
Abstract
Elevation of intraluminal pressure increases vasomotor tone, which thought to have a substantial role in regulation of cerebral blood flow (CBF). Interestingly, responses of cerebral vessels to increases in flow varied and have not been studied in human cerebral arteries. We hypothesized that increases in flow elicit constrictions of isolated human and rat cerebral arteries and aimed to elucidate the underlying mechanisms. Human cerebral arteries and rat middle cerebral arteries constricted to increases in flow (
Introduction
Regulation of cerebral blood flow (CBF) is the result of complex, multilevel, interrelated mechanisms to ensure many roles, such as nutritional supply of brain tissue, appropriate gas exchange between blood and tissue, maintenance of intracranial volume and pressure. The metabolic and neural regulation have been well delineated and described by Kontos and others, such as the role of pCO2, adenosine, intrinsic and extrinsic innervations, role of surrounding astrocytes, etc. (Harder et al, 1998; Kontos, 1981; van Beek et al, 2008). In addition, several endothelium-derived factors were shown to contribute to the regulation of CBF (Andresen et al, 2006; Shafi et al, 2008). Moreover, propagated vasodilation was also found to modulate cerebral circulation (Iadecola et al, 1997). Changes in hemodynamic forces are also known to affect CBF. Previous studies showed that increases in pressure elicit increases in resistance of cerebral vessels (Cipolla et al, 2002; Davis and Hill, 1999; Faraci et al, 1989; Heistad and Baumbach, 1992; Kontos, 1981; Osol et al, 2002; Wallis et al, 1996). However, vasomotor responses to increases in flow in isolated cerebral vessels varied: dilation (Drouin et al, 2011; Drouin and Thorin, 2009; Gaw and Bevan, 1993), constriction (Bryan et al, 2001a; Madden and Christman, 1999), and biphasic responses (Garcia-Roldan and Bevan, 1990, 1991; Ngai and Winn, 1995; Shimoda et al, 1996; Shimoda et al, 1998; Thorin-Trescases and Bevan, 1998) were observed. Importantly, no data are available regarding human cerebral vessels preventing the translation of knowledge from vertebrates to humans. In theory, flow-induced dilation would reduce the magnitude of myogenic constriction of cerebral vessels, which would reduce the gain of autoregulation of CBF, whereas if flow elicited constriction, it could contribute to a more efficient autoregulation of CBF.
In the present study, we hypothesized that increases in flow elicit constriction of isolated human and rat middle cerebral arteries and assess the potential contribution of flow-induced response to the autoregulation of CBF and aimed to elucidate the underlying molecular mechanisms.
Materials and methods
Isolation of Intracerebral Arteries of Human and Rat Middle Cerebral Arteries
All procedures were approved by the institutional animal care and use committe of New York Medical College, Valhalla, New York, USA and University of Pecs, Medical School, Pecs, Hungary. Studies of human samples were carried out under the approvement of the Regional Ethic and Review Commeette of the University of Pecs.
Brain samples were obtained from discarded tissues of patients undergoing neurosurgic treatment of epileptic disorder or cerebral tumors (
Flow-, Pressure-, and Simultaneous Flow- and Pressure-Induced Responses of Isolated Cerebral Arteries
After isolation, cerebral arteries were transferred into a custom-made pressure-flow chamber. Inflow and outflow pressures were controlled and measured by a pressure servo-control system (Living Systems Instrumentation, Burlington, VE, USA). Perfusate flow was measured by a ball flow meter (Omega, Omega Engineering Inc., Stamford, CT, USA). The internal diameter was measured by videomicroscopy with a microangiometer (Texas A&M University System). Changes in arterial diameter were continuously recorded digitally by PowerLab system (AD Instruments, Sydney, Australia) connected to a computer for later analysis. The size of glass pipettes used in this study was matched to both each other and the diameter of the vessels to achieve equal resistance (see our previous publications (Racz et al, 2010; Ungvari et al, 1999). In addition, inflow and outflow reservoirs and position of the chamber were built in a symmetrical manner providing equal pressures or generating flow in the presence of constant pressure in the midsection of vessels. By the end of the 60 minutes incubation, the vessels developed a spontaneous myogenic tone in response to 80 mm Hg of intraluminal pressure. First, changes in diameter of cerebral arteries were obtained to stepwise increases in flow elicited by pressure differences (ΔP; established by changing the inflow and outflow pressure to an equal degree, but opposite direction; Δ
Calculations
We have estimated the change in CBF (in arbitrary units) by using diameter values induced by only changes in pressure and then simultaneous changes in pressure and flow using the Hagen-Poiseuille equation (
Administration of Vasoactive Agents and Inhibitors
At 80 mm Hg intraluminal pressure, flow-induced diameter changes of HCA and MCA were repeated in the presence of 20-hydroxyeicosatetraenoic acid (20-HETE) synthesis inhibitor HET0016 (10−6mol/L; Gebremedhin et al, 2000) for 30 minutes. Then, flow-induced diameter responses of MCA were repeated in the presence of cyclooxygenase inhibitor indomethacin 2 × 10−6 mol/L for 30 minutes; thromboxane A2 (TXA2)/prostaglandin H2 (PGH2) receptor (TP receptor) blocker SQ 29,548 2 × 10−6 mol/L for 30 minutes; free-radical scavenger superoxide dismutase, 200 U/ml for 30 minutes; catalase, 130 U/ml for 30 minutes; and TXA2-synthase inhibitor ozagrel 10−5 mol/L for 30 minutes. Afterward, 20-HETE (10−7 mol/L) was directly administered into the vessel chamber and diameter changes were recorded. In a series of experiments in the presence of Δ40 mm Hg adenosine (10−5 mol/L) was added into the chamber. The intact vasomotor function of endothelium and smooth muscle was verified by dilation to administration of adenosine triphosphate (10−5 mol/L). To test the specificity of HET0016, acetylcholine-induced (10−5 mol/L,
Expression of CYP450 4A Proteins in Cerebral Vessels
Western blot analysis of vessels was performed as previously described. In brief, samples of MCAs and gracilis muscle arterioles, as control, isolated from rats were loaded onto a 10% acrylamide sodium dodecyl sulfate gel and electrophoresed at 100 V for 3 hours before being transferred to an Immobilon-P nylon membrane (Millipore, Billerica, MA, USA). The membrane was blocked in 5% milk/TBS/0.5% Tween for 2 hours at room temperature before the antibody was added and the incubation continued overnight at 4°C. The blots were washed and the secondary antibody was added for 2 hours at room temperature. The primary antibodies included anti-cytochrome P450 (1:4000 dilution, #ab22615, Abcam, Cambridge, MA, USA). The blots were washed and a 1:20,000 dilution of the anti-rabbit horseradish peroxidase secondary antibody (Amersham, Buckinghamshire, UK) was used. Bands were visualized using a Thermo Super-Signal West Pico kit (Thermo Scientific Pierce Protein Research Products, Rockford, IL, USA). Care was taken to ensure that band density remained within the linear range of the film and did not saturate the film, by performing exposures of different times. Band density was quantified using AlphaEaseFC software (AlphaInnotech, San Leando, CA, USA).
Detection of Superoxide Formation
Superoxide production was assessed in MCA of rat by the dihydroethidium fluorescence method Ethidium bromide (EB) thought to be specific primarily for superoxide (Fink et al, 2004). Middle cerebral arteries isolated from rat brain were placed in a vessels chamber, cannulated, and incubated in the presence of intraluminal flow generated by Δ40 mm Hg pressure difference in PSS at 37°C for 30 minutes. Some of these experiments were repeated in the presence of HET0016 10−5 mol/L for 60 minutes. Control MCAs were incubated in the same conditions in the absence of intraluminal flow. Then, dihydroethidium (3 × 10−5 mol/L) was added to the vials and incubated for 15 minutes. Afterward, MCAs were washed out with PSS and immersed in an embedding medium. Frozen sections of MCAs were visualized by a digital camera attached to a fluorescence microscope (Olympus BX61WI, Olympus Corporation, Center Valley, PA, USA). Intensity of EB fluorescence of the arterial wall was measured and quantified by Image J software (Image Processing and Analysis in Java, freely available from the National Institutes of Health). Relative EB fluorescence intensity was counted by extracting the intensity of the background. Measurement was repeated five times, and EB fluorescence was presented as intensity/total area.
Statistical Analysis
Statistical analysis was performed by two-way ANOVA followed by a Tukey's
Results
Flow-Induced Responses of Cerebral Arteries and Calculations of CBF
Original records show that increases in flow in the presence of constant pressure (80 mm Hg), elicited by increases in pressure differences caused substantial constrictions of a HCA (from 246 to 160 μm; Figure 1), as a function of time. Summary data (Figure 1) show that increases in flow elicited significant constrictions of vessels (human: from 74 ±4.9% to 63±5%, rat: from 63.8±0.8% to 48.8±1.5% of passive diameter at 80 mm Hg,
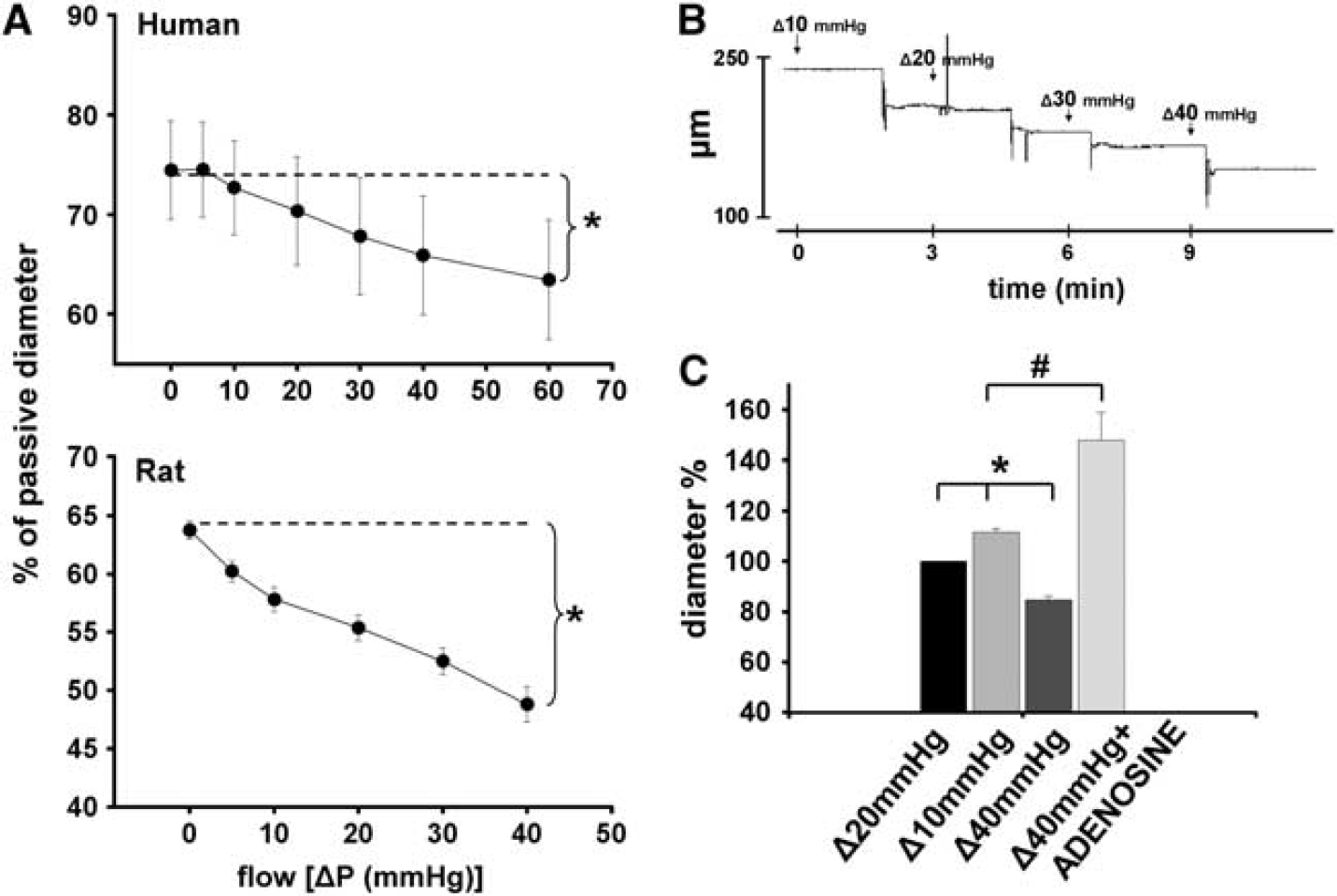
(
We found that increases in intraluminal pressure decreased normalized diameter of MCA (from 84±3% to 53 ± 4%,
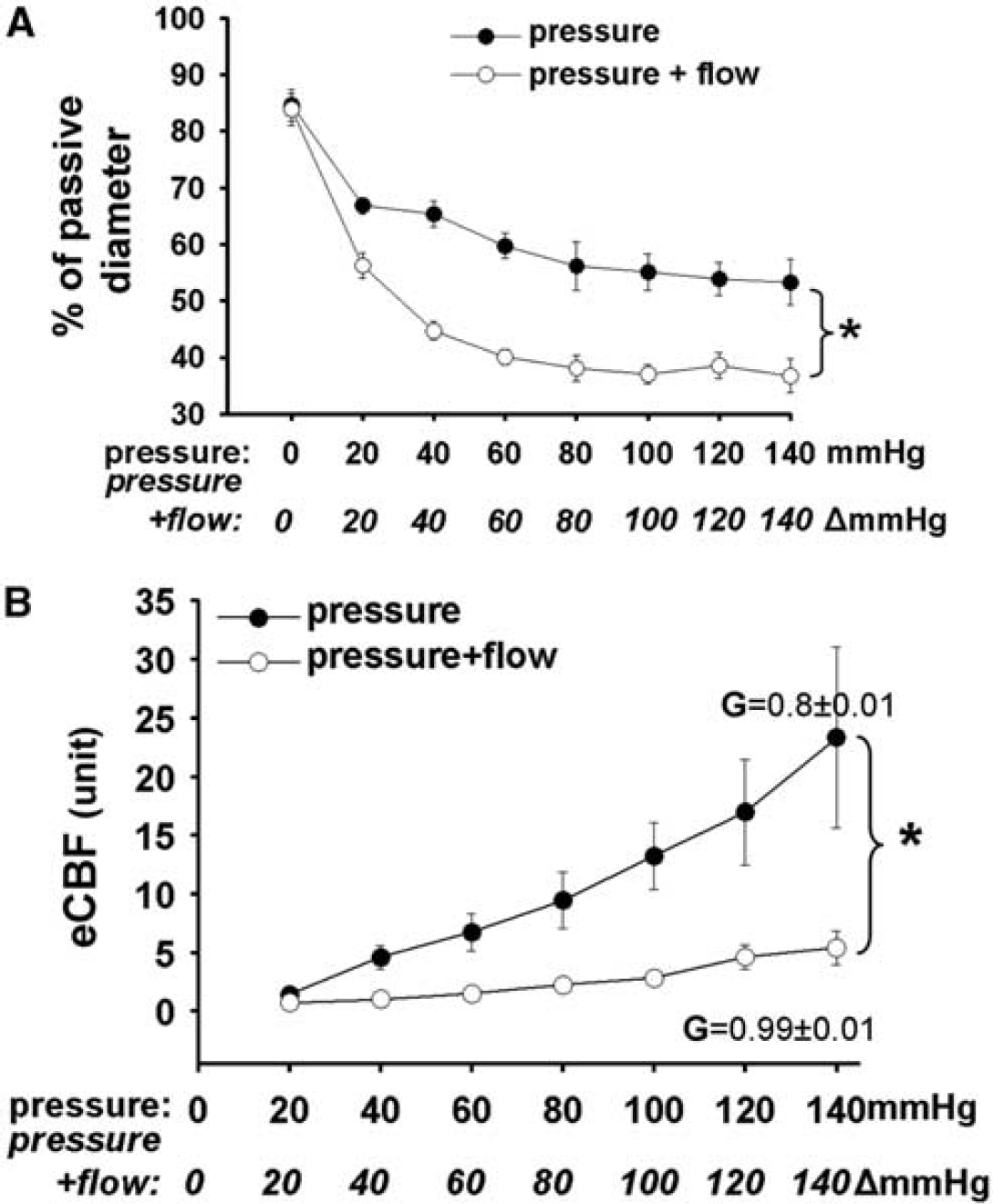
(
When only pressure was increased estimated CBF (eCBF) showed a linear increase from 1.4±0.1 to 23.3±7.6 in arbitrary units. In contrast, when pressure + flow increased simultaneously first the eCBF decreased significantly to 0.7±0.1 and then increased only to 5.4±1.4 in arbitrary unit (Figure 2B). The gain of autoregulation (
Mechanism of Flow-Induced Response of Cerebral Arteries
Summary data show that incubation of the vessels with HET0016 abolished the decrease in diameter of both HCA and MCA elicited by increases in flow (Figure 3A). Direct administration of 20-HETE (10−7mol/L) decreased the diameter of MCA similarly to flow (at Δ
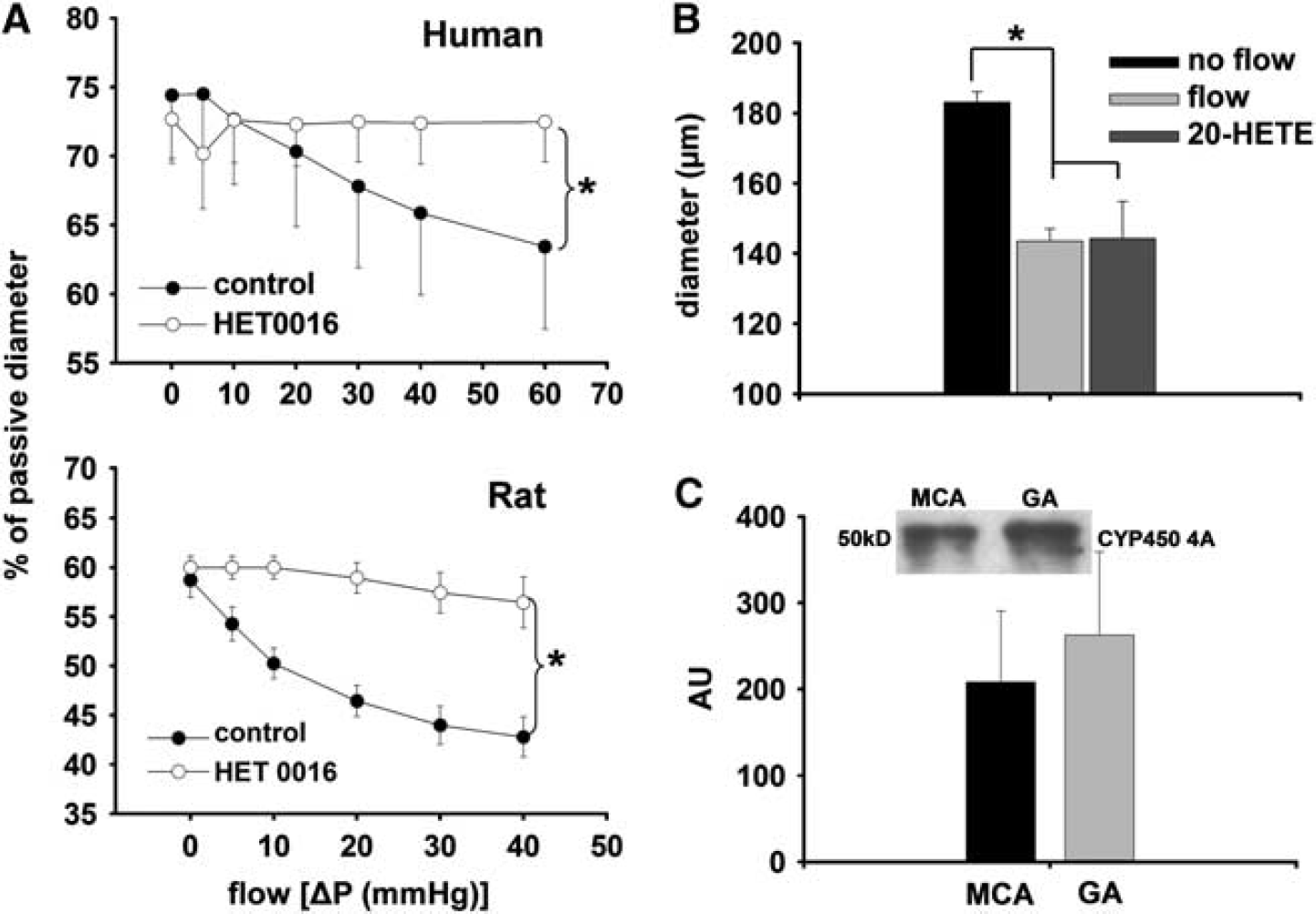
(
Dilations of MCA in response to acetylcholine (ACh) were not affected significantly by HET0016 (before: 53 ±4.6%, after: 46 ±5.4% of passive diameter). Figure 3C shows that cytochrome P450 4A enzymes are present in the MCA of rat.
Incubation of the vessels in superoxide dismutase/catalase significantly decreased the reduction in diameter of rat cerebral arteries elicited by increases in flow. Representative EB fluorescent images of sections of MCA and summary data show an enhanced EB fluorescence in the vessels exposed to flow compared with control (absence of flow). Enhanced EB fluorescence was reduced to the control level by HET0016 (10−6m/L; control: 0.05 ±0.02, flow: 0.18 ±0.04, flow + HET0016: 0.07 ±0.02 integrated intensity/total area, respectively;
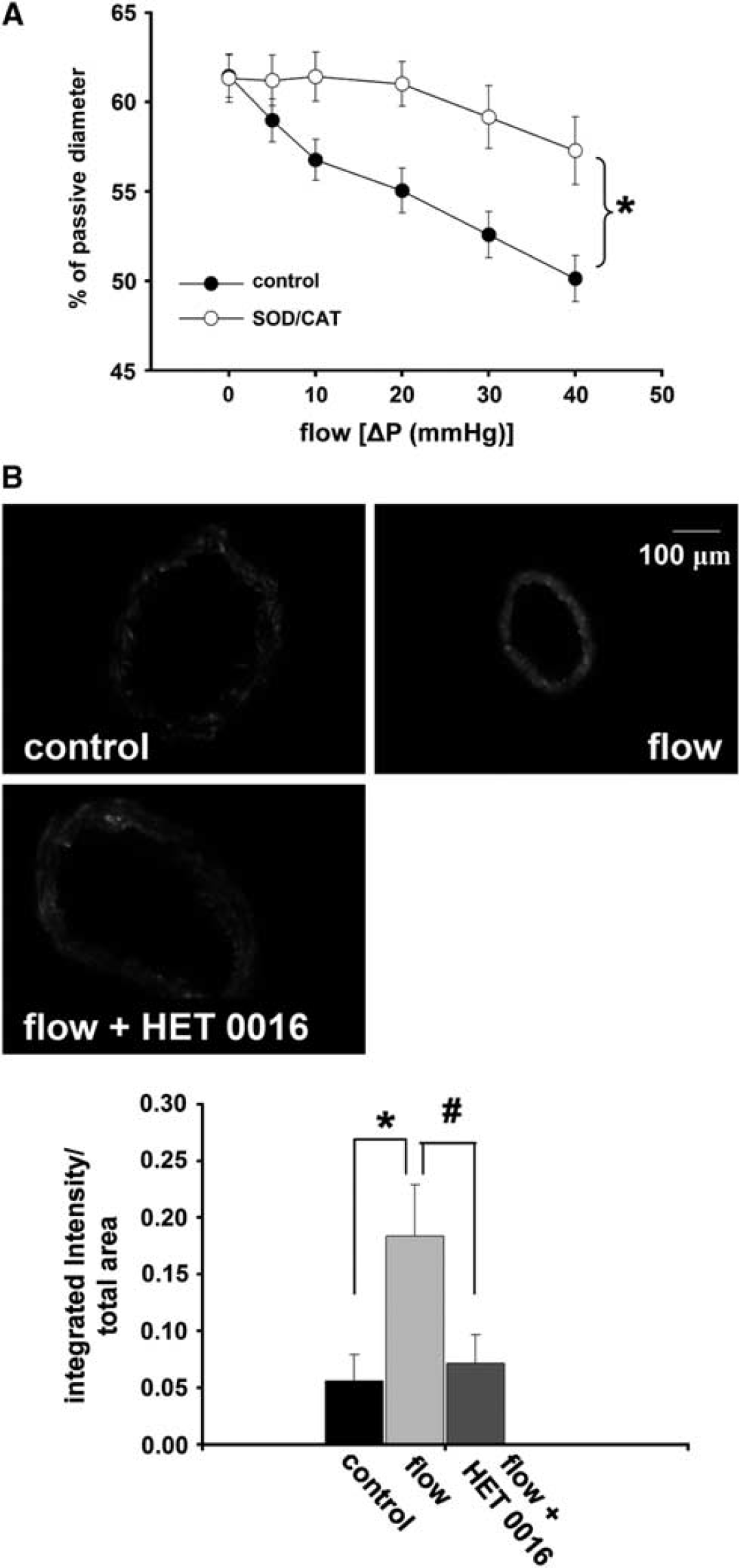
(
Summary data show that incubation of the vessels with indomethacin or SQ 29,548 inhibited the constriction of MCA to increases in flow, whereas ozagrel did not have an effect (Figure 5A and B).
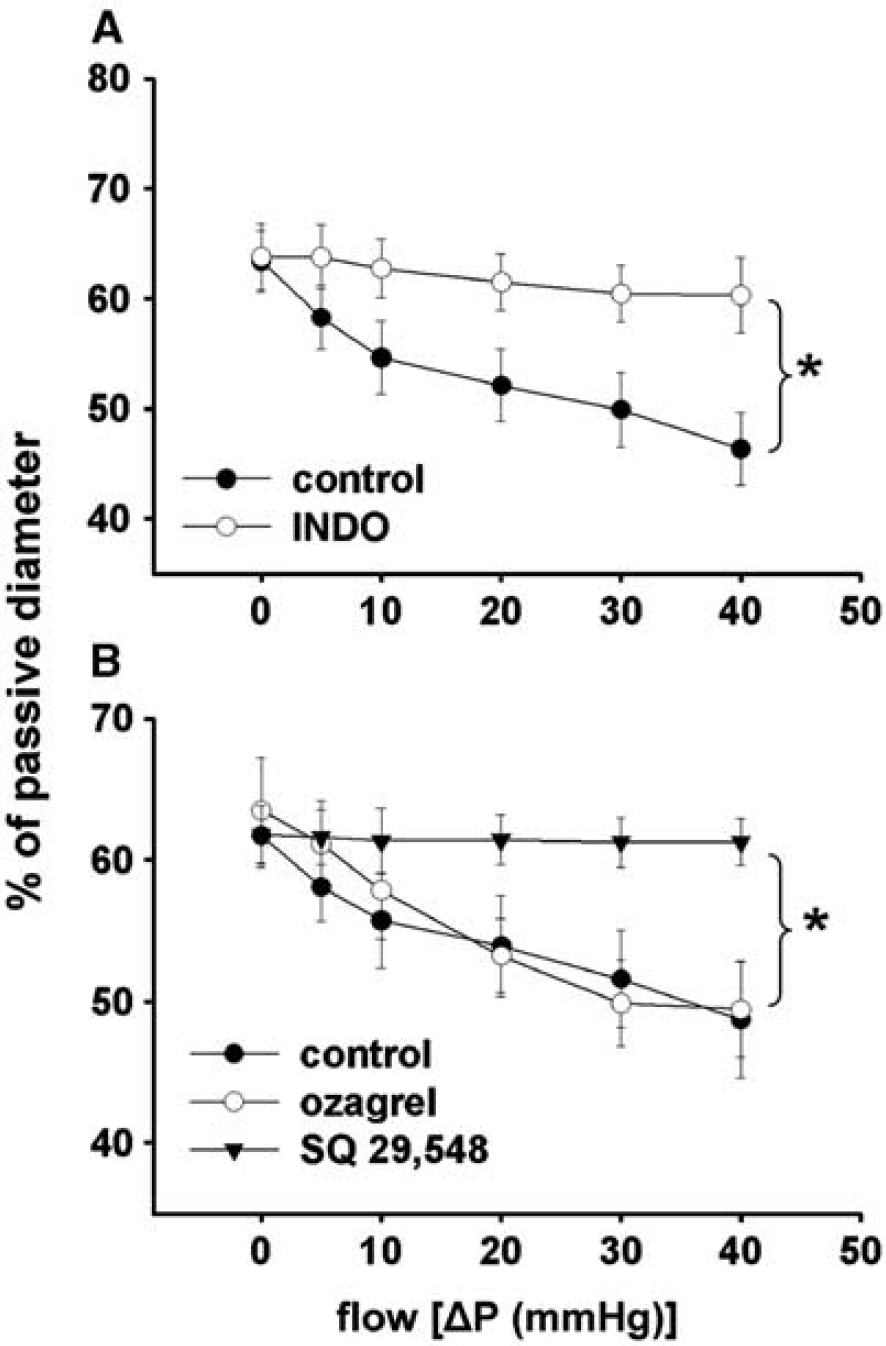
(
Discussion
The novel findings of the present study are: (1) increases in flow elicit constrictions in isolated human intracerebral arteries and rat middle cerebral arteries, (2) simultaneous increases of pressure and flow elicited significantly greater constriction than pressure alone, (3) the underlying mechanism of flow-induced constriction of cerebral arteries involves increased production of reactive oxygen species (ROS), activity of COX, and mediated by 20-HETE acting via TP receptors.
Flow-Induced Constriction of Isolated Cerebral Arteries and its Contribution to an Effective Autoregulation of CBF
At present, the myogenic vasomotor mechanism is used primarily to explain the presence of autoregulation of CBF, which aims to protect the brain from high volume and pressure (Faraci et al, 1989; Harper et al, 1984; Mellander, 1989; Mueller et al, 1977). Much less is known regarding the role of flow-dependent regulation of CBF, although
In peripheral vessels, the vascular response to increases in flow is dilation (Koller et al, 1993). In previous studies, flow-induced responses of isolated cerebral vessels varied: depending on the prevailing pressure, conditions and species dilations, constrictions and biphasic responses were observed (Bryan et al, 2001a; Bryan et al, 2001b; Fujii et al, 1991; Garcia-Roldan and Bevan, 1990,; Gaw and Bevan, 1993; Madden and Christman, 1999; Ngai and Winn, 1995; Paravicini et al, 2006; Shimoda et al, 1996). Importantly, no studies have been conducted to elucidate the nature of responses of isolated human cerebral arteries to increases in flow.
In theory, if cerebral arteries dilated to flow it would reduce the magnitude of myogenic constriction. Also, it was found that in isolated large cerebral arteries, which are responsible for significant part of total cerebrovascular resistance (in contrast to peripheral vascular beds) (Faraci et al, 1988; Kontos et al, 1978; Stromberg and Fox, 1972), in the physiologic range of perfusion pressure (∼40–140 mm Hg) increases in pressure do not elicit substantial reduction in diameter (Osol et al, 2002; Wallis et al, 1996), which would allow an increase in CBF. However,
Here, we showed for the first time that isolated HCAs constrict to increases in flow and confirmed similar findings of Bryan et al, (2001a) in rat MCA (Figure 1). Also, pressure + flow together caused a significantly greater constriction of cerebral arteries than pressure alone (Figure 2). By estimation (e) of CBF and the gain of autoregulation we aimed to extrapolate these
Whereas flow-induced constriction (together with the pressure sensitive myogenic response) may have an important role in the regulation and maintenance of cerebral volume and intracranial pressure, increased local neural activity/function (and consequent cellular metabolic changes) can increase the diameter of cerebral vessels adjacent to the neural cells and thus increase local CBF (Ingvar, 1976; Kontos, 1981; Sokoloff, 1977; van Beek et al, 2008).
This idea is in line with previous findings showing that metabolic effects (such as hypercapnia, acidosis, hypoxia, and adenosine) can override the vasoconstrictor effects of pressure (Raisis et al, 1979) and with the finding of the present study demonstrating that flow-induced constriction could be converted to dilation by adenosine, a key molecule of metabolic regulation (Figure 1). In addition, as shown by Iadecola et al, (1997) increased synaptic activity elicited local functional hyperemia of cerebral arterioles, which is then propagated to upstream vessels via intravascular mechanisms. Thus,
The present study aimed to investigate one vasomotor mechanism intrinsic to the vascular wall, thus the result should be carefully related directly to
In conclusion, we propose that during increases in systemic pressure the pressure- and flow-sensitive constrictor mechanisms ‘set’ the vasomotor tone, which can be modulated or overridden by other factors sensitive to the needs of neural tissues. Thus CBF is regulated by multilevel and redundant mechanisms to adapt to the lack of one or two mechanisms.
Mechanisms Mediating Flow-Induced Constriction of Cerebral Arteries
The mechanisms of flow-induced constriction of cerebral vessels are still not clarified. Arachidonic acid and its metabolites (such as CYP450 4A-derived 20-HETE, COX-derived TXA2,) have important roles in the regulation of cerebrovascular resistance (Dunn et al, 2008; Ellis et al, 1977; Gebremedhin et al, 2000). Harder et al, (1994) and Gebremedhin et al, (1998) showed that AA is metabolized by cytochrome P450 ω-hydroxylases (CYP450 4A) into 20-HETE and it has an important role in the regulation of cerebrovascular tone, by mediating agonists- and pressure-induced constrictions of vascular smooth muscle of cerebral vessels (Gebremedhin et al, 2000; Yu et al, 2004). Thus, it seemed to be logical to hypothesize that 20-HETE could be the constrictor factor mediating the flow-induced constriction of cerebral arteries as well. Indeed, we found that flow-induced constrictions of HCAs and MCAs of rat were abolished by administration of HET0016 (Figure 3), an inhibitor of 20-HETE production. Consistently to these functional findings we found that CYP450 4A enzymes are present in the MCA of the rat, a finding similar to that of Dunn et al, (2008) and Gebremedhin et al, (2000).
It is also known that direct administration and production of 20-HETE by cytochrome P450 can produce ROS (Terashvili et al, 2006). Thus, we hypothesized that in response to flow ROS are produced as well, and scavenging ROS would affect the constrictor response. We found that administration of ROS scavengers significantly reduced the flow-induced constriction of cerebral arteries (Figure 4). In addition, our findings showed increased EB fluorescence in MCA after exposing the vessels to flow, suggesting flow-induced increased ROS production. The enhanced production of ROS was reversed by inhibition of 20-HETE production (Figure 4), suggesting ROS is generated during synthesis of 20-HETE, which is elicited by increases in flow. This finding also confirms that ROS are generated during activation of CYP450 4A (Terashvili et al, 2006). Although HET0016 abolished flow-induced constriction, ROS likely have direct vasomotor effect, as well.
Previous studies (Harder et al, 1997; Zou et al, 1996) have proposed that 20-HETE constricts cerebral arteries by various pathways. For example, 20-HETE activates protein kinase C, depolarizes smooth muscle cells by inhibition of large-conductance KCa channel, and increases Ca2+ influx via L-type Ca2+ channels.
In addition, thromboxane A2 is another constrictor metabolite of AA and known to have a profound effect on cerebrovascular tone (Ellis et al, 1977). Thus, we evaluated the effect of TXA2 synthase inhibitor ozagrel, and TP receptor blocker SQ 29,548. Importantly, SQ 29548 abolished the flow-induced constriction, but ozagrel did not have any effects (Figure 5). Our results showing that both inhibition of 20-HETE production and antagonizing TP receptor abolished flow-induced constriction and suggest that 20-HETE may act on TP receptor. Consistently with this hypothesis, previous studies by Escalante et al, (1989) and Schwartzman et al, (1989) proposed that 20-HETE caused constriction of arteries via TP receptor after 20-HETE was metabolized by COX into 20-endoperoxides (20-OH-PGH2, 20-OH-PGG2). This finding is supported by our finding that indomethacin also blocked flow-induced constrictions of MCA. The proposed molecular mechanisms mediating flow-induced constriction of human and rat cerebral arteries are summarized in Figure 6.
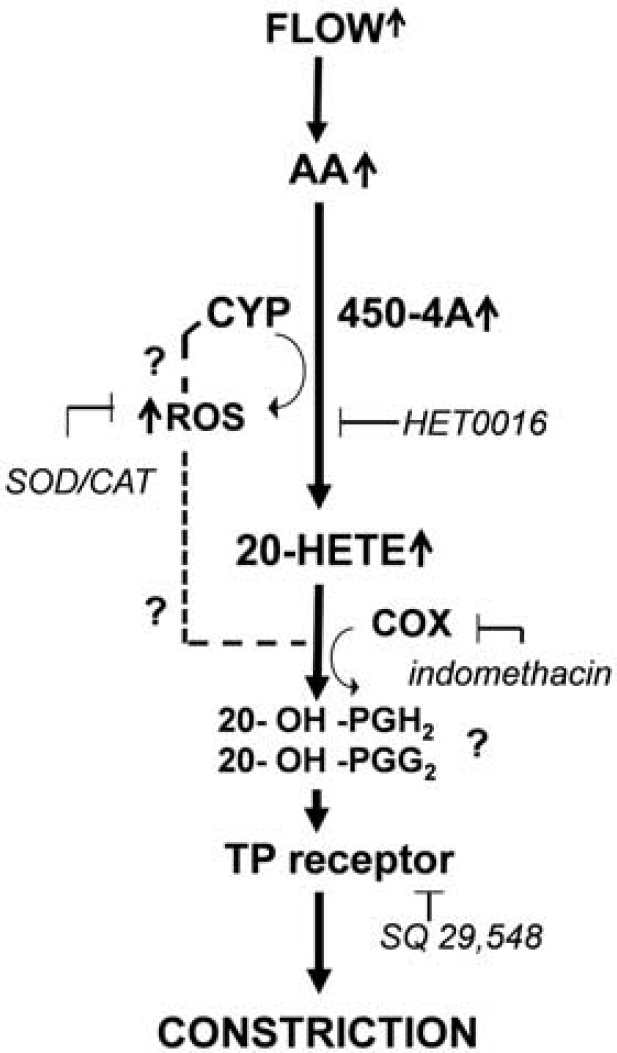
Proposed mechanisms of flow-induced constrictions of cerebral arteries. Increases in flow activate arachidonate production, which is further metabolized by cytochrome P450 4A enzymes (CYP450 4A) into 20-hydroxyeicosatetraenoic acid (20-HETE). CYP450 4A also produces reactive oxygen species (ROS), which contribute to the constriction. The flow-induced constriction is mediated via thromboxane A2/prostaglandin H2 (TP) receptors and requires COX activity.
Interestingly,
In conclusion, the novel findings of the present study are: (1) increases in flow elicit constrictions in isolated human intracerebral arteries and rat middle cerebral arteries, (2) simultaneous increases of pressure and flow elicited significantly greater constriction than pressure alone, (3) the underlying mechanism of flow-induced constriction of cerebral arteries involves increased production of ROS, activity of COX, and mediated by 20-HETE acting via TP receptors.
Footnotes
Acknowledgements
We thank Dr John G. Edwards, and Jamie Mathew for performing the western blot measurements.
The authors declare no conflict of interest.