
Abstract
The choice of reference region in positron emission tomography (PET) human brain imaging of the vesicular monoamine transporter 2 (VMAT2), a marker of striatal dopamine innervation, has been arbitrary, with cerebellar, whole cerebral, frontal, or occipital cortices used. To establish whether levels of VMAT2 are in fact low in these cortical areas, we measured VMAT2 protein distribution by quantitative immunoblotting in autopsied normal human brain (
Keywords
Introduction
The vesicular monoamine transporter 2 (VMAT2), a member of the solute carrier family 18 (SLC18A2) with 12 transmembrane domains, is the protein responsible for transporting monoamine neurotransmitters (dopamine, noradrenaline, serotonin) into synaptic vesicles for subsequent storage and release (Erickson et al, 1996). Tetrabenazine or its metabolite dihydrotetrabenazine (DTBZ) is a specific inhibitor of the transporter used in radioligand-binding studies to measure the
A practical issue with PET VMAT2 imaging is selection of a reference region for the measurement of free and nonspecific binding (Chan et al, 1999). In principle, an ideal reference region should have identical free and nonspecific properties as compared with regions of interest with specific binding, a reasonably high uptake of the radiotracer, and a negligible contribution of specific binding to the tracer uptake in this region, that is, very low levels of the protein target. Among the PET VMAT2 studies, the selection of reference region has been arbitrary, with cerebellar (Chan et al, 1999), whole cerebral (Gilman et al, 1998, 1999), frontal (Frey et al, 1996; Gilman et al, 1996), parieto-occipital (Troiano et al, 2010), or occipital cortices (Chan et al, 1999; Koeppe et al, 1999; Boileau et al, 2008; Okamura et al, 2010) having been used. Currently, the occipital cortex appears to be a preferred reference region in human brain imaging studies apparently because the data obtained are less variable and of consistently higher values as compared with those using cerebellar cortex as the reference region (Chan et al, 1999). However, animal studies have repeatedly showed that the cerebellar cortex contains less DTBZ-binding sites than cerebral cortices (Scherman et al, 1988; Darchen et al, 1989; Vander Borght et al, 1995) and in the development of new DTBZ-based PET tracers the cerebellum, for practical reasons, continues to be used as the reference region in preclinical studies (Kilbourn et al, 2007).
To address the question of the most appropriate reference region for VMAT2 PET imaging, we reasoned that it would be helpful to know the relative concentrations of VMAT2 protein in different regions of the human brain. The current study therefore used quantitative VMAT2 immunoblotting in autopsied human brain. Given that several VMAT2-immunoreactive species of uncertain identity have been reported in the literature (Wang et al, 1997; Jassen et al, 2005; Cruz-Muros et al, 2008), we also made efforts to confirm these VMAT2 Western blot bands.
Materials and methods
Subjects
This study was approved by the Research Ethics Board of the Centre for Addiction and Mental Health at Toronto. A total of six (4M/2F) autopsied brains from neurologically normal subjects (age: 48 ± 0.3 years; postmortem interval: 16 ±3 hours; mean ± s.e.m.) were used in the VMAT2 regional distribution study. To help characterize the VMAT2-immunoreactive protein bands (which should be reduced in intensity in degenerative nigrostriatal dopamine deficiency disorders) in the Western blots, autopsied brains were also obtained from a total of 10 patients with PD (5M/5F), 10 patients with progressive supranuclear palsy (PSP; 9M/1F), 9 patients with multiple system atrophy (MSA; 5M/4F), and 10 normal subjects (4M/6F). No significant difference (one-way analysis of variance) was found among the four groups in postmortem interval (control: 12 ± 1; PD: 14 ± 2; PSP: 11 ± 2; MSA: 14 ± 2hours) or in age (control: 70 ± 3; PD: 75 ± 2, PSP: 73 ± 4; MSA: 64 ± 4 years). One-half brain was used for neuropathological examination, whereas the other half was frozen for neurochemical analyses. The characteristics and pathological findings of the patients have been previously published (Tong et al, 2010). The causes of death for the neuropathologically normal control subjects were cardiovascular illnesses (
Tissue Sample Preparation, Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis, and Western Blotting
Cerebral cortical subdivisions were excised according to Brodmann classification. Dissection of the subcortical areas from the ∼ 3 mm thick coronal sections followed published procedures (Kish et al, 2005; Tong et al, 2010). Brain tissue homogenates were used throughout this study. Sample preparation, sodium dodecyl sulfate polyacrylamide gel electrophoresis, and Western blot followed published procedures (Kish et al, 2005). Protein concentration was determined using the Bio-Rad Protein Assay Kit (Bio-Rad, Hercules, CA, USA) with bovine plasma albumin as the standard. Five concentrations of tissue standard (0.25 to 3 μg of protein), consisting of a pooled human striatal samples, were run on each blot together with the samples (2 to 40 mg of protein, depending on regional levels of VMAT2; see Figure 4). As VMAT2 has a strong propensity to aggregate during heating or long-term storage in sodium dodecyl sulfate polyacrylamide gel electrophoresis sample buffer in the freezer, the samples were prepared by incubation at room temperature for 30 minutes immediately before use. The antibody used for quantitative determination of VMAT2 levels was a well-characterized (Miller et al, 1999; Haycock et al, 2003) rabbit polyclonal antibody raised against the 19-amino-acid C-terminus peptide of human VMAT2 (AB1767; Chemicon International, Temecula, CA, USA). For measurement of the level of the ∼ 35 k Da VMAT2 band IV (see Figures 1–4), blots were cut between the two colored molecular weight (MW) markers (47.5 and 32.5 kDa), with the upper and lower half probed with the primary antibody at a dilution of 1:10,000 and 1:3,000, respectively. The same dilution of 1:10,000 was used for the goat anti-rabbit IgG (H + L) horseradish peroxidase secondary antibody (cat# 4050-05; Southern-Biotech, Birmingham, AL, USA). Another recently available anti-human VMAT2 monoclonal antibody (clone 9E11, cat# TA500506, OriGene Technologies, Rockville, MD, USA) raised against the recombinant, full-length VMAT2 protein was also used to help characterize the VMAT2-immunoreactive protein bands. Western blot of the ‘control’ protein neuron specific enolase followed published procedures (Kish et al, 2005).
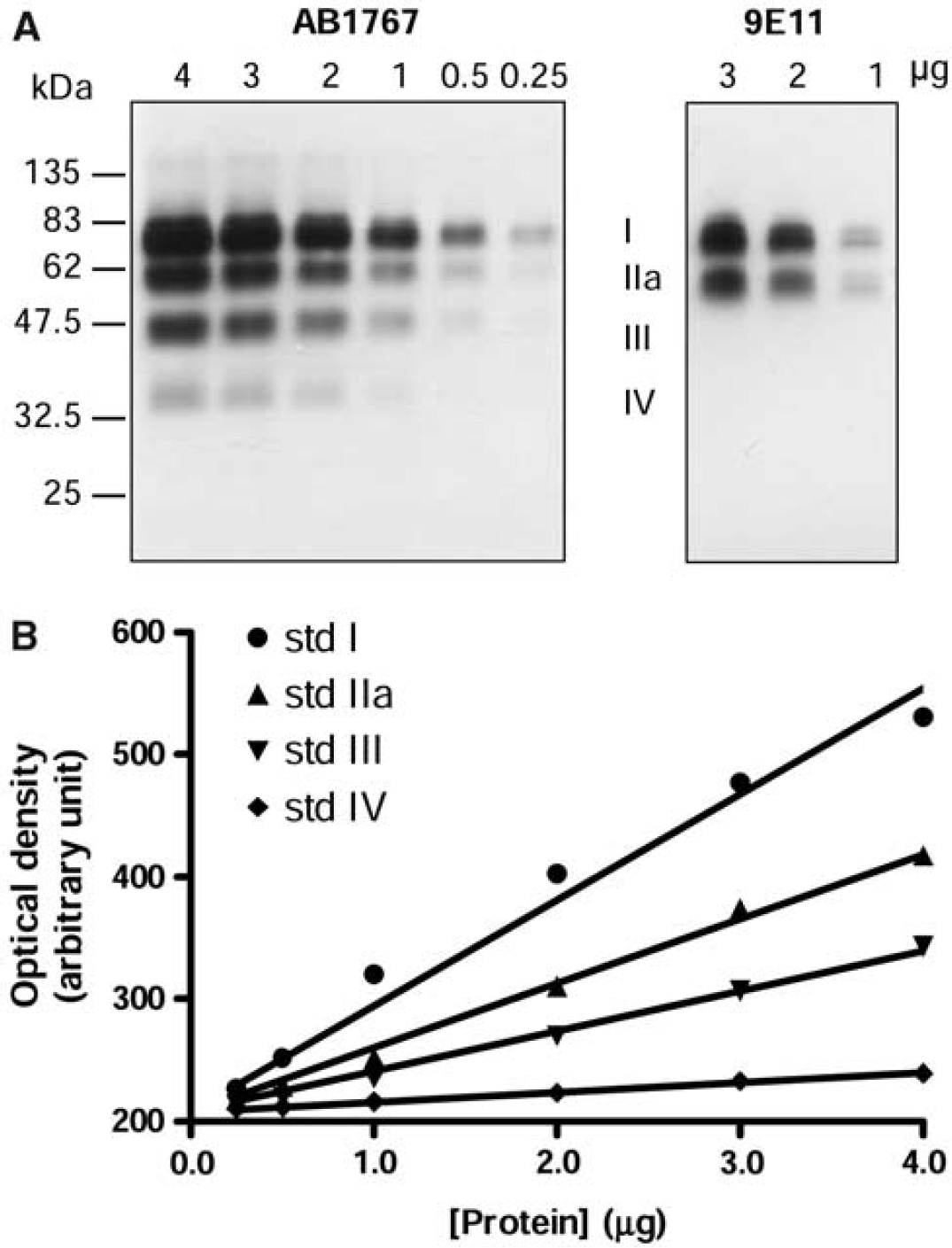
Western blotting of vesicular monoamine transporter 2 (VMAT2) in the striatum of autopsied human brain. (
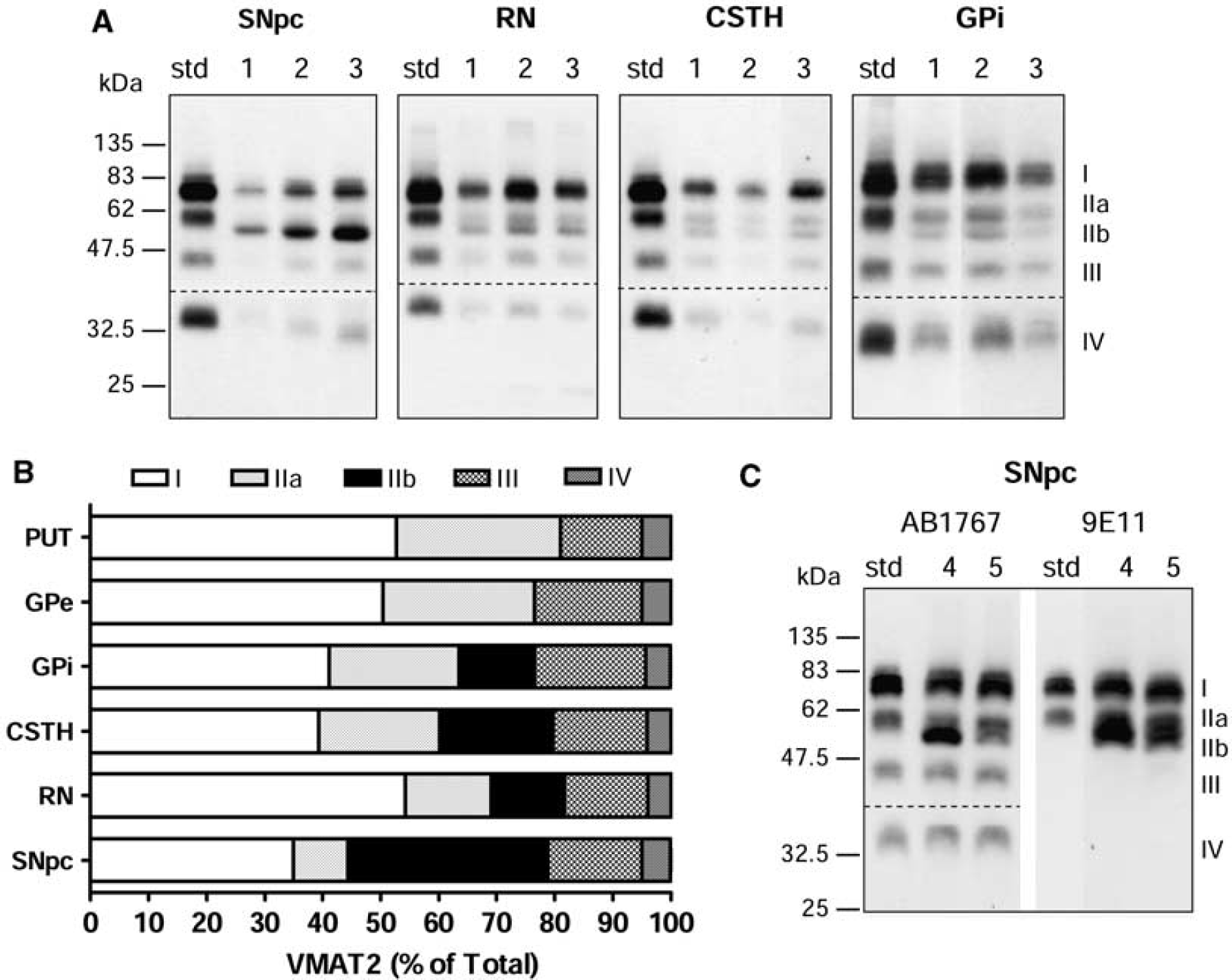
Vesicular monoamine transporter 2 (VMAT2) band IIb in the substantia nigra and neighboring regions of autopsied human brain. (
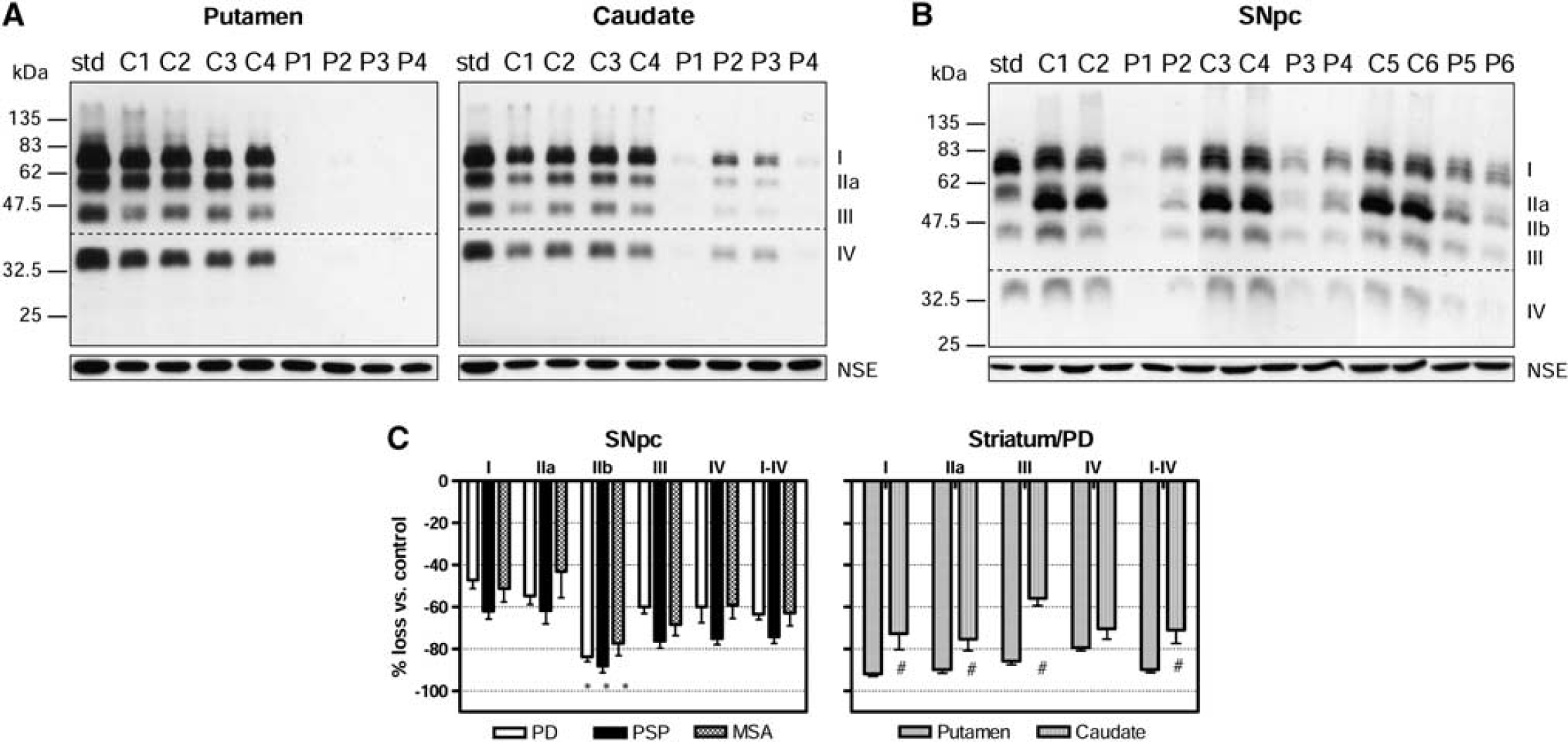
Vesicular monoamine transporter 2 (VMAT2) in the brain of patients with Parkinson's disease (PD), progressive supranuclear palsy (PSP), and multiple system atrophy (MSA). (

Representative immunoblots of vesicular monoamine transporter 2 (VMAT2) in 37 brain regions of a normal control subject. ‘A’ refers to Brodmann cerebral cortical areas; A23, cingulate gyrus posterior; A24, cingulate gyrus anterior; A25, parolfactory gyrus; CCc, corpus callosum caudal; CCr, corpus callosum rostral; Cereb, cerebellar cortex; CN, caudate; CNA, hippocampal Ammon's horn; CSTH, subthalamic nucleus; GD, dentate gyrus; GH, hippocampal gyrus; GPe, globus pallidus external; GPi, globus pallidus internal; GUNC, gyrus of uncus; Hypothal, hypothalamus; ICr, internal capsule rostral; Insular, insular cortex; MDTH, mediodorsal thalamus; N. basalis, nucleus basalis; NAM, amygdala; NAV, anterior ventral nucleus of thalamus; NL, nucleus lateralis of thalamus; NLV, lateral ventral nucleus of thalamus; NPM, medial pulvinar of thalamus; PUT, putamen; RN, red nucleus; SBI, substantia innominata; SNpc, substantia nigra pars compacta.
Data Analyses
Four linear standard curves were constructed for each of the VMAT2 protein bands (I, IIa, III, and IV; see Figure 1B for an example) on each blots. The relative levels of the four VMAT2 protein bands in the tissue standard were determined by ratios of the slopes of the standard curves of band IIa, III, and IV to that of band I, that is,
Differences in VMAT2 levels among control and parkinsonian patients were assessed by one-way analysis of variance followed by
Results
Characterization of Vesicular Monoamine Transporter 2-Immunoreactive Protein Bands in Western Blots of Normal Autopsied Human Striatum
The polyclonal antibody raised against a 19-amino-acid C-terminus peptide of human VMAT2 (AB1767 from Chemicon) has been widely used to examine VMAT2 protein by Western blot in striatum of human brain (Miller et al, 1999; Haycock et al, 2003; Little et al, 2003) and in the brain of rodents under a variety of conditions (Wang et al, 1997; Gainetdinov et al, 1998; Cruz-Muros et al, 2008). As expected (Haycock et al, 2003), the antibody detected in human striatal samples a total of four broad protein bands with an MW of ∼ 75 (I), 55 (IIa), 45 (III), and 35 kDa (IV), respectively (Figure 1A). The ∼ 75 k Da VMAT2 I was the predominant band, accounting for 53% ± 2% (
To address the literature controversy with respect to the identity of the multiple VMAT2-immunoreactive bands detected in the striatum (Wang et al, 1997; Gainetdinov et al, 1998; Miller et al, 1999; Jassen et al, 2005; Cruz-Muros et al, 2008), we performed immunoblotting with another recently available monoclonal antibody (9E11) raised against recombinant, full-length human VMAT2. As shown in Figure 1A, the monoclonal antibody detected in human striatal samples only the two larger forms (I and IIa) of VMAT2, with the relative intensity of the two bands also similar to that observed with the polyclonal antibody. Therefore, our observations with two antibodies against different epitopes of VMAT2 provide some support for the notion that the minor VMAT2-immunoreactive species at 45 and 35 k Da might be N-terminally truncated forms (Wang et al, 1997; Gainetdinov et al, 1998; Miller et al, 1999; Jassen et al, 2005).
Distinct Vesicular Monoamine Transporter 2 Immunoreactivity in the Normal Human Substantia Nigra
The four-band pattern (I, IIa, III, and IV) of VMAT2 detected with the polyclonal antibody was generally observed throughout the human brain regions examined (see Figure 4). However, we found that another protein band (∼ 52 kDa, IIb) of slightly smaller MW than the 55-kDa band could be detected by the polyclonal antibodies in substantia nigra pars compacta, the neighboring midbrain regions red nucleus and subthalamic nucleus, and in the internal portion of the globus pallidus (GPi) (Figure 2A, see also Figures 3B and 4). Moreover, in the substantia nigra, the 52-kDa band IIb was of (at least) comparable intensity as that of the mature 75 k Da band I (35% ± 1% versus 35% ± 2% of total levels of VMAT2, respectively; Figure 2B), with levels of the 55-kDa band IIa low in the dopamine cell body region (9% ± 1%). The 75-kDa band I nevertheless was predominant in other brain regions. The percentage levels of band I, IIa, and IIb were 54% ± 2%, 15% ± 1%, and 13% ± 2% in the red nucleus, 39% ± 3%, 21% ± 2%, and 20% ± 1% in the subthalamic nucleus, and 41% ± 2%, 22% ± 1%, and 13% ± 1% in the GPi. We also confirmed that the 52-kDa band IIb in the substantia nigra was immunoreactive to the monoclonal antibody (Figure 2C).
Vesicular Monoamine Transporter 2 Immunoreactivity Is Decreased in Substantia Nigra and Striatum of Patients with Degenerative Parkinsonian Conditions
To provide further evidence that the protein bands (I, IIa, IIb, III, and IV) detected by the antibodies were in fact derived from VMAT2, we examined brain of patients with PD, the classic degenerative dopamine, and VMAT2 deficiency condition (Frey et al, 1996; Wilson et al, 1996). As expected, in the striatum (putamen and caudate) of patients with PD, levels of all four VMAT2 bands I, IIa, III, and IV were markedly and evenly decreased, with the loss significantly more severe in putamen (−92% ± 1%, −90% ± 2%, −86% ± 2%, and −79% ± 2% for bands I to IV, respectively) than in caudate (−73% ±8%, −75% ± 6%, −56% ± 3%, and −70% ± 5%, respectively; Figures 3A and 3C). The overall loss of VMAT2 immunoreactivity was −90% ± 2% and −71% ± 6% for putamen and caudate, respectively, consistent with the literature showing that the putamen has more severe dopamine marker loss in PD than the caudate (Frey et al, 1996; Wilson et al, 1996). Similarly in the substantia nigra, levels of all five VMAT2-immunoreactive bands were significantly reduced (−47% ± 4%, −55% ± 4%, −84% ± 2, −60% ± 3, and −60% ± 8%, respectively) in PD although to a less overall extent (−63% ± 3%) than those in the striatal terminal regions (Figures 3B and 3C). Again, this is in line with the pathological finding that the loss of dopamine cell body is less severe than the loss of dopamine terminals in the striatum in PD (cf. Hornykiewicz, 1998). Interestingly, we found that the 52-kDa band IIb was preferentially lost as compared with the other bands (−84% versus −47% to −60%;
Levels of neuron specific enolase, a control protein, were normal in the putamen, caudate, and substantia nigra of patients with PD (Figures 3A and 3B). However, neuron specific enolase levels in the substantia nigra were significantly decreased in patients with PSP (−47%) and MSA (−33%).
Regional Distribution of Vesicular Monoamine Transporter 2 in Normal Human Brain
A total of 37 brain regions covering the entire human forebrain and including both primarily dopaminergic and nondopaminergic areas were examined (see Figure 4 for representative immunoblots). It should be mentioned that with higher loading of protein, additional protein bands including some high MW bands and two sharp bands immediately above and below the 35-kDa VMAT2 band IV could be observed with the polyclonal antibody, particularly in thalamus and cerebral cortices. However, these protein bands appeared to be nonspecific reaction products as, unlike VMAT2 proteins, they were enriched in the soluble fraction. All four or five VMAT2 protein bands were measured quantitatively and normalized against putamen, the brain region with highest levels of VMAT2. As shown in Figure 5, the regionally normalized data for levels of mature VMAT2 (band I), intact VMAT2 (bands I + II, including both IIa and IIb), and total VMAT2 were not significantly different (region × measure: F72,540 = 0.09 5,
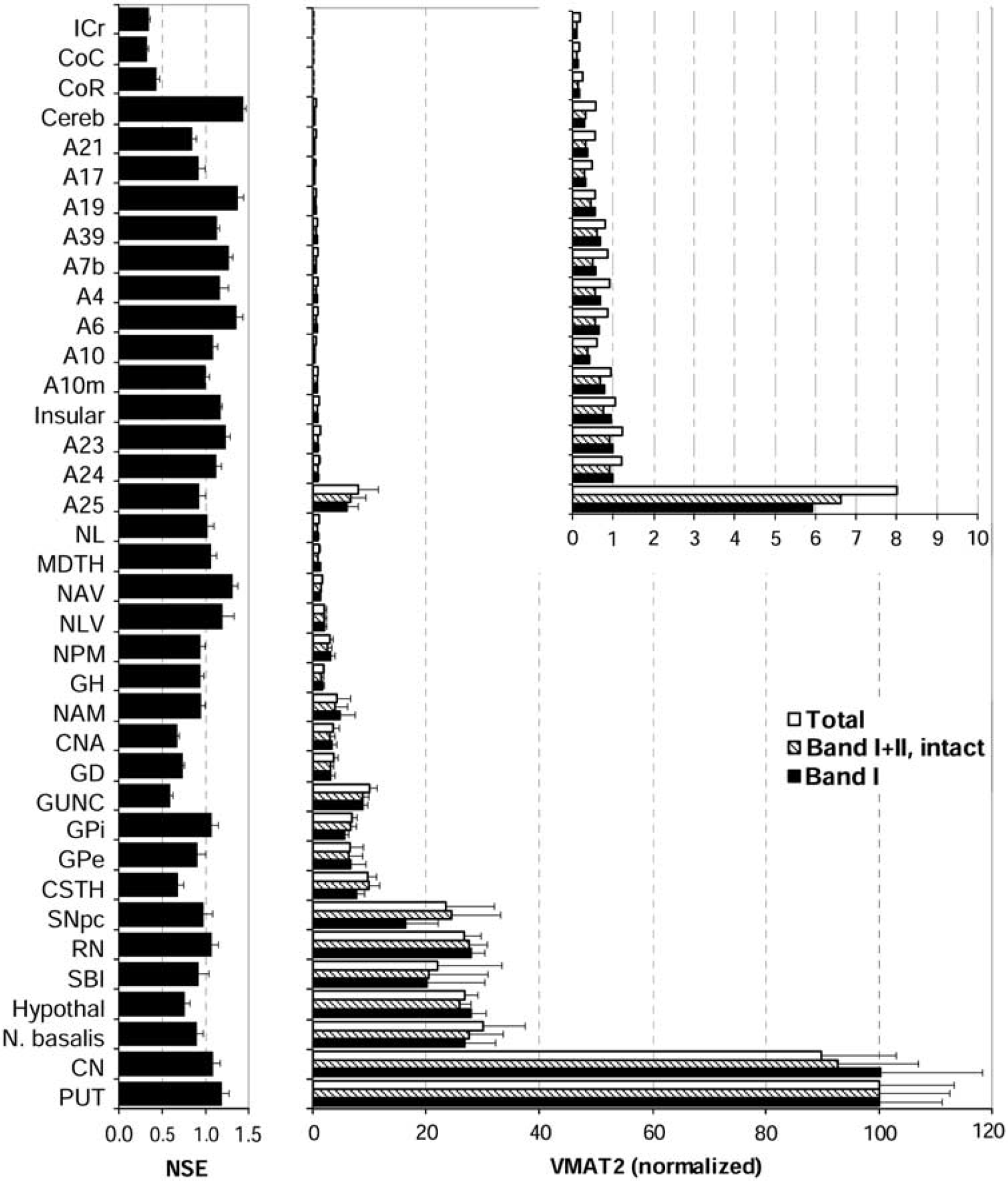
Regional brain distribution of vesicular monoamine transporter 2 (VMAT2) in normal human brain (
The brain gray matter regions could be separated into four groups according to VMAT2 protein levels. The highest levels of VMAT2 were in the striatum and were at least threefold higher than that of any other brain region. Concentrations of VMAT2 in putamen were somewhat higher than in caudate although the difference was not statistically significant. The basal forebrain regions including nucleus basalis, hypothalamus, and substantia innominata and the midbrain regions substantia nigra pars compacta and red nucleus had moderate concentrations of VMAT2 that were 20% to 30% of that of putamen. The other subcortical brain structures together with a few allocortical/archicortical areas, including the basal ganglia output regions subthalamic nucleus, globus pallidus external (GPe) and GPi, the medial temporal lobe regions gyrus of uncus, dentate gyrus, Ammon's horn, amygdala, and hippocampal gyrus, the thalamic subregions medial pulvinar, lateral ventral, anterior ventral, mediodorsal, and lateralis, and the parolfactory cortical area A25 contained low levels of VMAT2 (1% to 10% of putamen).
All of the cerebral neocortical areas and cerebellar cortex had a level of mature or intact VMAT2 that was < 1% of that of the striatum (Figure 5, inset). No statistically significant difference was observed among these brain regions, in particular between cerebellar and occipital cortex, although there were trends for the cingulate (A24 and A23) and insular cortices to have relatively higher levels of VMAT2 than other neocortical regions. The cerebral white matters, including the corpus callosum and internal capsule, had the lowest levels of VMAT2 (< 0.25% of putamen) that were barely detectable.
In contrast to the above (Figure 5), the housekeeping protein neuron specific enolase had a different brain distribution, with levels quite homogenous across the gray matter regions but, as expected, low in white matter, a finding consistent with our previous reports (Kish et al, 2005).
Given that VMAT2 is localized to dopamine, noradrenaline, and serotonin neurons, we reasoned that VMAT2 protein levels in the different brain areas (having variable dopamine–noradrenaline-serotonin ratios) might show a good correlation with the sum of the concentration of the three neurotransmitters. Using regional brain monoamine neurotransmitter data reported in our previous studies (a total of 27 brain regions; Tong et al, 2007), we found a positive correlation (Pearson) between protein levels of VMAT2 and tissue concentrations of total monoamines neurotransmitters (dopamine, noradrenaline, and serotonin;
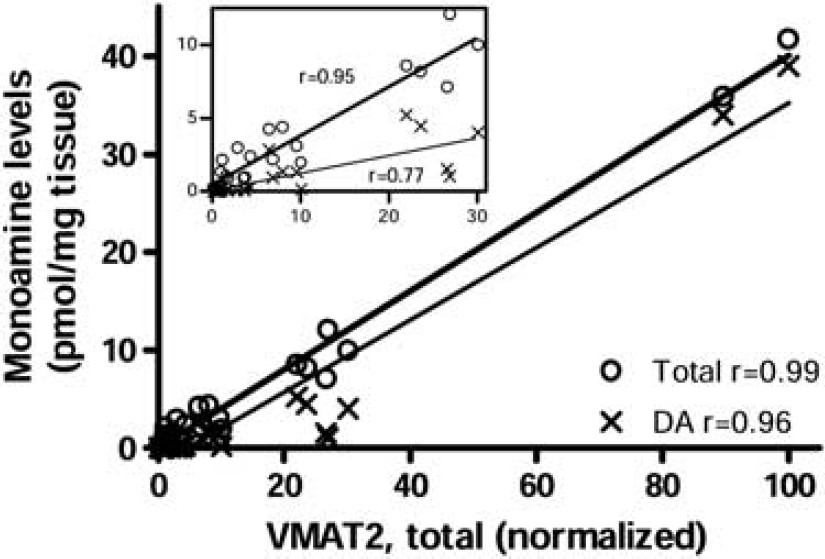
Correlation (Pearson) between levels of vesicular monoamine transporter 2 (VMAT2) protein and tissue concentrations of total monoamines (dopamine, noradrenaline, and serotonin) or dopamine (DA) alone among brain regions examined (
In our samples of normal subjects (
Discussion
To our knowledge, this is the first detailed quantitative examination of VMAT2-immunoreactive species in different human brain regions. The major finding from a practical neuroimaging perspective is that the levels of VMAT2 protein in the cerebellar cortex and the widespread cerebral neocortical areas were all at least 100-fold lower than those in striatum. This supports the usefulness of these areas as potential reference regions for PET imaging of VMAT2 in the human. We also found that the human substantia nigra was different in VMAT2 immunoreactivity from the dopamine terminal regions by containing a distinct dominant 52 k Da band IIb that was preferentially lost in parkinsonian conditions.
The multiple bands (I, IIa, IIb, III, IV) of VMAT2 detected by the commonly used C-terminus antibodies (AB1767 used in this study and another similar antibody AB1598P from Chemicon, initially developed by Erickson et al (1996) and Edwards and colleagues (Peter et al, 1995), respectively) and their uncertain identity have been a major challenge for quantitative studies of VMAT2 protein in the brain. Many studies thus chose to examine only the major band detected (Sandoval et al, 2002; Haycock et al, 2003; Little et al, 2003; Gonzalez-Hernandez et al, 2004; Fortune and Lurie, 2009), which in most cases has been the mature band I (Wang et al, 1997; Miller et al, 1999; Haycock et al, 2003; Little et al, 2003; Gonzalez-Hernandez et al, 2004; Jassen et al, 2005; Cruz-Muros et al, 2008) although in a few instances a 55-kDa band was found to be the major band in rodent brains (Holtje et al, 2003; Duchemin et al, 2009; Fortune and Lurie, 2009). The reason for the inconsistencies is not clear but could arise from differential sample preparation, particular Western blot conditions and antibody used, and/or the batch-to-batch variance of the antibodies. We have used four batches of the same version (AB1767) of the rabbit polyclonal antibodies against the C-terminus 19 amino acids of human VMAT2 from different commercial sources and have obtained the same results as observed by most of the studies, that is, the 75-kDa band I was dominant but accounted for only ∼ 50% of total VMAT2 immunoreactivity in the striatum.
In the present study, by using a newly available monoclonal antibody, by examining multiple brain regions, and by including brain of patients with degenerative parkinsonism, we established that (1) the minor smaller bands III and IV are most likely N-terminal truncated forms of VMAT2; (2) the banding pattern described above in the striatum (I, IIa, III, IV) also applies to most of the monoamine terminal fields except in the nigral dopamine cell body region (and its close proximity regions including red nucleus, subthalamic nucleus, and GPi), in which a band (IIb) of slightly smaller MW than that of band IIa could be detected; and (3) the overall extent of loss of VMAT2 immunoreactivity in the brain regions of patients with PD was correlated with the expected degree of dopaminergic neuronal damage (putamen > caudate > substantia nigra). An exception is band IIb in substantia nigra, with the extent of loss similar to that observed in the terminal regions caudate and putamen. This interesting finding, together with the limited distribution of band IIb in human brain that might be related to the distance from the dopamine cell body (e.g., GPi versus GPe), suggest that VMAT2 IIb might be the precursor to IIa and I being posttranslationally modified and transported to the terminals. However, we cannot exclude the possibility that bands I/IIa/b might represent the transporter associated with different monoaminergic neurons or different subcellular structures, for example, small synaptic vesicles versus large dense core vesicles (Nirenberg et al, 1996). The lack of an aging change of VMAT2 protein in the nigrostriatal system in our limited sample size and within a limited age span is nonetheless consistent with a previous Western blotting study (Haycock et al, 2003) and a recent PET VMAT2 imaging report (Troiano et al, 2010) although the question of the influence of aging on striatal VMAT2 in the human is still controversial (cf. Troiano et al, 2010).
Notwithstanding the generic limitation of Western blot investigations that bands with different MW could well be differentially transferred and detected, we attempted to measure the levels of total, intact, and mature VMAT2 proteins. As the tetrabenazine-binding site on VMAT2 has been mapped to the juxtaposed N- and C-terminals (Sievert and Ruoho, 1997), in particular a conserved phenylalanine (F135 of hVMAT2) in the second transmembrane domain (Gros and Schuldiner, 2010) and glycosylation does not appear to influence the transporter's activity and binding of tetrabenazine (Yelin et al, 1998), the VMAT2 proteins that are capable of binding DTBZ might include all the intact species, that is, I, IIa, and IIb, but not the N-terminally truncated III and IV. However, since the distribution patterns of total, intact, and mature VMAT2 immunoreactivity were all similar, possibly differential DTBZ-binding capacity of the different VMAT2 species should reasonably not affect the regional pattern of DTBZ binding (except, possibly, in the substantia nigra due to the unique abundance of VMAT2 IIb). Indeed, the overall brain distribution of VMAT2 proteins was generally consistent with
However, one notable difference between the results of our
In conclusion, although no cerebral gray matter region in the human brain is free of VMAT2 protein, levels of the transporter in the cerebral and cerebellar cortex were sufficiently low to satisfy this criterion as a reference region in PET VMAT2 imaging. We also found that the cerebellar cortex and the (now preferred reference region) occipital cortex were not different in VMAT2 protein concentrations. Differences in PET VMAT2 binding between the two regions might in this respect provide an example of bias caused by imaging variables such as partial volume effects in the reference region.
Footnotes
The authors declare no conflict of interest.