPDirnaglU. Protective conditioning of the brain: expressway or roadblock?J Physiol2011; 589: 4147–4155.
2.
RuscherKFreyerDKarschMIsaevNMegowDSawitzkiB. Erythropoietin is a paracrine mediator of ischemic tolerance in the brain: evidence from an in vitro model. J Neurosci2002; 22: 10291–10301.
3.
PrassKRuscherKKarschMIsaevNMegowDPrillerJ. Desferrioxamine induces delayed tolerance against cerebral ischemia in vivo and in vitro. J Cereb Blood Flow Metab2002; 22: 520–525.
4.
RuscherKIsaevNTrendelenburgGWeihMIuratoLMeiselA. Induction of hypoxia inducible factor 1 by oxygen glucose deprivation is attenuated by hypoxic preconditioning in rat cultured neurons. Neurosci Lett1998; 254: 117–120.
5.
TomasevicGKammeFStubberodPWielochMWielochT. The tumor suppressor p53 and its response gene p21WAF1/Cip1 are not markers of neuronal death following transient global cerebral ischemia. Neuroscience1999; 90: 781–792.
6.
ZamanKRyuHHallDO'DonovanKLinKIMillerMP. Protection from oxidative stress-induced apoptosis in cortical neuronal cultures by iron chelators is associated with enhanced DNA binding of hypoxia-inducible factor-1 and ATF-1/CREB and increased expression of glycolytic enzymes, p21(waf1/cip1), and erythropoietin. J Neurosci1999; 19: 9821–9830.
7.
ZhanJEastonJBHuangSMishraAXiaoLLacyER. Negative regulation of ASK1 by p21Cip1 involves a small domain that includes Serine 98 that is phosphorylated by ASK1 in vivo. Mol Cell Biol2007; 27: 3530–3541.
8.
SuzukiATsutomiYAkahaneKArakiTMiuraM. Resistance to Fas-mediated apoptosis: activation of caspase 3 is regulated by cell cycle regulator p21WAF1 and IAP gene family ILP. Oncogene1998; 17: 931–939.
9.
RossigLJadidiASUrbichCBadorffCZeiherAMDimmelerS. Akt-dependent phosphorylation of p21(Cip1) regulates PCNA binding and proliferation of endothelial cells. Mol Cell Biol2001; 21: 5644–5657.
10.
HarmsCAlbrechtKHarmsUSeidelKHauckLBaldingerT. Phosphatidylinositol 3-Akt-kinase-dependent phosphorylation of p21(Waf1/Cip1) as a novel mechanism of neuroprotection by glucocorticoids. J Neurosci2007; 27: 4562–4571.
11.
MergenthalerPKahlAKamitzAvan LaakVStohlmannKThomsenS. Mitochondrial hexokinase II (HKII) and phosphoprotein enriched in astrocytes (PEA15) form a molecular switch governing cellular fate depending on the metabolic state. Proc Natl Acad Sci USA2012; 109: 1518–1523.
12.
LangleyBD'AnnibaleMASuhKAyoubITolhurstABastanB. Pulse inhibition of histone deacetylases induces complete resistance to oxidative death in cortical neurons without toxicity and reveals a role for cytoplasmic p21(waf1/cip1) in cell cycle-independent neuroprotection. J Neurosci2008; 28: 163–176.
13.
LiuPKGrossmanRGHsuCYRobertsonCS. Ischemic injury and faulty gene transcripts in the brain. Trends Neurosci2001; 24: 581–588.
14.
ThierschMRaffelsbergerWFriggRSamardzijaMWenzelAPochO. Analysis of the retinal gene expression profile after hypoxic preconditioning identifies candidate genes for neuroprotection. BMC Genomics2008; 9: 73.
15.
el-DeiryWSTokinoTVelculescuVELevyDBParsonsRTrentJM. WAF1, a potential mediator of p53 tumor suppression. Cell1993; 75: 817–825.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.