
Abstract
Small ubiquitin-like modifier (SUMO)2/3 but not SUMO1 conjugation is activated after transient cerebral ischemia. To investigate its function, we blocked neuronal SUMO2/3 translation through lentiviral microRNA delivery in primary cortical neurons. Viability was unaffected by SUMO2/3 silencing unless neurons were stressed by transient oxygen–glucose deprivation (OGD). Both 15 and 45 minutes of OGD were tolerated by control microRNA-expressing neurons but damaged >60% of neurons expressing SUMO2/3 microRNA. Damaging OGD (75 minutes) increased neuronal loss to 54% (control microRNA) and to 99% (SUMO2/3 microRNA). This suggests that activation of SUMO2/3 conjugation is an endogenous neuroprotective stress response.
Keywords
Introduction
Small ubiquitin-like modifier (SUMO)1-3 is a protein family which binds to lysine residues of target proteins and modulates their activity, stability, and subcellular localization (Gareau and Lima, 2010). Both SUMO2 and SUMO3 proteins are closely related, with 95% sequence identity, but both are relatively distinct from SUMO1, with ∼50% overlap of the amino-acid sequence. SUMO2 and SUMO3 are usually denoted SUMO2/3, because available antibodies cannot distinguish between these two SUMO paralogs in their mature forms. A large proportion of SUMO conjugation target proteins has proven to be transcription factors or other nuclear proteins involved in gene expression or DNA integrity (Gareau and Lima, 2010). Therefore, any marked change in levels of SUMO-conjugated proteins can be expected to have a major impact on the fate of cells. Small ubiquitin-like modifier conjugation has been shown to be activated in cultured cells exposed to various stress conditions, including anoxic conditions, hypothermia, and hypoxia (Lee et al, 2009; Yang et al, 2009). In the brain, both SUMO1 and SUMO2/3 conjugation were found to be strongly activated in hibernating animals during the torpor phase; therefore, it has been suggested that this is a protective response shielding neurons from damage induced by low blood flow and substrate deprivation (Lee et al, 2007). Small ubiquitin-like modifier-2/3 conjugation was also shown to be significantly activated during deep hypothermia (Yang et al, 2009). Hypothermia is effective in protecting the brain in various clinical conditions, including resuscitation after cardiac arrest, and is considered a promising therapy for stroke (van der Worp et al, 2010). Furthermore, a significant increase in the levels of SUMO2/3-conjugated proteins has been found in the brains of animals subjected to transient global or focal cerebral ischemia (Yang et al, 2008a, 2008b). Changes have also been reported in the pattern of SUMO2/3 protein adducts and SUMO1 protein adducts after rat transient focal cerebral ischemia and in Naval Medical Research Institute (NMRI) mice after permanent focal cerebral ischemia (Cimarosti et al, 2008). The postischemic pattern of SUMO2/3-conjugated proteins supports the assumption that elevated SUMO2/3 levels could represent a protective stress response: After transient focal cerebral ischemia, the increase in levels of SUMO2/3-conjugated proteins was most pronounced in neurons located at the border of the ischemic territory, where various survival pathways are activated, and a short, nonlethal duration of ischemia was sufficient to activate this process. Small ubiquitin-like modifier-1 conjugation increased the resistance of primary neuronal cells to oxygen–glucose deprivation (OGD), an in vitro model of ischemia (Lee et al, 2009). Oxygen–glucose deprivation-induced cell death was more pronounced in neurons in which SUMO1 expression was silenced, and cells overexpressing SUMO1 were more resistant to OGD. To date, no information is available as to whether SUMO2/3 conjugation is a protective stress response, although it is the conjugation pathway of this SUMO paralog that is particularly activated after ischemia (Yang et al, 2008a, 2008b). Owing to its potential as a target to induce brain protection, it is of considerable interest to investigate the impact of SUMO2/3 conjugation on the fate of postischemic neurons. Here we provide, for the first time, functional evidence that SUMO2/3 conjugation does indeed protect neurons from ischemia-like conditions. We silenced SUMO2/3 expression using lentivirus encoding for neuron-specific SUMO2/3 microRNA and control microRNAs, which do not interfere with any mammalian gene. We exposed primary neurons to different durations of ischemia-like conditions causing degrees of damage ranging from none to severe damage and observed the viability of neurons over time in predefined regions of interest (ROIs) with a microscopic system equipped with an automated stage.
Materials and methods
Isolation of Mouse Cortical Neurons and Neuronal Cell Culture
Primary cortical neurons were derived from mouse C57BL6/N embryos (E16) and cultured using Neurobasal medium with B27 supplement as described recently (Harms et al, 2007).
Lentivirus Production and Titration
Third-generation lentiviral particles were generated as described previously (Hauck et al, 2008; Reich et al, 2011) with the following modification: microRNA delivery was driven by the synapsin promoter based on Addgene plasmid 27232 (Dittgen et al, 2004). Target sequences and microRNA designs to interfere with SUMO2/3 and control microRNAs (LacZ and non-targeting ‘scrambled’) have been described recently (Yang and Paschen, 2009). Neuronal cultures were transduced on days in vitro (DIV) 3. Transduction efficiencies and multiplicities of infection were determined and calculated from serial dilutions in neuronal cultures using enhanced green fluorescent protein (EGFP) fluorescence as a reporter after 96 hours.
Immunocytochemistry
Cells were transduced with lentiviral particles on DIV 3 and fixed with 4% paraformaldehyde in phosphate-buffered saline as described previously (Harms et al, 2007) and incubated with primary antibodies against microtubuli-associated protein 2 (Millipore, Hofheim, Germany) and anti-EGFP (Santa Cruz Biotechnology, Heidelberg, Germany). Slides were mounted with ProLong Gold antifade reagent with 4′,6-diamidino-2-phenylindole (Invitrogen, Eggenstein, Germany).
Immunoblots
Neurons were harvested as described previously (Yang and Paschen, 2009) and probed with a SUMO2/3-specific antibody (Invitrogen; 1:1,000), anti-GAPDH (anti-glyceraldehyde 3-phosphate dehydrogenase, Millipore; 1:75,000), and anti-EGFP (Santa Cruz Biotechnology, 1:1,000).
Adenosintriphosphate Assay
Adenosintriphosphate (ATP) levels of neuronal cultures were quantified using the CellTiter-Glo Luminescent Cell Viability Assay (Promega, Mannheim, Germany) according to the manufacturer's instructions. Each data point was derived from whole-cell lysates of one 24-well and calculated as the ratio to cultures not subjected to OGD. The durations and reoxygenation time points of OGD are indicated in Figure 2A.
Oxygen–Glucose Deprivation
Neuronal cultures were subjected to combined oxygen and glucose deprivation as described previously (Harms et al, 2007). In brief, cultures were washed twice and placed in an OGD chamber ‘IN VIVO2 300’ (Ruskinn, Pencoed, UK) with 5% CO2/0.3% O2 with a buffer free of glucose for 15, 45, and 75 minutes, respectively.
Propidium Iodide Incorporation
Cell viability was assessed after staining of naive cell cultures with propidium iodide (PI) to distinguish between living and dead cells (0.001 mg/mL for 5 minutes with subsequent rinsing) and five images per well were taken using an inverted IX81 microscope (Olympus, Hamburg, Germany). Viable neurons not incorporating PI (PI−) were counted in transmission images and quantified as ratios versus all neurons as described previously (Harms et al, 2007).
Lactate Dehydrogenase Activity
Lactate dehydrogenase activity (LDH) activity as a marker for the loss of cellular membrane integrity was measured in the supernatant and after total lysis of cells using 0.5% Triton-X for 20 minutes at room temperature as described previously (Harms et al, 2001). Data are presented as raw values (Figure 2D) or as ratios between the unit of activity per mL per well and the maximum LDH activity per well for linear regression analysis (Figure 2E).
Evaluation of Cell Survival and Damage
Epifluorescence microscopic images were taken on DIV 6, 9, 12, and 13 using EGFP as a reporter for lentiviral gene delivery and microRNA expression with an inverted IX81 microscope combined with CellM software (Olympus) and equipped with a motorized microscopic stage (Merzhäuser, Wetzlar, Germany). In all, 11 ROIs were preselected per well and repeatedly analyzed over time maintaining identical settings for all experiments. Enhanced green fluorescent protein-positive cells were counted in a blinded manner and ratios calculated to compare the effects of SUMO2/3 microRNA expression on survival and OGD-induced cell loss. Each ROI initially contained ∼85±10 cells on DIV 6. In total, an average of 85 × 11 × 3 × 3=8,415 cells per condition (ROI × microRNAs × OGD durations) were analyzed in triplicate (overall ∼25,245 neurons). For visual display of neuronal survival in a particular ROI, emitted fluorescence was pseudocolored green (just before OGD) and red (24 hours after OGD) and images were merged. The resulting yellow was indicative of surviving neurons.
Statistical Analysis
Data are presented as dot blots and mean±95% confidence interval except for Figure 2A, where mean±s.d. is presented. The numbers of experiments to detect a standardized effect size ≥0.15 were calculated by a priori power analysis with the following assumptions: α=0.05, β=0.2, mean: s.d. 10% of the mean. Using GraphPad Prism (GraphPad, San Diego, CA, USA), linear regression analysis or one-way and two-way repeated-measures ANOVA (analysis of variance) were carried out with Tukey's post hoc analysis. Two-way ANOVA followed by Tukey's post hoc tests were carried out where indicated. Experiments were performed in triplicate. P-values <0.05 were considered statistically significant.
Results
Neuron-Specific Expression of Small Ubiquitin-Like Modifier-2/3 MicroRNA does not Affect Survival of Cultured Neurons
The SUMO2/3 microRNA was expressed under control of the synapsin promoter and compared with expression of control microRNAs in primary mouse cortical neurons over 12 days. Lentiviral transduction efficiencies were above 90% as analyzed by comparing transmission images with EGFP fluorescence and counting EGFP-positive cells. Neuron specificity of microRNA delivery along with EGFP as a reporter was driven by the synapsin promoter and evaluated by immunocytochemistry with the marker for mature neurons microtubuli-associated protein 2 (Figure 1A). We confirmed the reduction of SUMO2/3 protein adducts in neuronal cultures transduced with SUMO2/3 in comparison to control microRNA-coding lentiviral particles by immunoblots probed with antibodies for SUMO2/3, EGFP, and GAPDH as a loading control (Figure 1B). The SUMO2/3 protein adducts and free SUMO2/3 proteins were reduced at baseline and 3 hours (peak of SUMO2/3 protein adduction) after 45 minutes of OGD as a result of RNA interference. These blots are representative and show similar results after 15 and 75 minutes of OGD. The increase in SUMO2/3 protein adducts after OGD in control microRNA-transfected neurons correlates inversely with the decrease of free SUMO2/3 protein. There was no apparent difference in the EGFP expression pattern between groups, indicative of equal multiplicities of infection. The effects of SUMO2/3 microRNA on neuronal survival were investigated comparing the ratios between EGFP-positive cells on DIV 9/6 and DIV 12/9 (Figure 1C). There was no statistically significant difference between groups. The expression of SUMO2/3 microRNAs in primary neuronal cultures did not lead to changes in vulnerability or viability in comparison with control microRNA-transduced neurons. Furthermore, even after prolonged periods of time in culture (DIV 12/9), the survival ratios of both groups were comparable to that of the shorter life span (DIV 9/6).
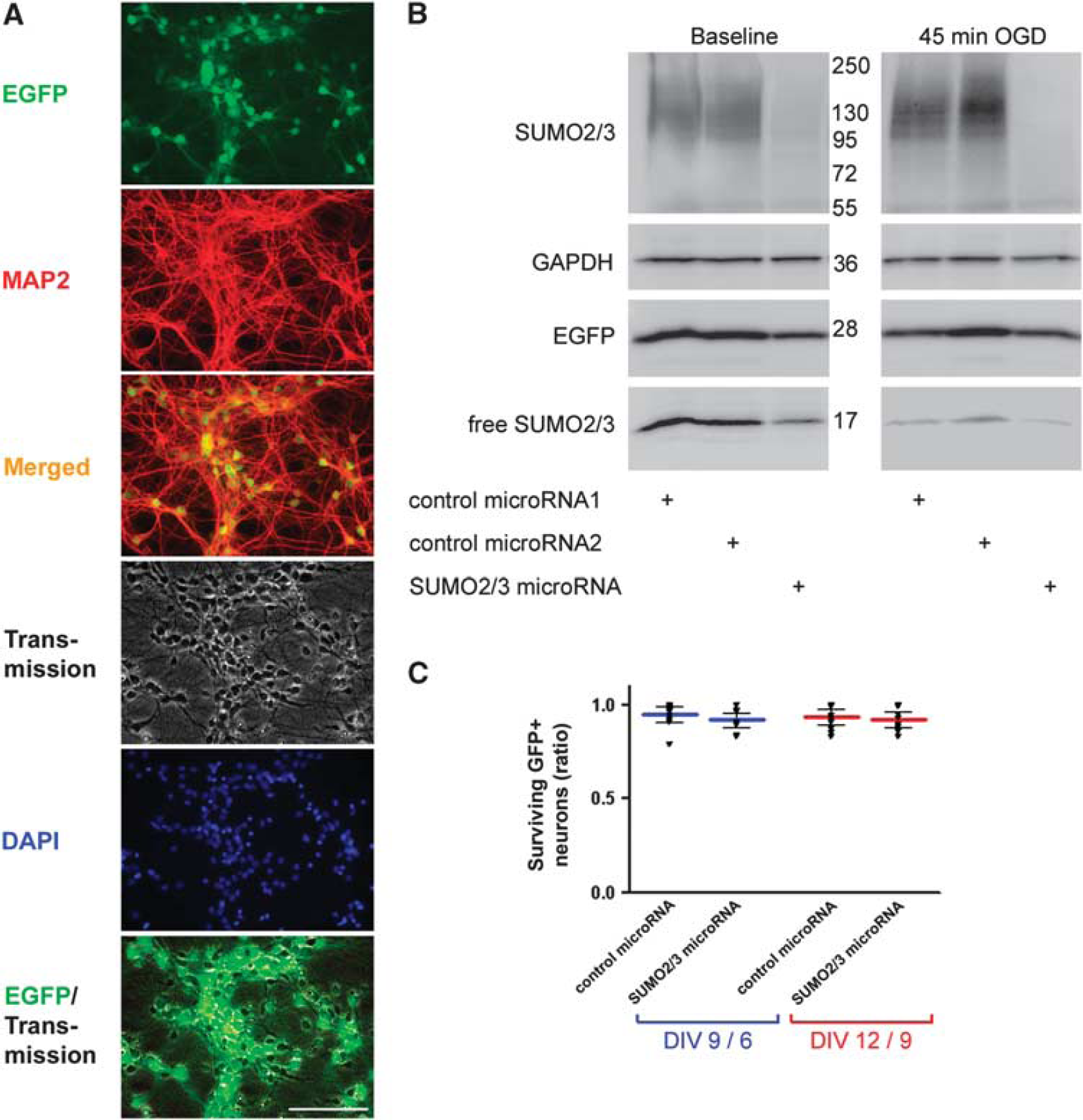
Neuronal specificity and efficacy of RNA interference with SUMO2/3 and its effects on survival of mouse primary cortical neurons. At days in vitro (DIV) 3, primary cortical neurons were transduced with lentiviral particles expressing EGFP as a reporter and either SUMO2/3 or control microRNA driven by the neuron-specific synapsin promoter. (
Tolerance to Oxygen–Glucose Deprivation is Lost in Neurons After Small Ubiquitin-Like Modifier-2/3 Silencing
Next, we investigated the influence of SUMO2/3 silencing in neuronal cultures under stress. Therefore, we chose the model of OGD that mimics ischemia-like stress in vitro. First, we quantified ATP levels in cell lysates to characterize the extent of metabolic disturbance after 15, 45, 75 minutes of OGD (Figure 2A). The ATP levels decreased to 73.2%±10.8%, 44.9%±2.0%, and 22.0%±2.1% of controls, respectively. Oxygen–glucose deprivation intervals of 15 and 45 minutes did not result in any significant cell loss of neurons expressing control microRNA (Figure 2F to 2H), which was reflected by recovery of ATP levels to 78.9%±3.3% and 73.7%±4.7% of control after 180 minutes of reoxygenation, respectively. In contrast, 75 minutes of OGD led to a significant cell loss to 54.3%±13.3% (Figure 2F and 2I) and poor recovery in ATP levels to 54.1%±17.5% after 180 minutes without lentiviral particles and to 43%±8.3% after 180 minutes in cultures expressing control microRNAs (Figure 2A, not significantly different). The maximum ATP depletion at 0-minute reoxygenation was not altered upon transduction with lentiviral particles expressing control microRNAs. Furthermore, the extent of depletion and recovery was not influenced by SUMO2/3 microRNA expression (e.g., 60.5±18.5 versus 60.4±4.1 after 0 minutes of reoxygenation and 89.5±7.5 versus 86.2±4.5 after 180 minutes of reoxygenation, control microRNA versus SUMO2/3 microRNA; 45 minutes OGD; mean±s.d.; n=4 derived from two independent experiments).
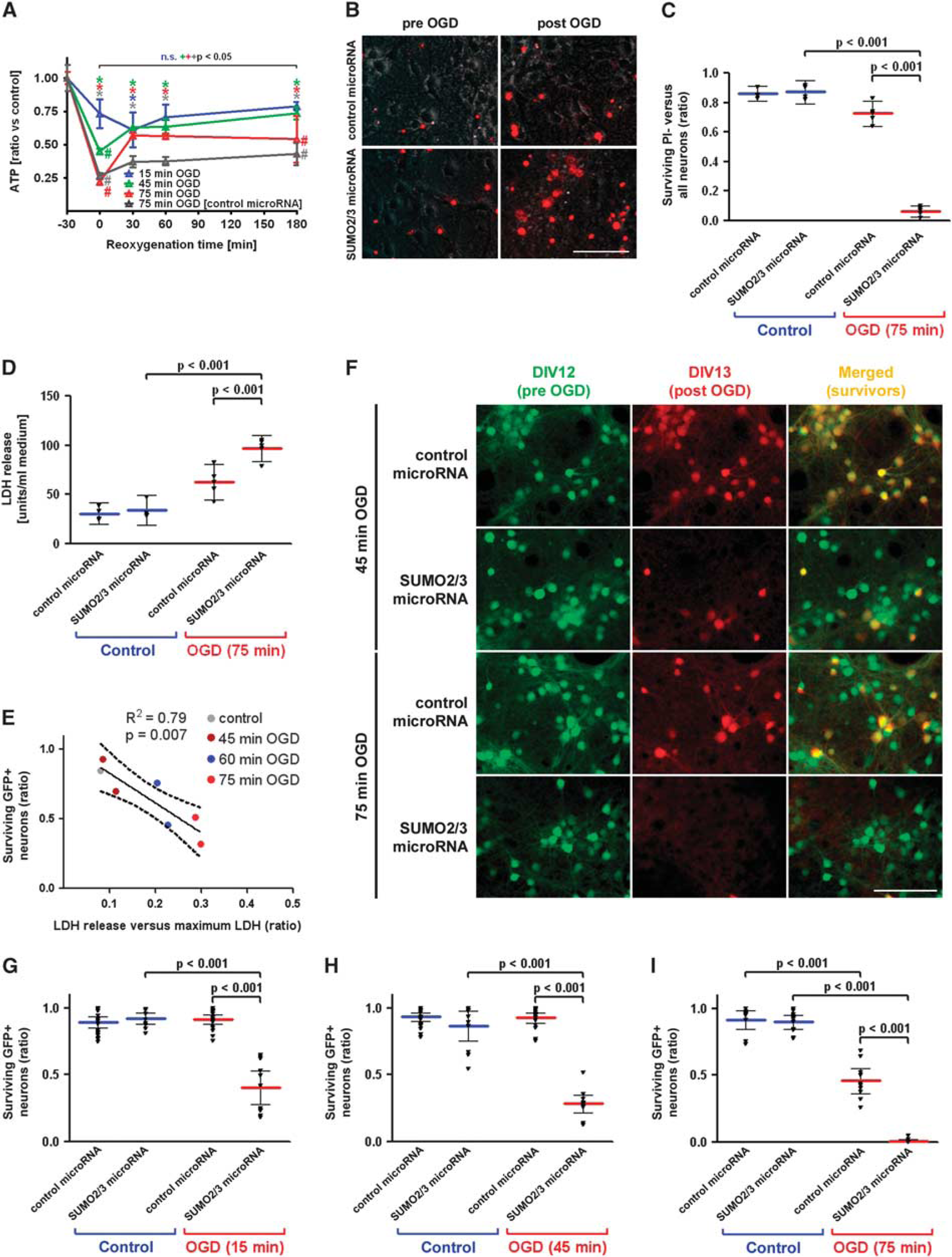
Characterization of ATP depletion and recovery after different durations of OGD and effect of SUMO2/3 microRNA expression on neuronal survival after OGD. (
An OGD lasting 75 minutes led to a significantly greater decrease of viable PI-negative neurons in the SUMO2/3 microRNA-transduced group compared with the decrease seen in neurons subjected to OGD transduced with control microRNA (Figure 2B and 2C). The loss of viable neurons depended heavily on the damage (microRNA × OGD interaction, two-way ANOVA, P<0.001). In congruence with these results, LDH activity in the supernatant, a marker for the loss of cell integrity, was significantly increased upon ischemia-like stress after 24 hours (P<0.001 for OGD × microRNA interaction, Figure 2D).
To validate our new method for determining survival rates of neurons expressing the EGFP reporter along with the microRNAs, we plotted the normalized LDH release from these cultures (shown as a ratio to the maximum LDH release) to the ratio of EGFP-positive cells. This correlation was performed at baseline and after three different lengths of OGD exposure in cultures transduced with control microRNA and analyzed 24 hours after OGD. The correlation that emerged is inverse (R2=0.79, P=0.007), signifying that the loss of GFP+ neurons in predefined ROIs before and 24 hours after OGD is accompanied by an increase in LDH activity in relation to the maximum activity achievable by total cell lysis in a given well (Figure 2E).
The RNA interference performed with neuron-specific SUMO2/3 microRNA resulted in reduction of cell numbers to 40.3%±18.8% with an OGD interval of 15 minutes, an insult which did not harm neurons expressing control microRNA (Figure 2G). With longer OGD durations, neurons expressing SUMO2/3 microRNA were even more susceptible to damage by OGD. Survival rates decreased to 28.1%±10.4% and 0.7%±1.5% for 45 and 75 minutes of OGD, respectively (Figure 2H and 2I). All comparisons between neurons transduced with SUMO2/3 or control microRNA resulted in P-values <0.001 for the interaction of microRNAs and OGD duration (Figure 2C and 2D, 2G to 2I).
Discussion
To investigate the function of SUMO2/3 induction after ischemic stress, we used primary cortical neuronal cell cultures as a model and applied RNA interference with lentivirally delivered and neuron-specific SUMO2/3 microRNA. Blocking SUMO2/3 translation and thereby target protein conjugation did not interfere with viability of neurons cultured for up to 12 DIV. Cells were then exposed to ischemia-like conditions to elucidate the role of SUMO2/3 protein conjugation in conferring resistance to experimental conditions associated with transient metabolic stress. The results provide strong evidence that neurons depend on SUMO2/3 conjugation to withstand ischemia-like stress conditions.
In this study, we used transient OGD as a simplified model to investigate the role of SUMO2/3 conjugation in ischemic stroke. As this approach does not involve cardiovascular and neurohumoral systems, we were able to focus on neuron-specific effects and conclude that metabolically compromised neurons activate a protective stress response to withstand stressful conditions. Importantly, RNA interference with SUMO2/3 did not alter the decrease and recovery of ATP per se in each degree of OGD. Understanding the molecular mechanisms underlying this process could help to establish new strategies of therapeutic intervention to increase neuronal resistance to stress.
For many years, SUMO conjugation of target proteins was believed to be a predominantly nuclear process modifying transcription factors, nuclear pore proteins, and other nuclear proteins critical for genome integrity. Recently, many SUMO conjugation target proteins have been identified in neurons that are cytosolic or cell membrane proteins, and SUMO conjugation has been found to modulate neuronal calcium fluxes and neurotransmitter release (Feligioni et al, 2009; Martin et al, 2007). Our study allows the conclusion that physiologic levels of SUMO2/3 in neurons at the onset of ischemic-like stress are neuroprotective. In addition, these data were not derived from a gain-of-function model with ectopic neuronal delivery of SUMO2/3 or ectopic delivery of the conjugation enzyme Ubc9, which might induce artificial SUMO protein adduction to target proteins, which are endogenously not conjugated. Further studies might identify and link particular SUMO2/3 target proteins to impact on the electrophysiological and metabolical/mitochondrial integrity of neurons to withstand various conditions of stress (Golebiowski et al, 2009). Former proteomics approaches and gene arrays after RNA interference with SUMO2/3 have identified a wide range of SUMOylated target proteins and transcriptional changes (Golebiowski et al, 2009; Vertegaal et al, 2004; Yang and Paschen, 2009). Therefore, it is likely that a total depletion of the free pool of SUMO2/3 does in fact interfere with the ability of neurons to react to changes in the cellular environment in general and that an ensemble of required SUMO2/3 modifications will not be performed. It would be intriguing to identify the influence of the loss of SUMO2/3 on upregulated or downregulated pathways on primary cortical neurons. Identifying this influence may in turn lead to the identification of critical target proteins in the context of vulnerability after OGD that can then be investigated experimentally. In addition, it is of pivotal importance to screen for specific drugs that activate SUMO2/3 conjugation or inhibit the deconjugation of SUMO2/3 target proteins and to modulate the fate of postischemic neurons for preventive or therapeutic purposes. Furthermore, in vivo data with stereotactical applications of lentiviral particles to achieve RNA interference with SUMO2/3 should complement and confirm the role of this endogenous stress response in the adult brain after ischemia.
Footnotes
Acknowledgements
The authors are grateful for excellent technical assistance to Anny Kretschmer, Janet Lips, and Nadine Weser.
The authors declare no conflict of interest.