
Abstract
It has been hypothesized that myelin acts like a mitochondrion, generating ATP across the membranes of its sheath. By calculating the proton motive force across the myelin membrane based on known values for the pH and membrane potential of the oligodendrocyte, we find that insufficient energy could be harvested from proton flow across the myelin membrane to synthesize ATP. In fact, if the respiratory chain were present in the myelin membrane, then the ATP synthase would function in reverse, hydrolyzing rather than synthesizing ATP. This calculation places the hypothesis of an energy-producing role for myelin in considerable doubt.
INTRODUCTION
The main method of ATP production in the central nervous system is oxidative phosphorylation, which is carried out by billions of specialized organelles, mitochondria. Mitochondria harvest the energy available in their transmembrane proton gradient to form ATP, which is then exported from the mitochondria for use by energy demanding processes in the cell. It has recently been suggested that myelin—the capacitance-reducing sheath surrounding axons—can carry out the same ATP-producing process. 1 We assess the plausibility of this idea, after outlining both the known mechanism of ATP synthesis in mitochondria and the proposed mechanism of ATP synthesis in myelin.
ATP Generation by Mitochondria
Mitochondria are composed of two compartments: an inner mitochondrial matrix bounded by an inner membrane, and an intermembrane space between the inner and outer membranes (Figure 1A). Within the matrix, the citric acid cycle produces NADH and FADH2, which are passed to the respiratory chain in the inner membrane. Chain complexes catalyze NADH and FADH2 oxidation and O2 reduction, and the energy released from these reactions is used to pump protons from the matrix to the intermembrane space. This makes the intermembrane space acidic (pH 6.9), leaving the matrix alkaline (pH 7.8) and negatively charged (–200 mV).2,3
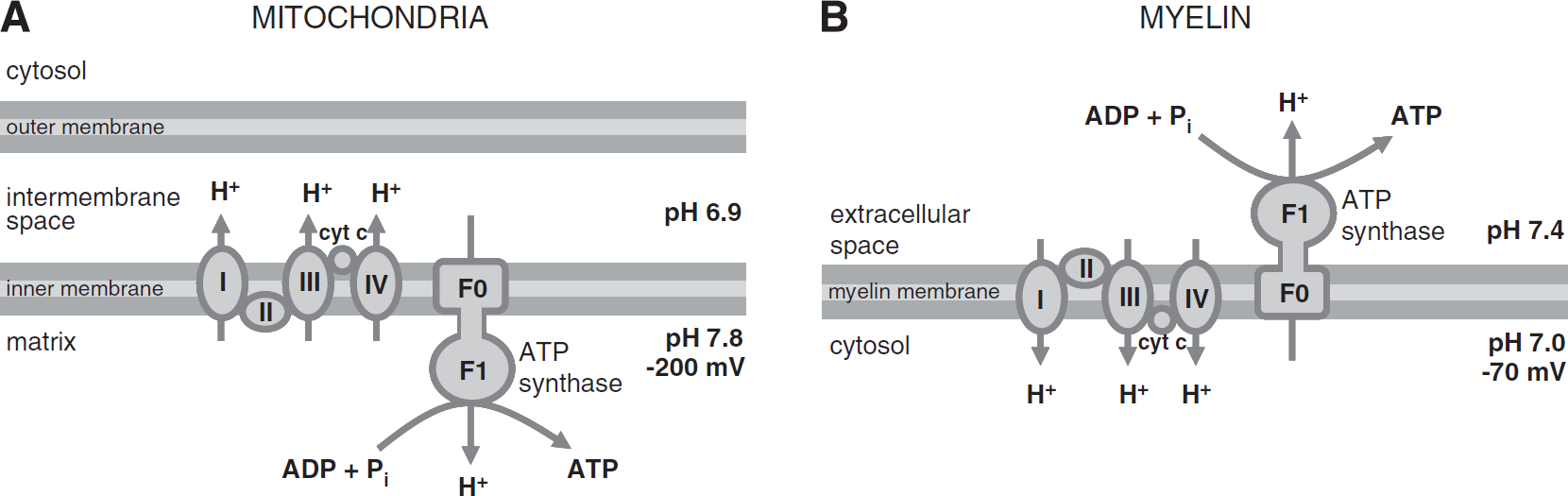
Powering the ATP synthase. (
This electrochemical gradient creates a proton motive force that powers the ATP synthase. Both the electrical and concentration gradients drive protons from the intermembrane space into the matrix. As they flow down this gradient through the FO segment of the ATP synthase, the energy released is used to power ADP phosphorylation by the F1 segment of the ATP synthase. One ATP molecule is generated for every 2.7 protons that enter the matrix this way. 4 The ATP is transported out of the mitochondrial matrix in exchange for a cytoplasmic ADP, and an extra proton is cotransported into the mitochondrial matrix with a Pi molecule (it is critical to keep the ratio of ATP concentration to ADP concentration in the mitochondrial matrix low, and the level of Pi high, so that ATP production rather than hydrolysis is favored).
ATP Generation by Myelin: The Hypothesis
Based on evidence from biochemical assays, western blot analysis, and immunocytochemistry, Panfoli's group have put forward the hypothesis that myelin is able to consume oxygen and produce ATP through the operation of an ATP synthase driven by a proton gradient across the membranes of the sheath.1,5,6 They suggest that mitochondrial fusion with the myelin membrane during formation of the sheath leads to ectopic expression of the respiratory chain complexes and F1FO-ATP synthase in the myelin membranes.
6
The respiratory complexes are proposed to pump protons
One potential problem with this arrangement is the possibility that cytochrome c expressed on the inner surface of the plasma membrane could be released into the cytoplasm where it would trigger caspase-mediated apoptosis of the oligodendrocytes (reviewed in Jiang and Wang 7 ). This is not normally a danger for mitochondrial cytochrome c, because it is confined to the mitochondrial intermembrane space by the outer mitochondrial membrane. Furthermore, whether axons, in fact, require metabolic support from ensheathing oligodendrocytes is debated. Several lines of evidence suggest that myelinating oligodendrocytes provide energetic substrates, such as lactate, to the axons they ensheath, and that this trophic support is essential for axonal integrity.8,9 In contrast, Harris and Attwell 10 have calculated that the glucose supply at nodes of Ranvier and the density of axonal mitochondria could make the need for glial metabolic support of central nervous system axons unnecessary, although it is possible that peripheral nervous system axons, with significantly longer internodes, do require such support.
Although it has been suggested that mitochondria routinely contaminate purified myelin preparations, which would provide an alternative explanation for the presence of mitochondrial proteins in myelin,11,12 we can evaluate the theory put forward by Ravera
METHODS AND RESULTS
For the F1FO-ATP synthase to generate ATP, protons must flow from a state of high potential energy on the side of the membrane expressing the FO segment to a state of low potential energy on the side of the membrane expressing the F1 segment (where ATP is generated). Both electrical and concentration gradients contribute to the potential energy for proton flow across the membrane. Calculating the change in Gibbs-free energy for proton flow (ΔGp) tells us whether protons will spontaneously flow across the membrane in the direction that is required for ATP generation.
In the mitochondrion, the F1 segment of the ATP synthase is in the matrix, so, to synthesize ATP, protons must flow from the intermembrane space to the matrix. Both electrical and chemical gradients across the inner membrane are large and in the right direction, resulting in a negative ΔGp (Table 1A). This indicates that protons will spontaneously flow across the membrane in the direction required for ATP generation.
Calculating proton motive force
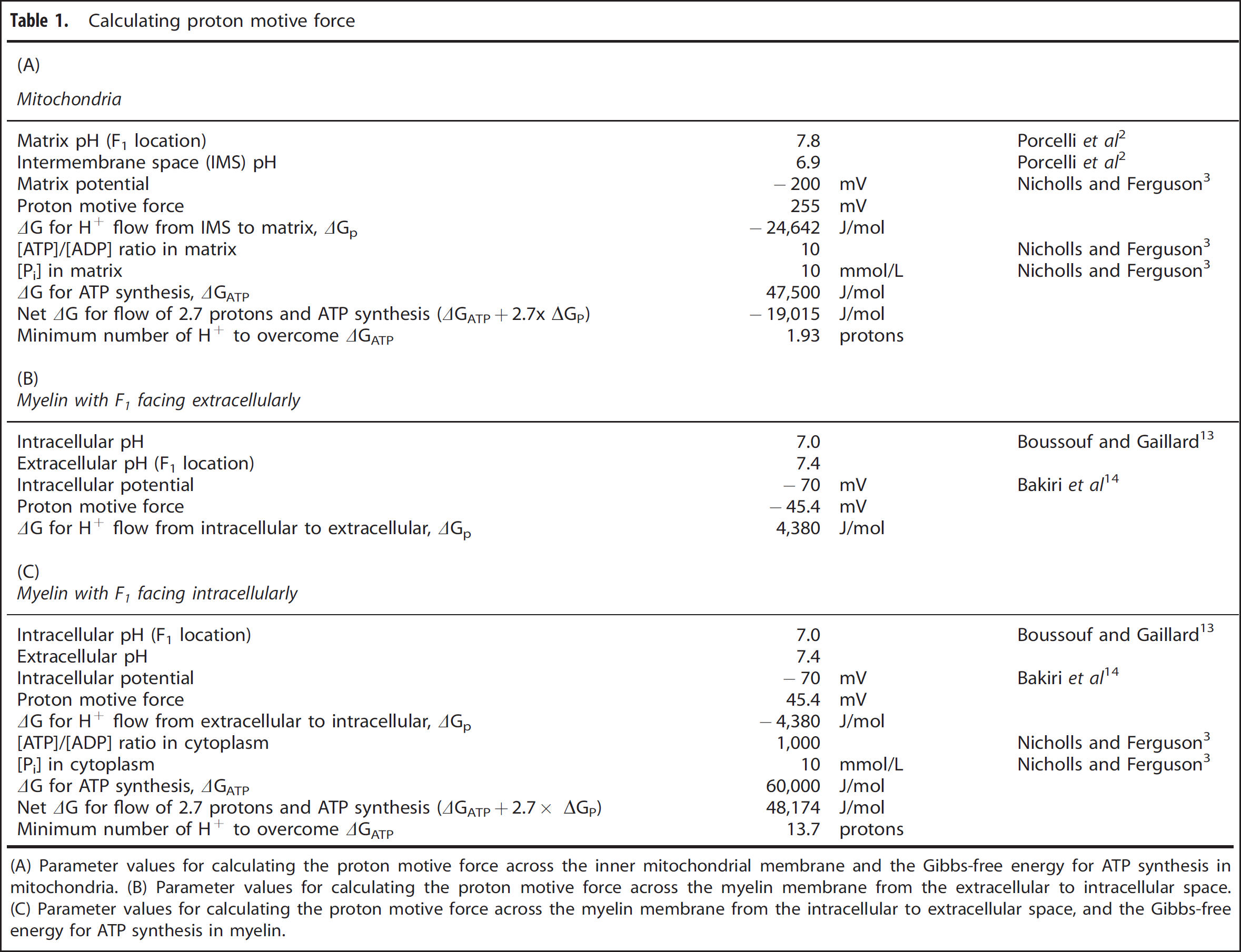
(A) Parameter values for calculating the proton motive force across the inner mitochondrial membrane and the Gibbs-free energy for ATP synthesis in mitochondria. (B) Parameter values for calculating the proton motive force across the myelin membrane from the extracellular to intracellular space. (C) Parameter values for calculating the proton motive force across the myelin membrane from the intracellular to extracellular space, and the Gibbs-free energy for ATP synthesis in myelin.
In the oligodendrocyte, the F1 segment of the ATP synthase is proposed to be in the extracellular space, so protons must flow from the intracellular to the extracellular space. With an intracellular pH of 7.0 (ref. 13) and a standard extracellular pH of 7.4, the concentration gradient is in the right direction. However, oligodendrocytes have a resting membrane potential of around −70 mV,
14
implying that the electrical gradient is in the wrong direction. These combine to give a positive ΔGp (Table 1B; the reversal potential for H+ is −24 mV, more positive than the resting potential), meaning that protons would not tend to flow across the membrane in the right direction (in fact, passively, they would flow in the opposite direction, into the cell) and cannot provide energy for ATP synthesis. In this scenario, the ATP synthase would operate in reverse, hydrolyzing ATP rather than phosphorylating ADP, while H+ ions enter the cell. Thus, if myelin expressed respiratory chain proteins in the manner suggested by Morelli
Ectopic expression of the F1FO-ATP synthase with the F1 segment on the extracellular side of the membrane has been proposed for several cells (reviewed in Champagne
Thus, for mitochondria, and for myelin with the F1 segment of the ATP synthase oriented intracellularly, protons would tend to flow in the right direction for ATP generation. But, for ATP to actually be generated, the energy released by this proton flow must overcome the positive ΔG required to phosphorylate ADP (ΔGATP), which is set by the ratio of substrate (ADP and Pi) to product (ATP) present. Knowing the cytoplasmic and matrix concentrations of ADP, ATP, and Pi, 3 we can evaluate ΔGATP in mitochondria and myelin (Tables 1A and 1C). For mitochondria, the energy provided by the proton motive force is sufficient to overcome the energy required for ATP synthesis if >1.93 protons per ATP flow through the ATP synthase. In fact, 8/3 = 2.7 protons enter per ATP formed (since movement of 8 H+ is required to turn the FO part of the synthase through one rotation which generates 3 ATP molecules 4 ). Consequently, ATP synthesis occurs (since ΔGATP + 2.7 × ΔGp < 0; Table 1A). For the same proton flow (2.7 protons/ATP) across the myelin membrane, on the other hand, even with the F1 segment of the ATP synthase facing intracellularly so that protons would tend to passively flow in the right direction, the proton motive force is not large enough to provide sufficient energy for ATP synthesis (since ΔGATP + 2.7 × ΔGp > 0; Table 1C). In this arrangement, as with the F1 segment facing extracellularly, therefore, the ATP synthase would run backwards, hydrolyzing instead of generating ATP and pumping H+ out of the cell. It would theoretically be possible for the ATP synthase to generate ATP using the same proton motive force if the energy provided by at least 14 protons entering the cytoplasm could be harvested for the synthesis reaction. However, this is far more than the 2.7 protons per ATP molecule normally used 4 and, even if not mechanistically impossible, indicates that powering ATP synthesis by the proton motive force across the myelin membrane would, at the very least, be extremely inefficient.
DISCUSSION
We have assessed the plausibility of an ATP-generating function for myelin at the energetic level. It does not seem possible that enough energy could be provided by the electrochemical gradient for protons across the myelin membrane—no matter which direction the ATP synthase is oriented in—to power the synthesis of ATP. In fact, if the ATP synthase were present in the myelin membrane, then it would function in reverse, hydrolyzing instead of synthesizing ATP.
This analysis is based on known values for the mature oligodendrocyte's resting potential and pH. It is possible that these values, measured at the soma (for the resting potential
14
) and in the absence of proper myelin wraps (for the pH
13
), are drastically different from those of the
We therefore suggest that previous data indicating the presence of the F1FO-ATP synthase in myelin membranes reflect contamination of myelin preparations by mitochondrial membrane (possibly from mitochondria located in the Schmidt-Lanterman incisures or inner/outer myelin tongues 11 ) and nonspecificity of antibody labeling. Furthermore, apparent extracellular synthesis of ATP may, in fact, reflect regulated release of intracellular ATP (reviewed in Corriden and Insel 19 ). Although here we have focused on the hypothesis of ATP production by myelin, the same line of reasoning is worth considering for similar theories of extracellular ATP production in other cells, e.g., endothelial cells 20 and hepatocytes. 18
Finally, it is possible that the F1FO-ATP synthase is expressed ectopically in myelin, but not in an energy-producing capacity. In hepatocytes, for example, ectopic F1FO-ATP synthase expression has a role in regulating the extracellular ADP concentration by hydrolyzing—but not by synthesizing—ATP. 21
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflict of interest.
Footnotes
ACKNOWLEDGEMENTS
The authors thank Lorena Arancibia-Carcamo, Michael Duchen, Renaud Jolivet, and Peter Rich for helpful comments on the manuscript. Our attention was drawn to this topic by an interesting comment from A Morelli and I Panfoli (![]() ) in response to Harris and Attwell.
10
) in response to Harris and Attwell.
10