
Abstract
Soluble epoxide hydrolase (sEH), a key enzyme in the metabolism of vasodilator eicosanoids called epoxyeicosatrienoic acids (EETs), is sexually dimorphic and suppressed by estrogen. We determined if the sex difference in blood flow during focal cerebral ischemia is linked to sEH. Soluble epoxide hydrolase expression in brain, hydrolase activity in cerebral vessels, and plasma 14,15-dihydroxyeicosatrienoic acid (14,15-DHET) were determined in male and female wild-type (WT) and sEH knockout (sEHKO) mice. Male, female, and ovariectomized female WT and sEHKO mice were subjected to 2-h middle cerebral artery occlusion (MCAO) and infarct size was measured at 24 h of reperfusion. Laser—Doppler cortical perfusion during MCAO was compared among groups and differences in cortical blood flow rates were confirmed using in vivo quantitative optical microangiography. Cerebrovascular expression and activity of sEH and plasma 14,15-DHET were lower in WT female than male mice, and blood flow during MCAO was higher and infarct size was smaller in WT female compared with male mice. Sex differences in cerebral blood flow and ischemic damage were abolished after ovariectomy and were absent in sEHKO mice. We conclude that sEH is an important mechanism underlying sex-linked differences in blood flow and brain damage after cerebral ischemia.
Introduction
Stroke incidence and mortality rates are lower in premenopausal women relative to men of the same age, but the incidence of cerebrovascular events rapidly increases in women after menopause, and because women live longer, more women are affected by stroke than men (Reeves et al, 2008). Changes in stroke risk and sensitivity in women after menopause have been linked to estrogen. Estrogen promotes vasodilation and exhibits vasoprotective properties (Miller and Duckies, 2008). We have previously shown that female rats maintain higher cerebral blood flow (CBF) and sustain smaller infarct size after middle cerebral artery occlusion (MCAO) compared with male rats, and the difference in blood flow and relative protection from stroke in female rats was lost after ovariectomy (Alkayed et al, 1998). The mechanism of protection by estrogen and the underlying mechanism for the differential blood flow response to cerebral ischemia remain unknown.
Soluble epoxide hydrolase (sEH) is an important enzyme involved in the metabolism and terminal inactivation of lipid signaling molecules referred to as epoxyeicosatrienoic acids (EETs; Liu et al, 2004). We have recently shown that sEH knockout (sEHKO) male mice were protected from ischemic brain injury, and that protection was associated with higher blood flow during MCAO compared with wild-type (WT) male mice (Zhang et al, 2008). In peripheral tissue, sEH expression is sexually dimorphic and regulated by sex hormones (Pinot et al, 1995; Sinal et al, 2000). We have also shown that sEH expression in brain is downregulated by estrogen (Koerner et al, 2008). In the current study, we tested the hypothesis that lower sEH expression in cerebral vasculature in female versus male mice contributes to higher blood flow during MCAO and smaller infarct size in female compared with male mice. To test this hypothesis, we subjected male and female WT and sEHKO mice to a transient MCAO, and monitored blood flow during MCAO using optical microangiography (OMAG), a novel, noninvasive optical imaging method for in vivo, online monitoring and quantification of blood perfusion in brain through an intact skull.
Materials and methods
This study was conducted in accordance with the National Institutes of Health guidelines for the care and use of animals in research, and protocols were approved by the Institutional Animal Care and Use Committee at Oregon Health and Science University, Portland, OR, USA.
Animals
Homozygous sEHKO and WT littermates were generated in-house by breeding heterozygous sEHKO mice. Genotype was confirmed by PCR, as previously described (Zhang et al, 2008). Briefly, 250 ng of genomic DNA was analyzed by PCR using the following primers: F1, 5'-TGGCAC-GACCCTAATCTTAGGTTC-3'; R2, 5'-CACGCTGGCATTT TAACACCAG-3'; and R3, 5'-CCAATGACAAGACGCTG GGCG-3'. Wild-type DNA yielded a 233-bp band, homozygous (sEHKO−/-), a 190-bp band and heterozygous (sEHKO+/-) DNA double bands at 233 and 190 bp.
Middle Cerebral Artery Occlusion in Mice
Transient focal cerebral ischemia was induced in mice (25 to 28 g body weight) using the intraluminal MCAO technique (Zhang et al, 2007). Mice were anesthetized with isoflurane (1.5% to 2%) and kept warm with water pads. A laser—Doppler probe was affixed to skull to verify vascular occlusion and reperfusion. A silicone-coated 6–0 nylon monofilament was advanced into the right internal carotid artery until laser—Doppler signal decreased to less than 30% of baseline. After securing the filament in place, the surgical site was closed. The animal was awakened and assessed at 2 h of occlusion for neurologic deficit. Mice with clear neurologic deficit were reanesthetized, laser—Doppler probe repositioned over the same site on the skull, and the occluding filament was withdrawn to allow reperfusion. Brains were harvested at 24 h, sliced into five 2-mm-thick coronal sections, and stained with 2,3,5-triphenyltetrazolium chloride. Sections were incubated in 1.2% 2,3,5-triphenyltetrazolium chloride in saline for 15 mins at 37°C and then fixed in formalin for 24 h. Sections were photographed, and infarcted (unstained) areas were measured with MCID (InterFocus Imaging Ltd, Linton, Cambridge, UK) imaging software. To account for the effect of edema, we estimated infarct size indirectly by subtracting the uninfarcted area in the ipsilateral hemisphere from the total area of the contralateral hemisphere.
Western Blotting
Brain tissue was homogenized in solution A containing sucrose (250 mmol/L), KCl (60 mmol/L), Tris-HCl (15 mmol/L), NaCl (15 mmol/L), EDTA (5 mmol/L), EGTA (1 mmol/L), PMSF (0.5 mmol/L), and DTT (10 mmol/L), then centrifuged at 2,000g for 10 mins at 4°C. Protein samples (15 μg) were separated by SDS—polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes. Blots were then blocked in 5% dry milk and incubated at 4°C overnight with a primary rabbit anti-sEH antibody (Zhang et al, 2007). The signal was visualized using a biotinylated secondary antibody with an ECL Plus chemiluminescence detection kit. Autoradiograms were scanned and band optical densities were quantified and expressed relative to β-actin.
Isolation of Cerebral Vessels
Mice were deeply anesthetized with isoflurane and perfused transcardially with prechilled heparinized saline (1 U/mL) before decapitation; and brains were rapidly dissected. Pial vessels and large cerebral vessels were dissected and placed on ice in homogenization buffer (20 mmol/L Tris-HCl (pH 7.4); 0.32 mol/L sucrose; 1 mmol/L EDTA) containing protease inhibitor cocktail. Six mice per group were dissected, and vessels from three animals were combined to obtain sufficient material for vascular hydrolase activity.
14,15-DHET ELISA
Total plasma 14,15-dihydroxyeicosatrienoic acid (14,15-DHET) was measured using a commercial ELISA kit (Detroit R&D Inc., Detroit, MI, USA). Plasma lipids were extracted three times with acidified ethylacetate (pH 3 to 4), followed by saponification with 20% KOH. Samples were then dried and resuspended in methanol.
Hydrolase Activity Assay
Hydrolase activity in isolated cerebral vessels was measured by incubating vessels with 14,15-EET and assaying for 14,15-DHET as described above. Briefly, homogenization buffer (20 mmol/L Tris-HCl (pH 7.4); 0.32 mol/L sucrose; 1 mmol/L EDTA) was added to isolated vessels (100 μL for each 5 mg). Samples were sonicated on ice three times for 5 to 10 secs and then centrifuged for 10 mins at 1,000g. Protein concentration was determined in supernatants, hydrolase enzymatic reactions were initiated by adding 1 μmol/L 14,15-EET and incubated in a 37°C shaking water bath for 1 h.
Optical Microangiography
Blood perfusion is visualized and quantified based on endogenous light scattering from moving blood cells within biologic tissue, with no need to use exogenous contrasting agents. In the mouse, optical perfusion imaging can be achieved through an intact skull, and reaches regions as deep as 1.5 mm beneath skull, covering the entire cerebral cortex in mouse. We used a superluminescent diode with a central wavelength of 1,310 nm and a full-width, half-maximum bandwidth of 50 nm to illuminate the OMAG system (Wang et al, 2007; Wang and Hurst, 2007; Wang, 2007). The spectral interferograms formed by the reference light and the light backscattered from the tissue sample were detected by a custom-built, high-resolution and high-speed spectrometer. A final volume data cube of 1,000 × 500 × 512 (x—y—z) voxels was built from which the three-dimensional OMAG structural and flow images were computed. The three-dimensional scan represented a physical volume with the following x—y—z dimensions: 2.5 × 2.5 × 2.0 mm3.
Iodoantipyrine Autoradiography
Mice were instrumented with femoral artery and jugular vein catheters, and 1 μCi of [14C] iodoantipyrine (IAP) in 75 μL saline was infused intravenously for 45 secs. Free-flowing arterial blood was simultaneously sampled at 5-sec intervals for the arterial input function. Mice were decapitated at 45 secs after the start of infusion, and brains were quickly removed and frozen in 2-methylbutane on dry ice. Brains were sectioned on a cryostat into 20-μm-thick coronal slices, which were thaw-mounted on cover glass and then apposed for 3 weeks to film together with 14C standards. The concentration of [14C] IAP in blood was determined by liquid scintillation spectroscopy. Autoradiographic images representing three different coronal levels (0, −1, and −2 from bregma, five images each) were digitized, and rCBF in the same 12 locations, represented in Figure 5A, were determined with the use of image analysis software (MCID, 7.0).
Statistical Analysis
Data were analyzed with a t-test for two groups and analysis of variance with post hoc Newman—Keuls test for multiple groups. The criterion for statistical significance was set at P < 0.05. All values are reported as mean ± s.e.m.
Results
The level of sEH in brain was higher in WT male compared with WT female mice (Figure 1A and 1B) and was undetectable in sEHKO male and female mouse brain (not shown). Cerebrovascular hydrolase activity, as measured by conversion of 14,15-EET to 14,15-DHET, was higher in WT male compared with female mice, and was low in vessels from male and female sEHKO mice (Figure 1C). Plasma levels of 14,15-DHET in WT females (364 ± 77 pg/mL, n = 8) were significantly lower than the values we previously reported in males (4,179 ± 482 pg/mL) (Zhang et al, 2008), and were not different than plasma 14,15-DHET in sEHKO females (236 ± 44 pg/mL). Wild-type female mice sustained smaller infarcts after MCAO compared with males. Male and female sEHKO mice had similar infarcts, which were significantly smaller than WT male mice, but not different than WT females (Figure 2). The sex difference in infarct size in WT mice disappeared when females were ovariectomized (OVX). However, ovariectomy did not increase infarct size in sEHKO mice (Figure 2). Similarly, laser—Doppler perfusion (LDP) at the end of MCAO was significantly higher in WT female compared with WT male mice, and the difference disappeared after ovariectomy (Figure 3). Laser—Doppler perfusion was significantly higher in male and OVX sEHKO compared with corresponding male and OVX WT mice. Laser—Doppler perfusion was not different between WT and sEHKO female mice. We confirmed LDP findings using quantitative OMAG (Wang et al, 2007; Wang and Hurst, 2007). The images in Figure 4A illustrate the reduction in blood flow in the ipsilateral (I) compared with the contralateral (C) cerebral cortex during MCAO in male and female WT and sEHKO mice. Ischemic images were obtained at 30 mins after filament insertion, and compared with preocclusion baseline. Because baseline blood flow was not different between WT and sEHKO mice, in this figure we used baseline images only from sEHKO mice. Cortical perfusion rates in these images were quantified and compared among groups. Figure 4B confirms laser—Doppler data in Figure 3 and shows in deeply situated cortical regions that blood flow rates in the ischemic cortex were significantly higher in WT female compared with male mice. The graph also shows that male sEHKO mice maintained significantly higher perfusion rates during MCAO than WT males, but there were no differences in brain tissue perfusion during MCAO between WT and sEHKO females. Finally, the gender difference in blood flow observed in WT mice was absent in sEHKO mice. In fact, blood flow rate in male sEHKO mice was numerically higher than female sEHKO mice, although the difference was not statistically significant. Figure 5A depicts the locations of regions selected for correlative studies using OMAG in vivo and IAP autoradiography post mortem. Figure 5B is a scatter plot relating blood flow rates obtained by OMAG in vivo against post-mortem measurements using IAP autoradiography. Figure 5C shows plot of the averaged rCBF over the hemispheres for each mouse obtained from OMAG in vivo and IAP post mortem. The correlation between OMAG and IAP was ~0.96. Thus, OMAG agreed with IAP with acceptable errors.
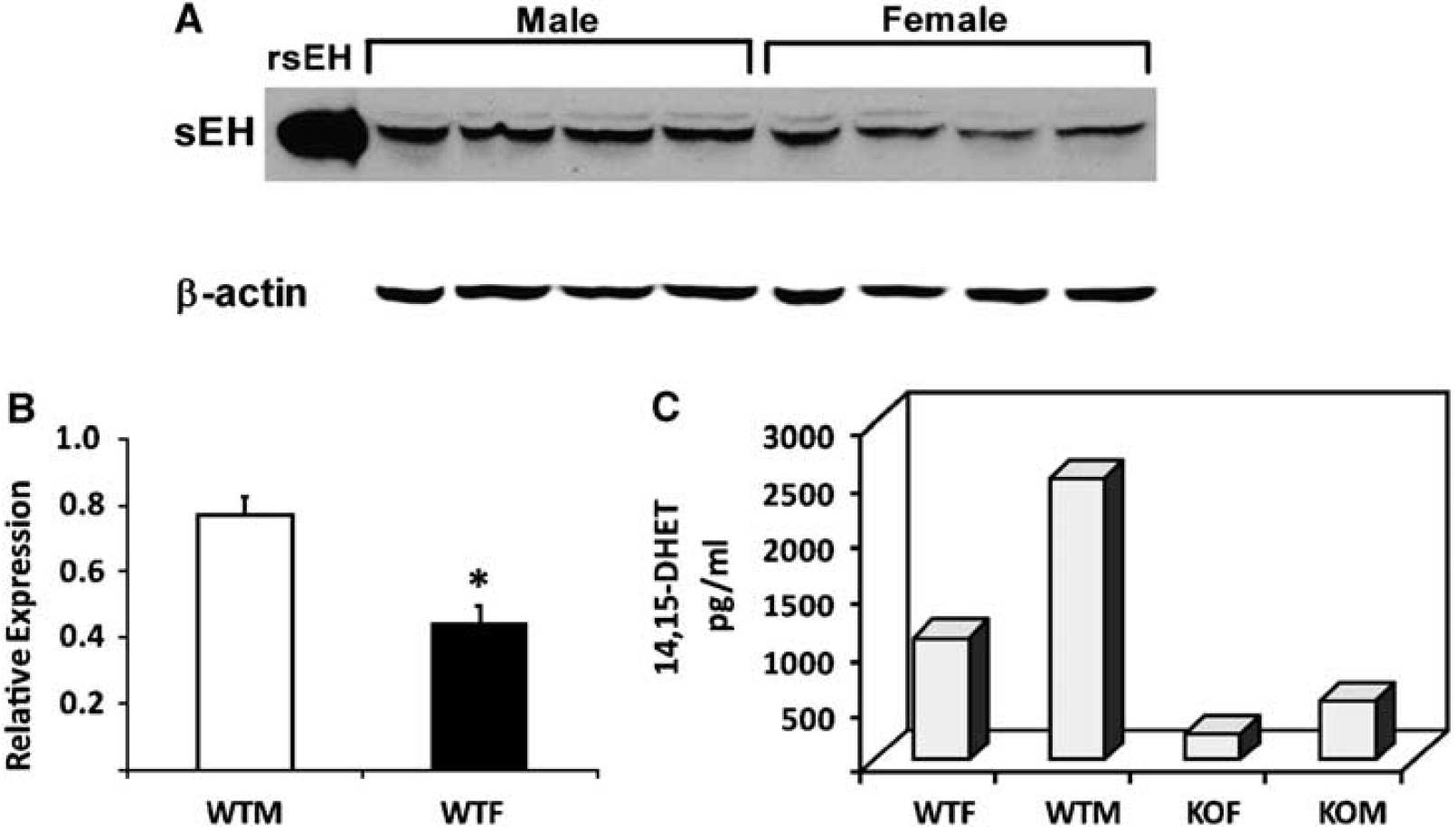
Sex differences in soluble epoxide hydrolase (sEH) protein expression in brain. (
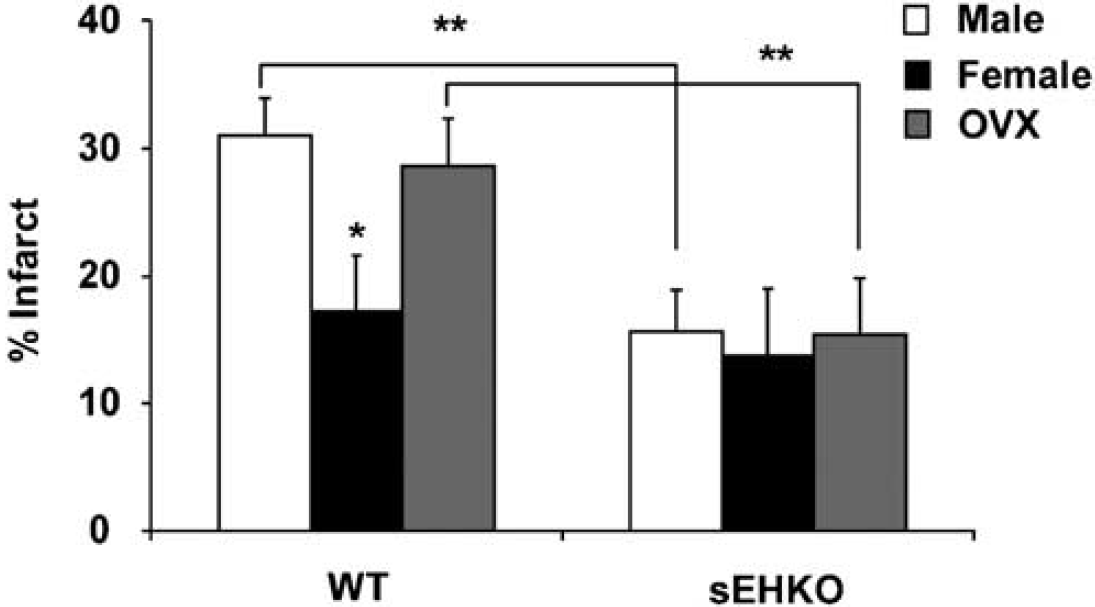
Role of sEH in the sex difference in infarct size after MCAO in mice. Female WT mice sustained smaller infarcts after MCAO compared with male WT mice (17.1% ± 4.2%, versus 31.1% ± 2.9%, n = 10, P < 0.05). Infarct size was smaller in sEHKO compared with WT male mice (15.6% ± 3.3% versus 31.1% ± 2.9%, n =10 each, P < 0.01), but not in sEHKO versus WT female mice (13.7% ± 5.0% compared with 17.1% ± 4.2%, n = 10 each, P > 0.05). When female mice were ovariectomized (OVX), infarct size was increased in WT (from 17.1% ± 4.2% to 28.6% ± 2.9%, P < 0.05), but not in sEHKO mice (13.7% ± 5.0% versus 15.5% ± 4.4%, n = 10 each, P > 0.05). *Different than WT males and OVX females. **Significant difference between corresponding WT and sEHKO groups.
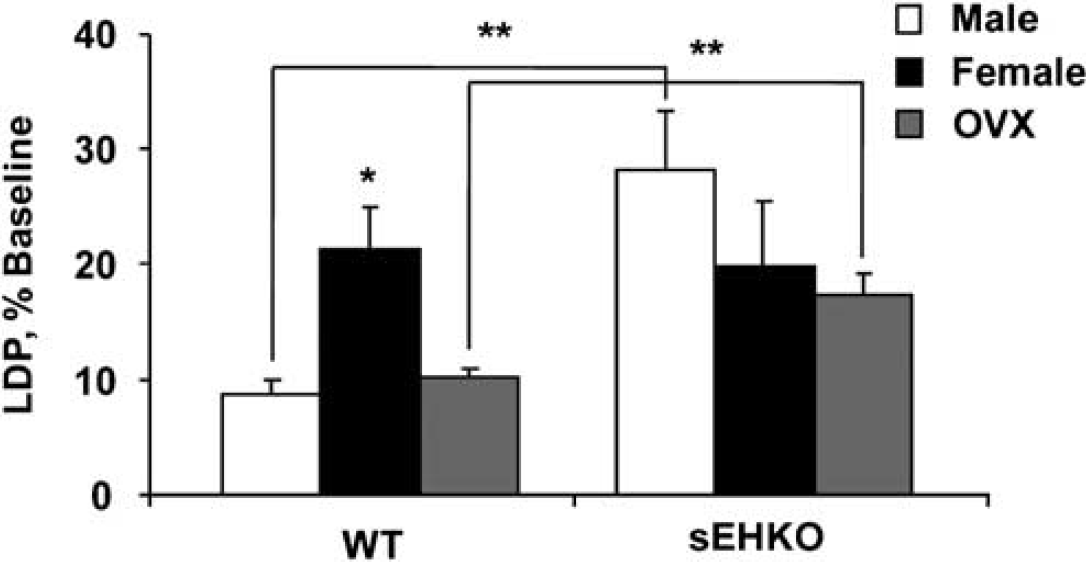
Role of sEH in the sex difference in laser—Doppler perfusion (LDP) during MCAO. Relative changes from baseline in LDP were higher in WT females compared with males and ovariectomized females (21.4% ± 3.6% versus 8.8% ± 1.3% and 10.1% ± 0.9%, n = 10 each, P < 0.05). LDP was significantly higher in male and ovariectomized sEHKO mice compared with male and ovariectomized WT mice (28.3% ± 5.0% and 17.3% ± 2.0% versus 8.8% ± 1.3% and 10.1% ± 0.9%, n =10 each, P < 0.05). *Different than WT males and OVX females. **Significant difference between corresponding WT and sEHKO groups.
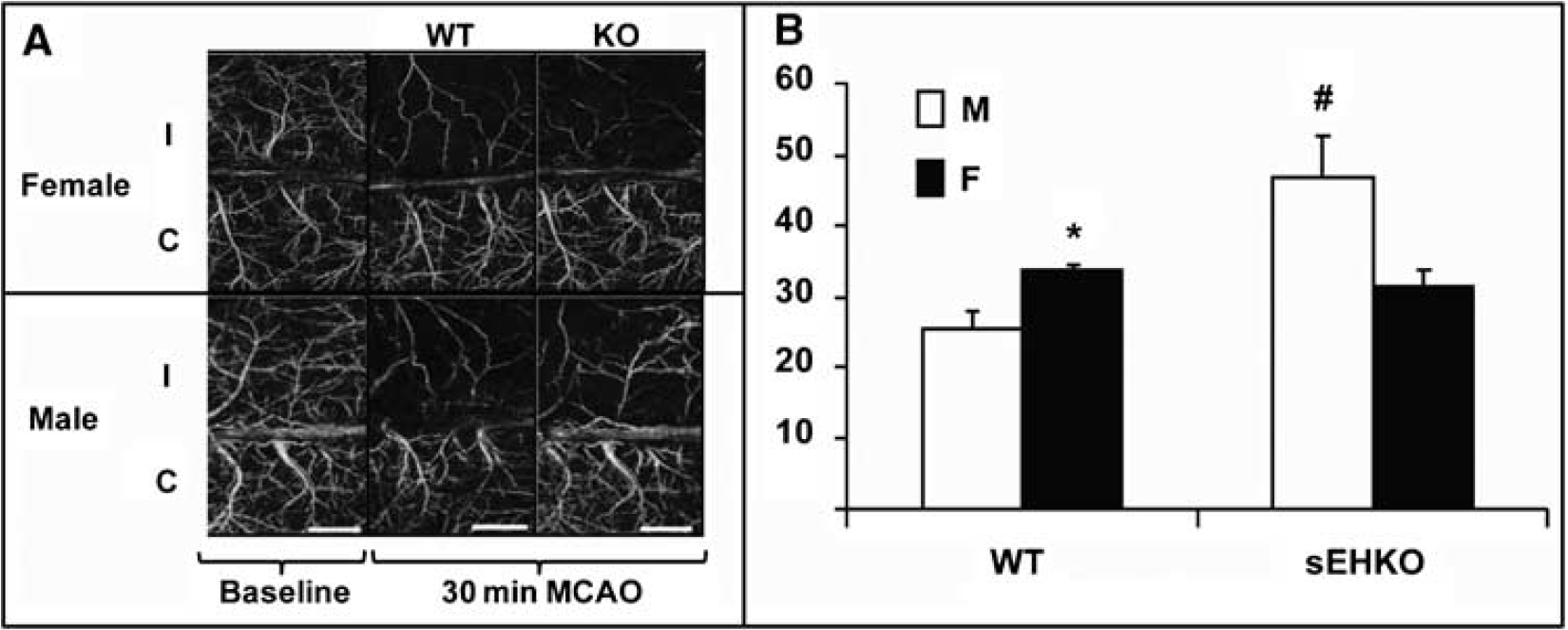
Cerebral blood flow imaging using optical microangiography (OMAG) in ipsilateral (I) and contralateral (C) cerebral cortex of male and female WT and sEHKO mice at baseline and 30 mins after MCAO. (
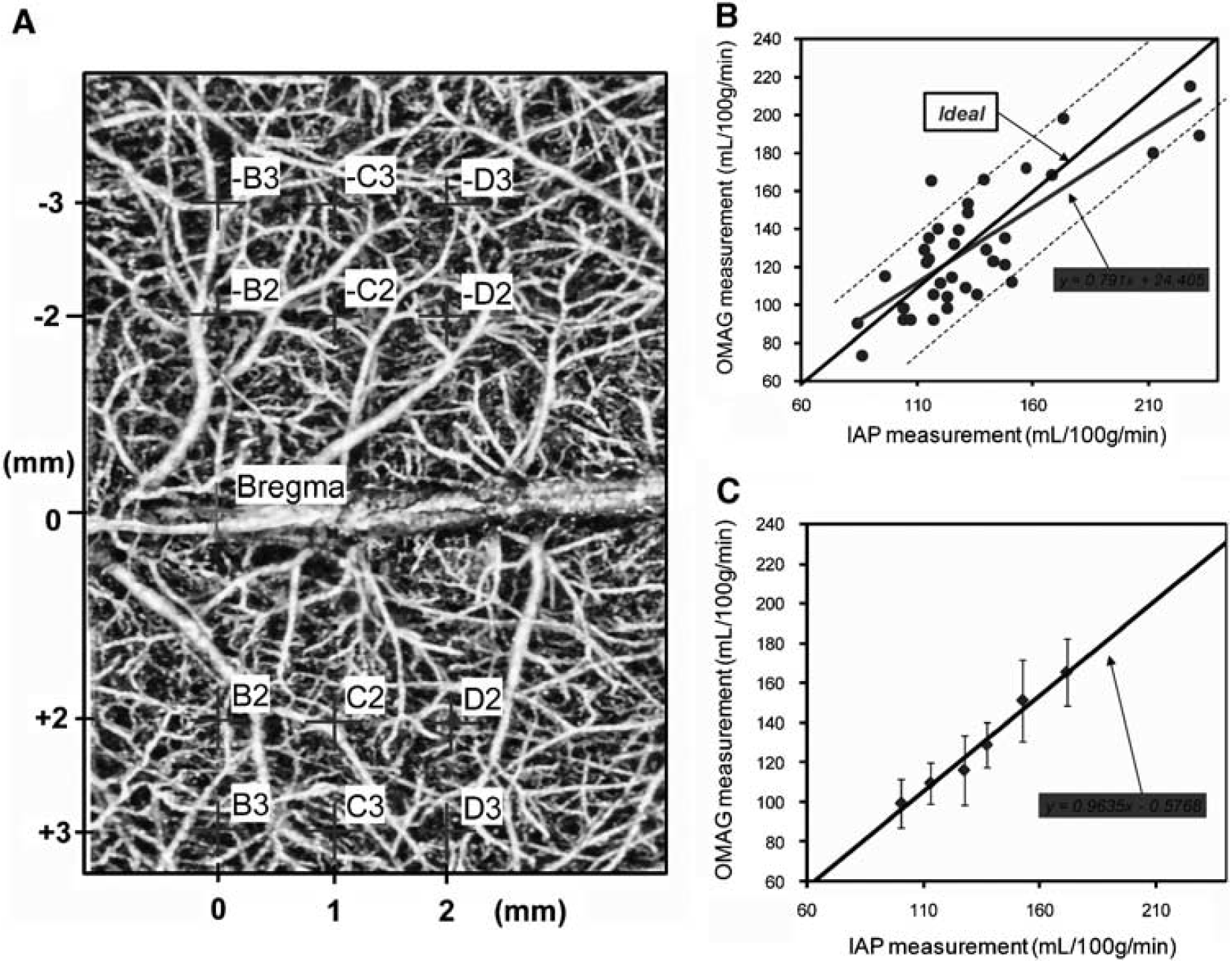
Correlation of in vivo measurements of cortical perfusion using optical microangiography (OMAG) and blood flow rates using [14C] iodoantipyrine (IAP) autoradiography. (
Discussion
We found that female mice have lower cerebrovascular expression and activity of sEH, and higher blood flow and smaller infarct after MCAO compared with male mice. Deletion of sEH increases blood flow and decreases infarct size in male, but not female sEHKO mice, eliminating the differences between the two sexes. We further found that ovariectomy decreases blood flow and increases infarct size in WT, but not sEHKO female mice. The findings suggest that the protection enjoyed by females against ischemic injury is in part because of their higher ability to increase blood flow in response to focal vascular occlusion, and that higher vascular capacity in females is linked to lower cerebrovascular sEH. The findings are in agreement our previous observations that sEH is downregulated in brain by estrogen (Koerner et al, 2008), and that sEH is an important regulator of blood flow during MCAO (Zhang et al, 2008).
Soluble epoxide hydrolase is abundantly expressed in brain, both in neurons (Zhang et al, 2007) and blood vessels (Zhang et al, 2008). The role of sEH in neurons remains unclear. However, vascular expression of sEH is believed to be important in terminating the biologic activity of EETs, which are known vasodilators in the cerebral circulation (Liu et al, 2004). In agreement with this idea, lower expression and activity in females correlated with lower plasma levels of 14,15-DHET, the metabolic product of 14,15-EET formed by sEH, and with higher blood flow during MCAO. In this model of focal cerebral ischemia, higher residual perfusion during MCAO may be explained by more extensive collateral blood flow, due either to higher vasodilator capacity or higher small vessel and capillary density in females compared with male mice. EETs have been shown to be released during hypoxia ischemia (Nithipatikom et al, 2001; Kiss et al, 2008), which can promote vasodilation and angiogenesis (Munzenmaier and Harder, 2000; Webler et al, 2008; Suzuki et al, 2008). By suppressing cerebrovascular sEH, estrogen may increase bioavailable EETs, enhancing vasodilation and new vessel formation.
Vasodilation by EETs is likely mediated by activation of large-conductance Ca2+-activated K+ channels in vascular smooth muscle membrane, leading to hyperpolarization and vascular smooth muscle relaxation (Spector and Norris, 2007). The angiogenic and proliferative effects of EETs are attributed to actions on such signaling pathways as mitogen-activated protein kinase, protein kinase A, and other tyrosine kinases and phosphatases (Fleming, 2007).
Our study results suggest that sEH expression in females is inhibited by ovarian steroids, likely estrogen (Koerner et al, 2008), and that loss of hormones removes this inhibition, increasing sEH and contributing to the exacerbation in ischemic damage observed after ovariectomy. In support of this notion, removal of sex hormones by ovariectomy did not increase infarct size in OVX female sEHKO mice, suggesting that sEH is important in the increase in ischemic damage observed after ovariectomy.
Because laser—Doppler flowmetry reflects brain surface perfusion and depicts the relative change in perfusion rather than absolute CBF, we used a newly developed quantitative optical imaging technique to quantify CBF within MCA-dependent territory during occlusion and to confirm the differences in CBF observed with the laser—Doppler. We validated blood flow rates obtained with this method using the standard method of blood flow measurement using IAP autoradiography. OMAG values obtained in vivo correlated well with post-mortem values obtained with autoradiography.
In summary, we have shown that female mice sustain more favorable outcome after ischemic brain injury, in part because of flow-preserving effects of reduced sEH expression compared with males. Our finding provides insight into one of the neuroprotective mechanisms of estrogen in stroke. Understanding the function of sEH in ischemic brain injury will contribute to our understanding of stroke-related brain damage and may afford new approaches for stroke injury treatment and prevention in both sexes.
Footnotes
Acknowledgements
We acknowledge the Department of Anesthesiology Animal Models Core (Dr Stephanie Murphy, Rui Yang, Nicole Libal and Sarah Mader), the OHSU Bioanalytical Core (Dr Dennis Koop) and editorial assistance by Kathy Gage. Studies were supported by R01 NS044313 (NJA), R01 HL093140 (RKW) and P01 NS049210 (NJA and PDH).
The authors declare no conflict of interest.