
Abstract
Activation of endogenous stem cells has been proposed as a novel form of therapy in a variety of neurologic disorders including traumatic brain injury (TBI). Vascular endothelial growth factor (VEGF) is expressed in the brain after TBI and serves as a potent activator of angiogenesis and neurogenesis. In this study, we infused exogenous VEGF into the lateral ventricles of mice for 7 days after TBI using mini-osmotic pumps to evaluate the effects on recovery and functional outcome. The results of our study show that VEGF significantly increases the number of proliferating cells in the subventricular zone and in the perilesion cortex. Fate analysis showed that most newborn cells differentiated into astrocytes and oligodendroglia and only a few cells differentiated into neurons. Functional outcome was significantly better in mice treated with VEGF compared with vehicle-treated animals after TBI. Injury size was significantly smaller at 90 days after TBI in VEGF-treated animals, suggesting additional neuroprotective effects of VEGF. In conclusion, VEGF significantly augments neurogenesis and angiogenesis and reduces lesion volumes after TBI. These changes are associated with significant improvement in recovery rates and functional outcome.
Keywords
Introduction
At least 1.4 million Americans have traumatic brain injury (TBI) each year according to the Center for Disease Control (Langlois et al, 2004). Of these, 50,000 die, 235,000 are admitted to hospital, and 90,000 are left with permanent disability (Langlois et al, 2004). Neuroprotective strategies aimed at preventing cellular death in the affected brain have been largely disappointing, and to date, no neuroprotective agent is clinically available (Beauchamp et al, 2008; Leker and Shohami, 2002).
Repair mechanisms based on the proliferation of endogenous cells have been identified in the post-traumatic brain, suggesting that precursors at the ventricular surface proliferate, migrate to sites of injury, and differentiate into neurons (Chirumamilla et al, 2002; Dash et al, 2001). However, spontaneous neurogenesis is not sufficient to induce meaningful recovery (Shetty et al, 2004). Therefore, augmenting neurogenesis is a reasonable approach to improve functional outcome after TBI. Owing to the many similarities in response to injury that exist between cerebral trauma and ischemia (Leker and Shohami, 2002) important insights into the development of clinically significant therapies can be learned from our experience in stroke. Preliminary results in a model of cerebral ischemia show that neurogenesis can be significantly enhanced with the continuous delivery of basic fibroblast growth factor (FGF2) into the injured cortex (Leker et al, 2007). However, merely augmenting the number of newborn cells in the brain may not suffice as these cells depend on newly formed blood vessels for their metabolic and trophic needs (Gage et al, 1998; Horner and Palmer, 2003; Palmer et al, 2000).
Vascular endothelial growth factor (VEGF) is a trophic factor that is expressed in the CNS after injury (Dore-Duffy et al, 2007; Hayashi et al, 1997; Skold et al, 2005; Wang et al, 2005) and induces angiogenesis (Nag et al, 1997; Nieto et al, 2001). VEGF may also have beneficial effects on the survival of newborn neuronal precursors (Widenfalk et al, 2003) and has been implicated in neurogenesis (Jin et al, 2002) after cerebral ischemia (Sun et al, 2003; Wang et al, 2007) and neurite outgrowth (Jin et al, 2006). Furthermore, VEGF has important implications in increasing the size of the subventricular zone (SVZ) (Gotts and Chesselet, 2005), and inhibition of VEGF expression after injury may actually exacerbate outcome (Skold et al, 2006). However, VEGF could be a double-edged sword as it may increase vascular permeability and exacerbate cerebral edema, thus causing unwanted effects (Hayashi et al, 1997). Furthermore, as previously shown, VEGF antagonists can reduce brain edema formation after brain insult (van Bruggen et al, 1999). Therefore, this study was designed to explore the effect of continuous infusion of VEGF into the lateral ventricles of mice on neurogenesis, differentiation, and functional recovery of mice after TBI.
Materials and methods
Animals and Traumatic Brain Injury Model
The study was conducted according to the Institutional Animal Care and Use Committee guidelines in compliance with National Institutes of Health guidelines. Adult Sabra male mice weighing 40 g (
BrdU Injections
From day 4 after injury or sham operation, and during 6 days, all animals received i.p. injection of the tracer 5-bromo-2-deoxyuridine (BrdU; 50 mg/kg, q12 h) to label dividing cells.
Evaluations
Neurobehavioral evaluation:
Animals were weighed and examined with a standardized neurologic severity score (NSS) during 3 months after injury. During the first week, testing was performed every other day, then until the end of the second month every week, and finally, at 90 days after TBI. Ten different tasks were used to evaluate motor ability, balancing, and alertness of the tested animal and points were given for failure to perform a task (Beni-Adani et al, 2001). The pathologic scores correlate very well with clinical disability scores and with the degree of brain edema (Shapira et al, 1988). The first NSS was obtained at 1 h after TBI, and it reflects the initial severity of injury. Neurologic severity score at 1 h is predictive of both mortality and morbidity and it also correlates well with the extent of damage seen on MRI (Tsenter et al, 2008). Therefore, we selected only animals with an NSS = 6 at 1 h for this test. The extent of recovery is calculated as the difference between the NSS at 1 h and at any subsequent time point (ΔNSS).
Injury size:
At 90 days after the surgery, the animals were deeply anesthetized and perfusion fixed with 4% paraformaldehyde. Their brains were frozen-sectioned at 10 μm. Brain slices 200 μm apart between bregma + 1.42 and bregma −0.8 were stained with Giemsa stain-modified solution (1:1; Fluka, Sigma-Aldrich Corporation, St Louis, MO, USA) and digitally photographed. The volume of injured tissue was measured with ImageJ.40 g software (National Institutes of Health, Bethesda, MD, USA). Damaged tissue volume was calculated by dividing the volume of the injured hemisphere by that of the nonlesioned hemisphere (as shown in Figure 2C; Swanson et al, 1990) The results are expressed as a percentage of hemispheric tissue.
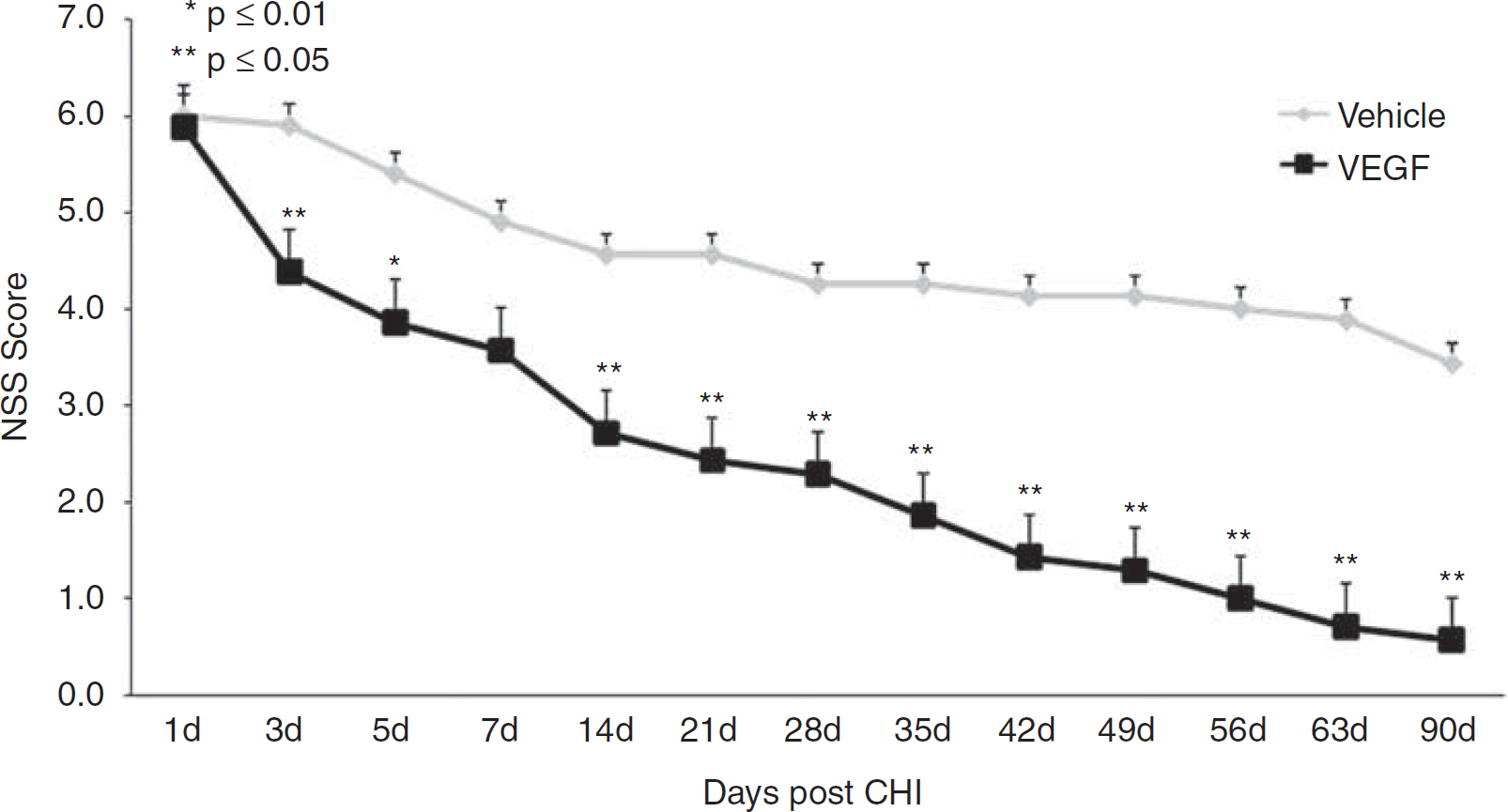
Vascular endothelial growth factor (VEGF) improves neurologic disability. Animals underwent closed head injury (CHI) and were treated with VEGF or vehicle. Neurologic disability was evaluated at different time points after CHI with the neurologic severity score (NSS). Mice treated with VEGF showed significantly larger improvement beginning at day 3 after CHI and the curves continued to diverge until the end of the study.
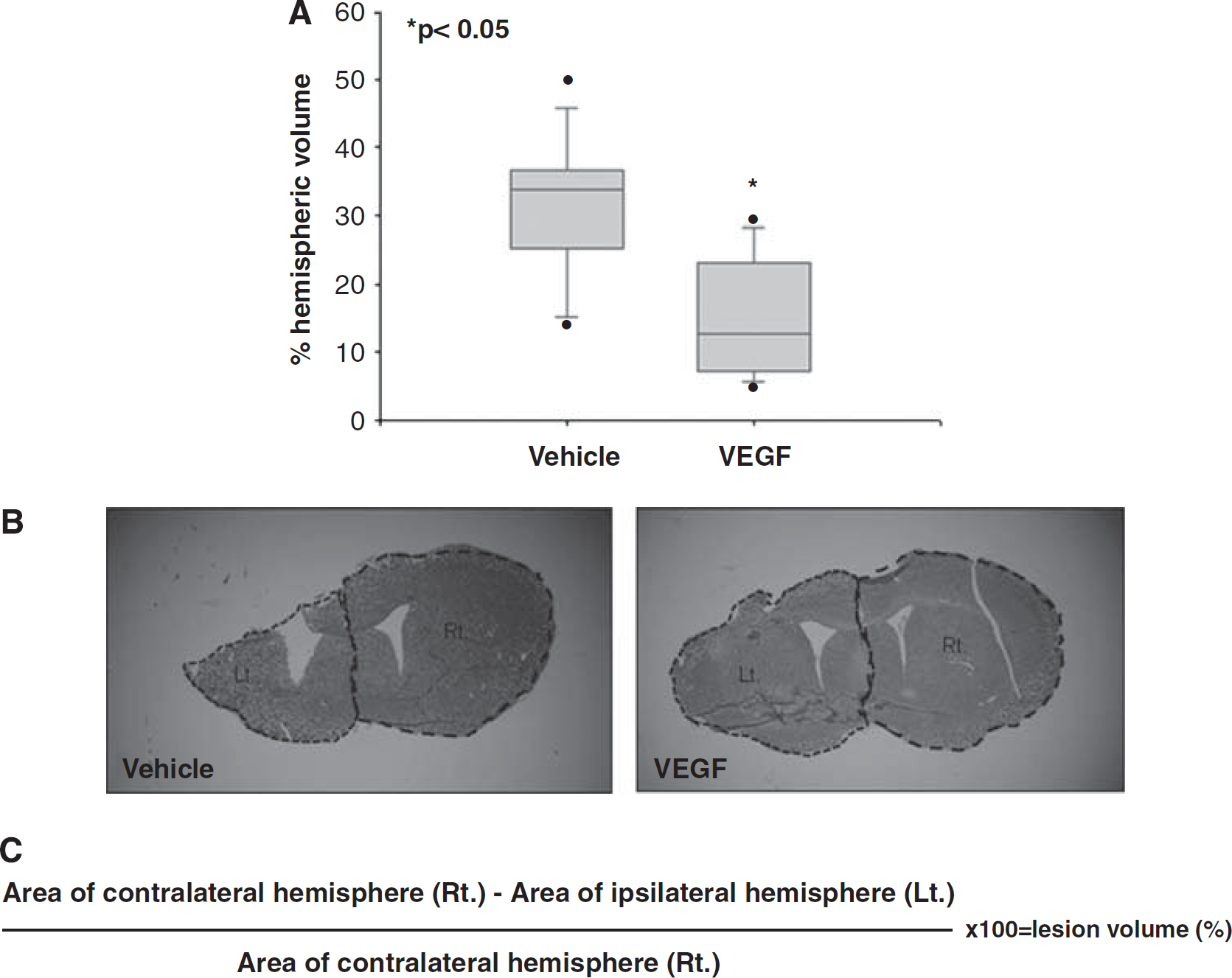
Vascular endothelial growth factor (VEGF) treatment significantly reduces lesion size as measured at 90 days after traumatic brain injury (TBI). Seven equidistant brain slices were stained with Giemsa and the volume of injured tissue was measured (
Immunohistochemistry:
Overall, 7 evenly spaced slices were counted for each brain between bregma +1.42 and bregma −0.8, and in each slice, 10 evenly spaced high-power magnification fields (× 400) in the entire area surrounding the infarct were counted. As the lesion core had already liquefied by 90 days after CHI, the area forming the outer boundary of the brain represents the border zone that survived. This area included mainly the cortex and subcortical tissue but not the striatum. Brain slices were double or triple stained for immunohistochemical evaluation using fate-specific antibodies that included rat anti-BrdU (marker for cell proliferation; 1:200; Accurate, New York, New York, USA), goat anti-doublecortin (DCX, marker for migrating neuroblasts, 1:200; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit anti-GFAP (marker for astrocytes, 1:200; Dako, Glostrup, Denmark), mouse anti-NeuN (marker for mature neurons; 1:100), rabbit anti Gal-C (marker for oligodendrocytes; 1:200) were all from Chemicom (Temecula, CA, USA), mouse anti-rat RECA1 (marker for blood vessels, 1:200; AbD Serotec, Oxford, UK) and mouse anti-CD31 (marker for endothelial cells, 1:100; Abcam Inc., Cambridge, MA, USA). Alexa 488 and Alexa 555 conjugates were used as secondary antibodies (1:200; Molecular Probes, Leiden, the Netherlands) and 46-diamidino-2-phenyl indole (Sigma Israel) was used to visualize nuclei.
Cell counting:
Immunopositive cells were counted using an epifluorescent Olympus (Tokyo, Japan) microscope in prespecified regions of interest including the SVZ, corpus callosum, and the area surrounding the lesioned cortex. Overall, 10 equidistant fields were counted per region of interest (30 fields per slide and 210 fields per brain at × 400 magnification). Confocal Z sections on an Olympus system were used to determine colocalization.
Western blot analysis:
An additional 21 mice underwent CHI as described above, were treated with VEGF (
Statistical Analysis
Analysis was performed with the SigmaStat software package (Systat, Richmond, CA, USA). Data are presented as mean ± s.e.m. as indicated in the legends. Values were compared using one-way analysis of variance followed by Student–Newman–Keuls posttest method.
Results
VEGF Significantly Increased Functional Gain after TBI
Neurologic deficits were measured at predetermined time points after CHI with the NSS. A more rapid and pronounced improvement in NSS was observed in VEGF-treated animals, compared with those treated with vehicle. The absolute NSS was decreased and the ΔNSS from days 1 to 90 was significantly greater in animals treated with VEGF (Figure 1). The differences in NSS were apparent as early as day 3 after TBI, suggesting a neuroprotective effect, and the difference continued to increase with time until the conclusion of the study. It is noted that the slope in NSS increased significantly after 14 days and the curves diverged significantly more at this time point similar to what was seen after cerebral ischemia (Shetty et al, 2004).
VEGF Decreases Lesion Volume after TBI
In this set of experiments, exogenous VEGF was administered through mini-osmotic pumps for 1 week after TBI. A significant 50% reduction in lesion size was identified at 90 days after TBI in VEGF-treated mice (Figures 2A and 2B), implying a neuroprotective effect for exogenous VEGF.
VEGF Increases the Number of Newborn Cells after TBI
To assess the effect of VEGF on cell division, we measured the total number of newborn BrdU + cells was measured at 90 days after injury. An increase in the number of BrdU + cells was found in mice treated with VEGF at all regions of interest including the SVZ (2.5-fold increase), the corpus callosum (1.7-fold increase), and the area surrounding the lesioned cortex (1.7-fold increase) (Figure 3). At 90 days after TBI, most BrdU + cells were localized to the immediate perilesion area suggesting accumulation of newborn cells around the injured brain area. These results suggest a possible mitogenic or prosurvival effect of VEGF on dividing cells.
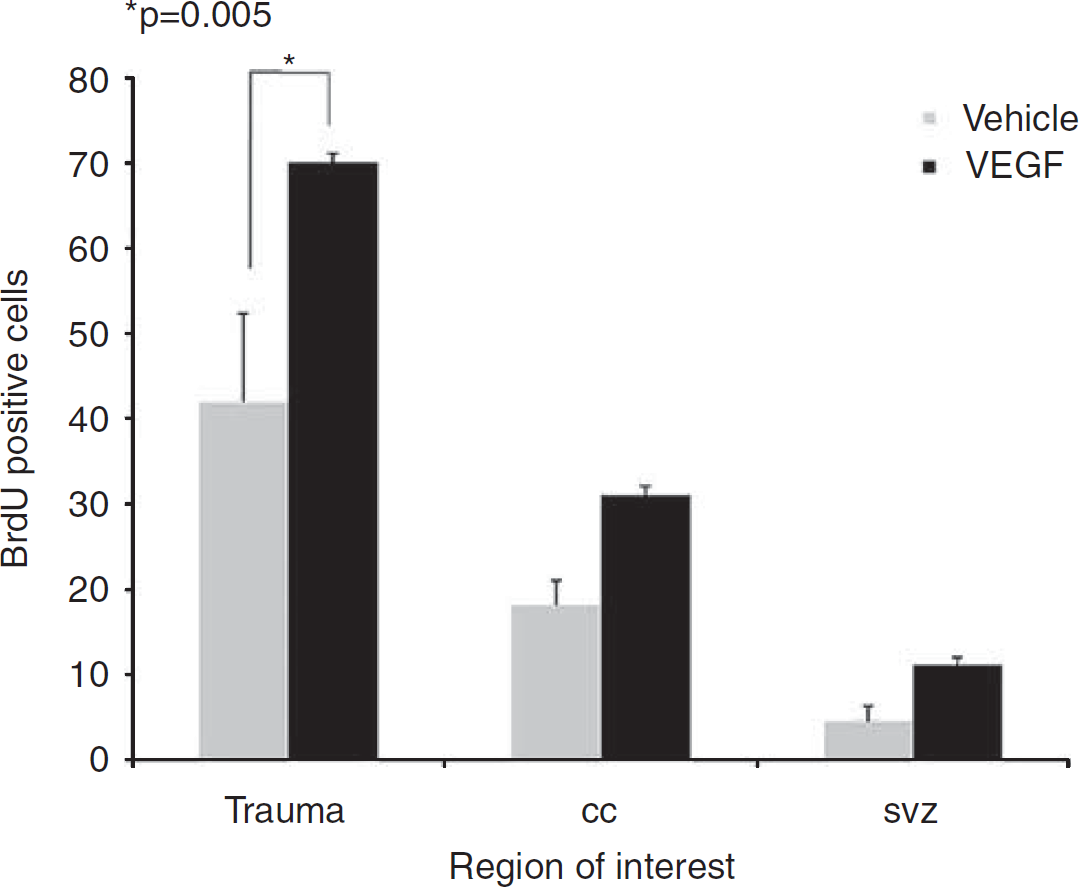
Vascular endothelial growth factor (VEGF) increases cell proliferation after traumatic brain injury (TBI). Animals were injected with 5-bromo-2-deoxyuridine (BrdU) after closed head injury (CHI). The absolute number of BrdU + cells was measured at 90 days after TBI at various regions of interest including the subventricular zone, the corpus callosum, and the cortex on the side of injury. Note that VEGF led to significantly more BrdU + cells in the cortex surrounding the injury.
VEGF Increases Gliogenesis and Neurogenesis after TBI
To determine the fates of newborn cells after TBI, we used immunohistochemical methods with double and triple labeling and confocal microscopy with Z sectioning for colocalization. Most newborn cells in VEGF-treated mice differentiated into astrocytes (GFAP+; Figures 4A and 4B) or oligodendroglia (GalC+; Figures 4C and 4D) and a smaller amount differentiated into newborn neurons around the SVZ region (DCX+; Figures 4E and 4F) and into mature neurons (NeuN+; Figures 4G and 4H). Most newborn astrocytes and neurons were located around the injured cortex and most newborn oligodendroglia were located in white matter tracts such as the corpus callosum suggesting site-appropriate differentiation of the newborn cells. Overall, the total numbers of newborn astrocytes and neurons at the lesion border were significantly higher in VEGF-treated mice (twofold increase compared with vehicle-treated mice,
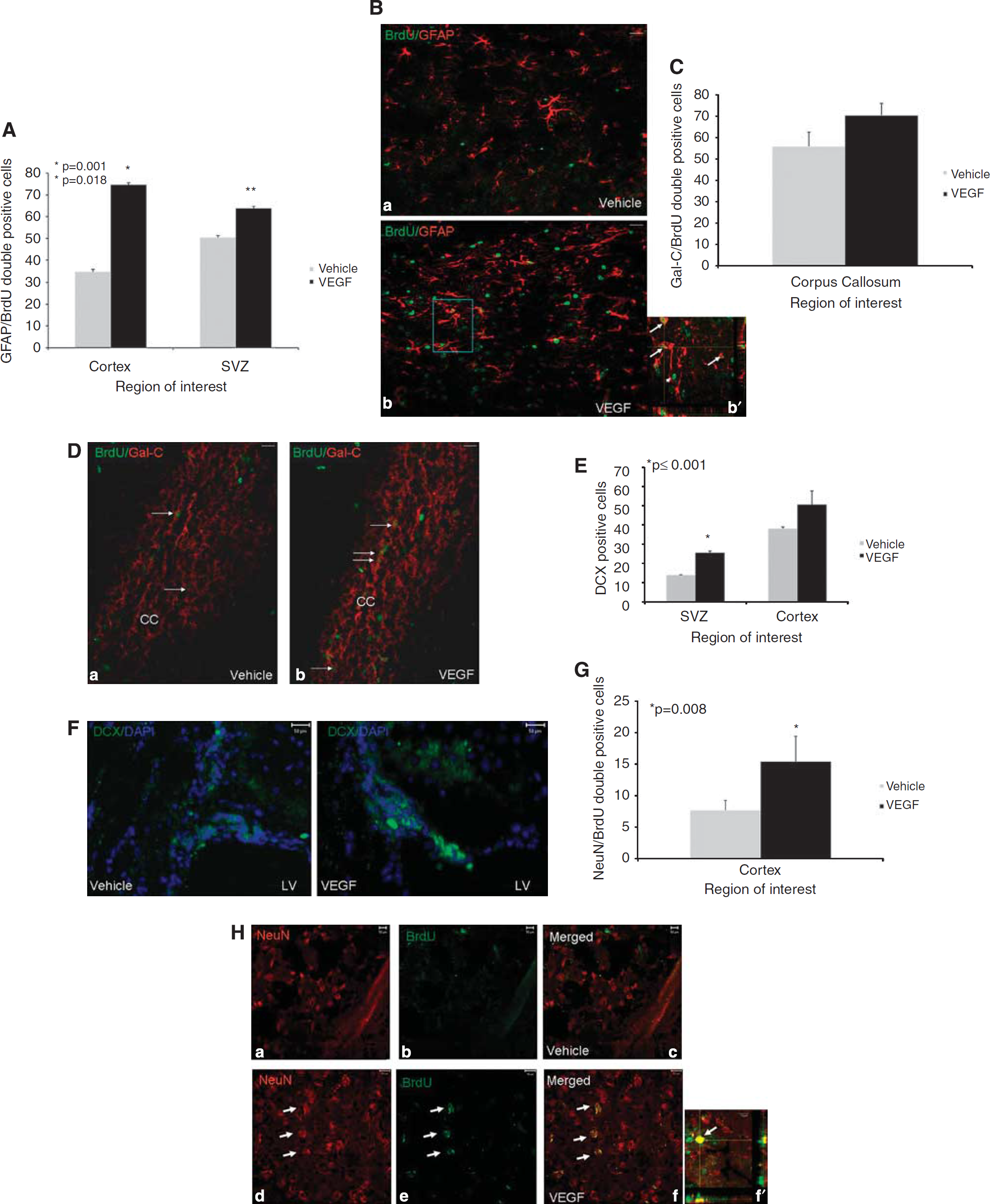
Effects of vascular endothelial growth factor (VEGF) on newborn cell differentiation. Animals underwent closed head injury (CHI) and were killed 90 days later. Newborn cell fate was determined with immunohistochemistry. Most newborn cells in animals treated with VEGF differentiated into GFAP + astrocytes in the perilesion cortex (
Although more newborn oligodendroglia were found in the corpus callosum of the VEGF-treated mice, compared with vehicle-treated animals, this difference did not reach statistical significance (70% versus 56%, respectively).
Taken together, these results suggest that VEGF may have an instructive role toward glial and neuronal differentiation but does not significantly affect oligodendroglial differentiation.
VEGF Increases Angiogenesis after TBI
At least part of the benefit afforded by VEGF could be attributed to its effect on angiogenesis. To examine the effect of exogenous VEGF on angiogenesis, we stained newly formed blood vessels with an antibody against the endothelial marker RECA1. A significant increase in the number of blood vessels (3.3-fold increase compared with vehicle-treated mice,
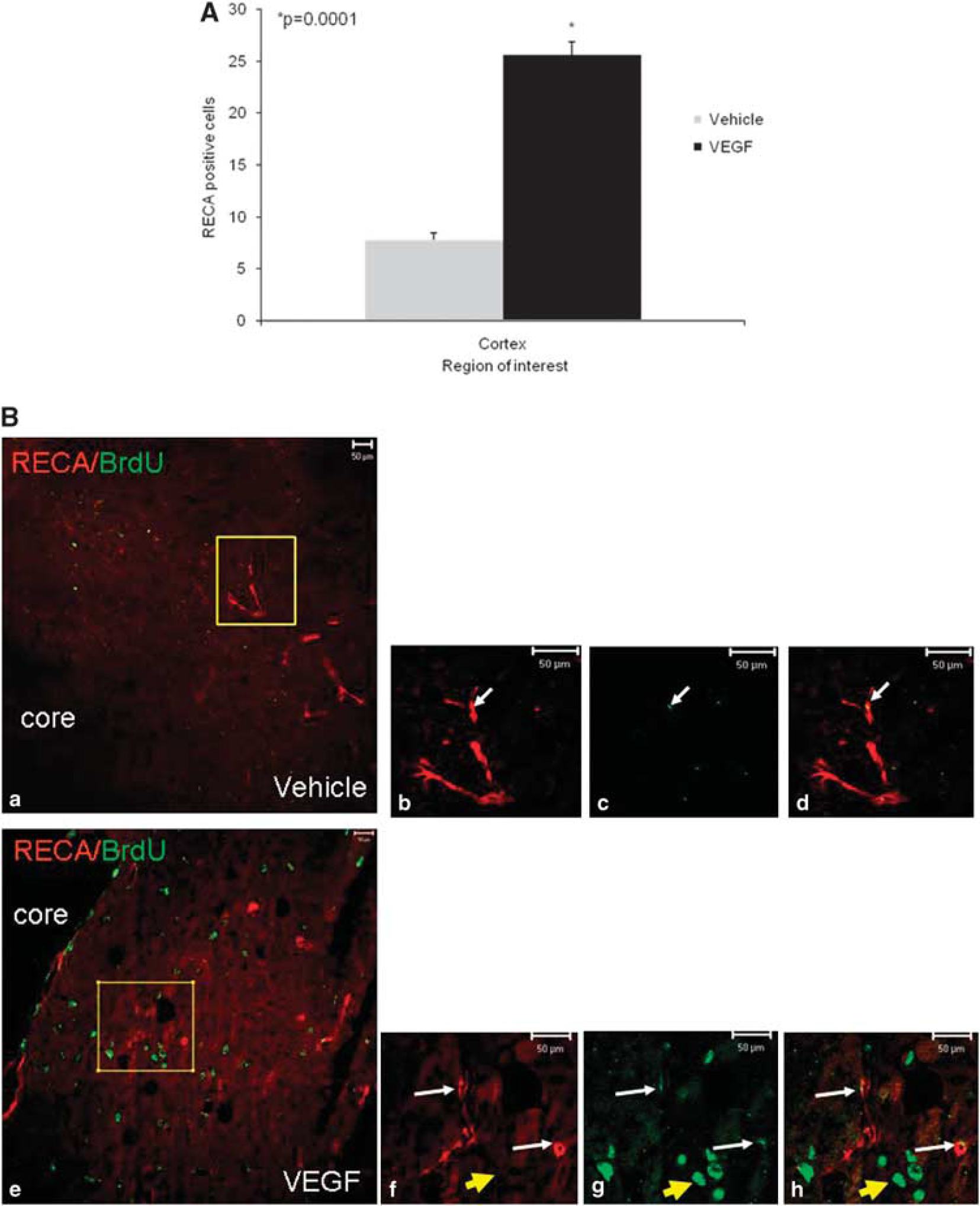
Vascular endothelial growth factor (VEGF) increases angiogenesis after traumatic brain injury (TBI). Animals were killed 90 days after TBI and the number of blood vessels was counted in the perilesion cortex. Significantly more vessels were seen in the cortical area surrounding the damage in VEGF-treated mice (
VEGF Activates the Survival Signaling Pathway by Increasing Phospho-Akt Levels
To elucidate the molecular mechanisms responsible for the salutary effects of VEGF, we tested treatment effects on the expression of several mediators of survival
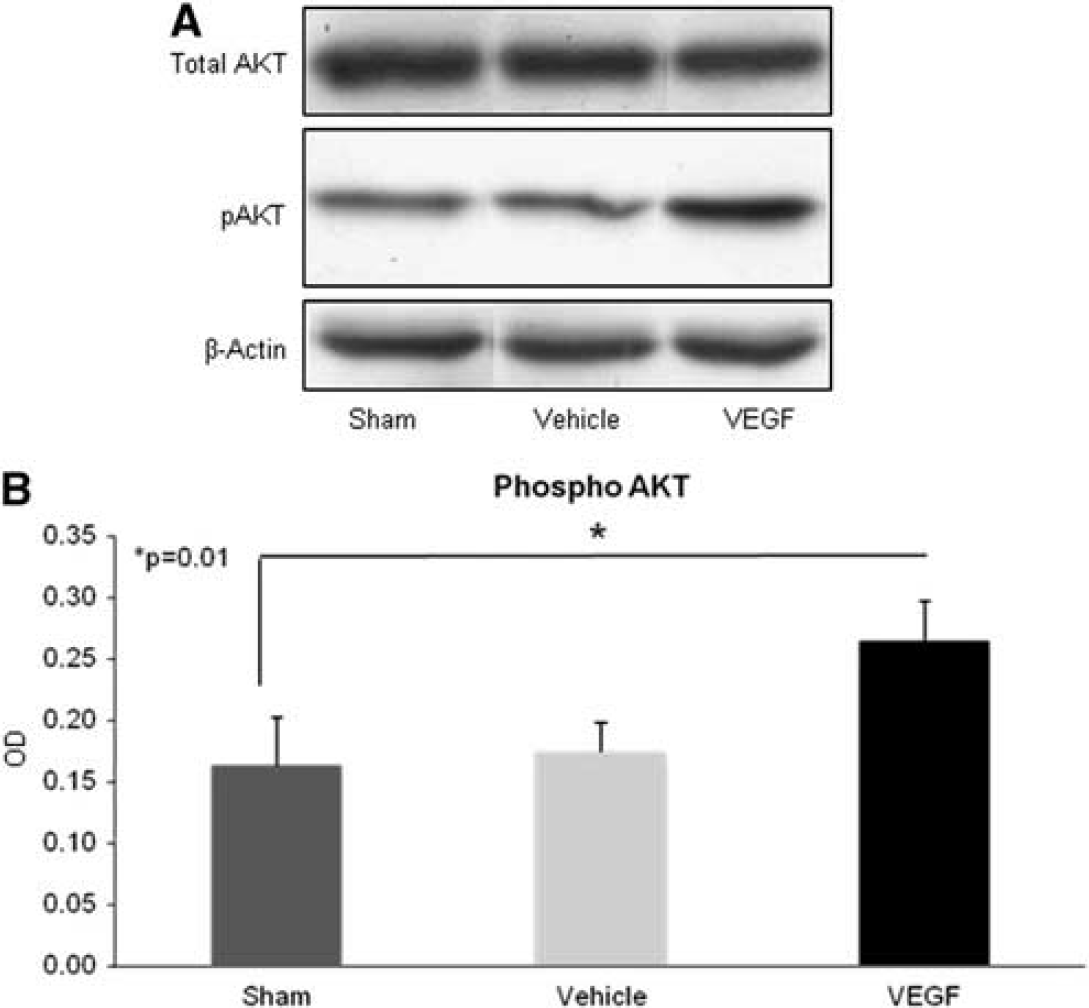
Vascular endothelial growth factor (VEGF) upregulates pAKT after traumatic brain injury (TBI). Animals underwent TBI, were treated with VEGF or vehicle for 7 days administered through mini-osmotic pumps, and were killed within minutes of end of therapy on day 7. pAKT levels were measured with western blots on cortical tissue. (
Discussion
The results of this study show that the administration of exogenous VEGF after TBI increases cell proliferation at the SVZ and leads to migration of cells through white matter toward the lesioned area. Larger proportions of newborn cells differentiated into astrocytes and neurons in VEGF-treated animals compared with controls. As expected, VEGF also significantly increased angiogenesis as shown by the increase in the number of blood vessels in proximity to the lesion. Importantly, the rates of functional motor and sensory recovery after TBI were significantly higher in animals treated with VEGF already at the early postinjury period (days 3 to 7) and increased even further from day 14 till the end of the follow-up period at 90 days. Thus, exogenous VEGF had a significant neuroprotective effect as evidenced by the early improvement in NSS, which proceeded up to the time of neurogenesis-related events. Moreover, the significantly decreased lesion volume observed at 90 days after TBI attests to the early neuroprotective effects of VEGF. However, the continuous improvement in functional deficits, namely, the decrease of NSS in the VEGF-treated mice, in contrast to the near-plateau levels reached in the control seen from day 14 onward, suggests an effect that is typically seen when neurogenesis and angiogenesis take place (Shetty et al, 2004).
Cells in the SVZ continue to proliferate for long periods of time after ischemic lesions (Leker et al, 2007; Thored et al, 2007), and this appears to be also true for TBI as shown in this work.
One of the intracellular pathways that are activated after VEGF binding is phosphorylation of the protein kinase Akt, a central player in a variety of cellular and physiologic events (Parcellier et al, 2008). It is now well established that the prosurvival effects of Akt after brain injury include antiapopototic, proangiogenic, and neuroprotective effects (Kilic et al, 2006; Shein et al, 2007). In this set of experiments, we found an early increase in pAkt in the brain shortly after VEGF administration, suggesting that the early neuroprotective effects of exogenous VEGF in TBI are probably mediated, at least in part, through induction of survival-signaling through pAkt, and a reduction in apoptosis (Kilic et al, 2006; Parcellier et al, 2008; Shein et al, 2007).
The functional status of TBI mice continued to improve at the later postinjury phase, although only a small fraction of the newly born cells differentiated into neurons. This implies that functional status does not entirely depend on neuronal replacement by newborn cells and could have several explanations. First, it was shown that newborn cells secrete a plethora of growth- and survival-promoting factors (Forsberg-Nilsson et al, 1998; Ogunshola et al, 2000; Shetty et al, 2005) and that these cells also have immunomodulatory effects that may improve the outcome (Ben-Hur, 2007) by influencing surviving neurons in the area surrounding the damage. Second, VEGF is an important modulator of angiogenesis (Morgan et al, 2007; Ogunshola et al, 2000), and it is therefore probable that at least part of the late beneficial effects on functionality are related to better perfusion in the area immediately adjacent to the trauma, which could lead to a better metabolic state in this area and thus to a better functional state. Third, it is possible that local factors within the area surrounding the lesion in TBI diminish neuronal differentiation. Such potential factors include myelin-associated factors and other signaling molecules such as NOGO (Zhao et al, 2007) and netrins (Cirulli and Yebra, 2007).
Vascular endothelial growth factor was shown to increase both neurogenesis and angiogenesis in the SVZ and to increase the size of the stem cell niche (Gotts and Chesselet, 2005). The niche is essential for stem cell survival and maturation by vascular recruitment and remodeling (Palmer et al, 2000). Surrounding the niche, neural stem cells were shown to proliferate and differentiate to a lesser extent in the absence of VEGF (Skold et al, 2006). Similar to what has been described in ischemic brain injury (Leker et al, 2007), we noticed an accumulation of newborn cells in the cortex surrounding the injury in treated animals, suggesting that VEGF also promoted migration of these cells toward their target site. The larger absolute numbers of newborn neurons and glia, as well as the percentage of BrdU + cells that differentiated into these cell types in VEGF-treated animals, suggest that VEGF may have an instructive role in the differentiation of neural progenitors. In future experiments, it would be important to test whether the use of VEGF could be combined with that of a proneuronal differentiating agent such as FGF2 or erythropoietin to yield even better outcomes as the result of further increments in the number of newborn neurons.
In conclusion, the result of our study shows that the administration of VEGF after TBI leads to proneurogenic, proangiogenic, and neuroprotective effects that are mediated through phospho-Akt signaling and that result in better functional outcome.