196. The effect of healthy aging on cerebral and systemic hemodynamic responses to posture change
B. Edlow1, M. Kim2, T. Durduran2, E. Buckley2, C. Zhou2, G. Yu2, M. Putt3, A. Yodh2, J. Greenberg1 and J. Detre1
1Neurology, University of Pennsylvania; 2Physics and Astronomy; 3Biostatistics, University of Pennsylvania, Philadelphia, Pennsylvania, USA
Objectives: Studies of healthy subjects have demonstrated that the systemic response to posture change, the baroreceptor reflex, is impaired with aging.1 In addition, aging is associated with lower baseline cerebral blood flow velocities,2 lower baseline cerebral perfusion3 and an increased incidence of orthostatic hypotension.4 The effect of healthy aging on cerebrovascular autoregulation, however, has yet to be fully elucidated. We used a novel, hybrid optical instrument to assess the impact of healthy aging on cerebral and systemic hemodynamics during posture change.
Methods: Diffuse Correlation Spectroscopy and Near-infrared Spectroscopy were used concurrently to measure relative cerebral blood flow (rCBF), total hemoglobin concentration (THC), microvascular oxyhemoglobin concentration (HbO2), and deoxyhemoglobin concentration (Hb) in the frontal lobe cortex of 60 healthy subjects ranging from age 20 to 78. Subjects were studied for 5 min at each of the following sequential positions: head of bed (HOB) at 30°, supine, standing, and supine. Noninvasive measurements of systemic hemodynamics, arterial oxygen saturation (O2Sat) and end-tidal CO2 (EtCO2) were performed continuously. A linear mixed effects model was used to quantify the effects of posture and age on each hemodynamic parameter, with age treated as a continuous variable.
Results: The study population had a mean age of 42.3 years and was comprised of 28 men and 32 women. Mean body mass index was 24.7 kg/m2, blood pressure 115.4/74.5 mm Hg, heart rate (HR) 71.3 beats per minute, and EtCO2 39.4 mm Hg. None of these baseline measurements varied significantly with age. Supine-to-standing posture change caused significant declines in rCBF (P<0.001), THC (P = 0.002), and HbO2 (P<0.001), and an increase in Hb (P = 0.001) across the age continuum (See Table). Diastolic blood pressure (DBP, P<0.001), mean arterial pressure (MAP, P = 0.018), and HR (P<0.001) increased, while EtCO2 (P<0.001) decreased upon standing. Aging was associated with a smaller magnitude of postural change in HR (P = 0.002), EtCO2 (P = 0.006), and HbO2 (P = 0.012), but no difference in the postural change in rCBF (P = 0.287).
Postural changes in cerebral and systemic hemodynamics (N = 60)
Larger magnitude of postural change with increasing age, P<0.05.
Smaller magnitude of postural change with increasing age, P<0.05.
Conclusions: Healthy aging does not alter postural changes in frontal cortical perfusion, despite age-related postural effects on HR, EtCO2, and HbO2.
756. Sexual dimorphism in the response of the middle cerebral artery to acute hyponatremia in the presence of vasopressin
K. Klapczynska and E. Kozniewska
Neurosurgery, Mossakowski Medical Research Centre, Polish Academy of Sciences, Warsaw, Poland
Objectives: Hyponatremia contributes to impaired recovery of neurosurgical patients after subarachnoid hemorrhage (SAH), brain injury or stroke. The decrease in serum sodium concentration in these patients is usually caused either by natriuresis or retention of water. Both effects depend on vasoactive hormones. One of them—vasopressin is a strong vasoconstrictor, particularly under pathological conditions. In our previous study we have found that isolated, male middle cerebral artery (MCA) dilates in endothelium-dependent manner in response to hyponatremia. Our present study addresses the question of the effect of hyponatremia on female MCA and the influence of vasopressin on the response of male and female MCA to hyponatremia.
Methods: MCAs harvested from the brains of the adult male and female (in estrus phase of the menstrual cycle) Wistar rats were used for this study. The vessels were mounted in the small organ chamber filled with 3-(N- morpholino) propanesulfonic acid (MOPS)—buffered saline solution and pressurized. The chamber was placed on inverted microscope equipped with videocamera for the recording of changes in the diameter of MCA. Hyponatremia was induced by decreasing intra—and extraluminal concentration of Na+ from 144 to 120 mmol/L. In the experiments with vasopressin (AVP), the peptide was added to the extravascular bath before induction of hyponatremia in a concentration of 15 pg/ml which is comparable with that found in the cerebrospinal fluid of patients after SAH.
Results: Lowering of Na+ concentration to 120 mmol/L resulted in a comparable endothelium-dependent dilation of male and female MCA by about 20% (P<0.05). Inhibition of NO/cGMP pathway abolished the effect of hyponatremia on male MCA and attenuated, but did not eliminate, hyponatremic response of female MCA. This suggests that hyponatremia stimulates production of another vasodilator, in addition to NO, in the endothelium of female MCA. Addition of AVP to the extraluminal bath before hyponatremia reversed hyponatremic response of male MCA to 14% constriction (P<0.005). This effect of AVP was eliminated by pretreatment of MCA with a selective V1a receptor antagonist and attenuated by a nonselective endothelin receptor antagonist. In the case of female MCA, pretreatment with AVP attenuated only the hyponatremic dilation.
Conclusions: These results demonstrate that hyponatremic dilation of male and female MCA depends on different endothelium-dependent factors. They also indicate that modulation of hyponatremic response of MCA by vasopressin is sexually dimorphic and suggest that vasopressin-induced hyponatremia may increase the risk of vasospasm in male subjects.
Acknowledgements: This study was supported by the grant N401 19032/3924 from the Ministry of Science and Higher Education.
283. When does angiogenesis accompanied with RBC flow develop in ischemic tissue after permanent MCA occlusion in C57BL/6J mice?
M. Tomita1, J. Tatarishvili1, H. Toriumi1, Y. Tomita1,2, M. Unekawa1, H. Hattori1 and N. Suzuki1
1Neurology; 2Department of Preventive Medicine for Cerebrovascular Disease, Keio University School of Medicine, Tokyo, Japan
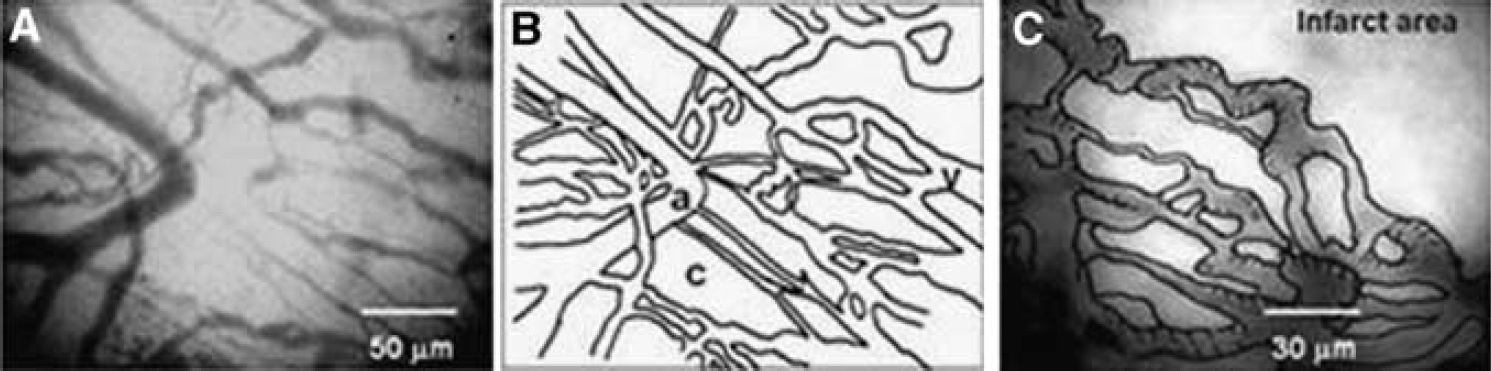
The time of angiogenesis in ischemic tissue may vary, depending upon levels of angiogenic factors (e.g., VEGF, FGFb, EGF, or TGF-b), or the degree of functional recovery (BBB or arterial regulation via muscles, glial sheathing and nerves). In this study, morphological development of new vessels and resumption of RBC flow after ischemia were targeted. Sixteen male C57BL/6J mice, 10 weeks old, under isoflurane anesthesia, with a closed skull window in the left parieto-temporal cortex, were observed every day after permanent MCA occlusion (sham-operated control n = 8). The temporo-parietal area centering arteriolo-arteriolar anastomoses (AAA) between MCA and ACA were observed. FITC-labeled RBCs was injected into the tail vein at each day. The mouse head was fixed to a head holder with a landmark bar, so that long-term changes of the same microvasculature could be observed every day. A laser scanning confocal fluorescence microscope was used for RBC tracking and RBC velocity and number measurements with our software KEIO-IS2 (Matlab domain)(Tomita M et alMicrocirculation 2008; 15:163). A halogen lamp (for general observation), laser beam at 488 nm (FITC for RBC) or at 533 nm (RITC for capillaries) was used as appropriate. The mouse was returned to the animal house after each day′s experiment and brought back to the laboratory next day. This procedure was repeated for a week. Day 0–1: MCAO induced immediate RBC disappearance (anoxia) in the core ischemic region, and to a lesser extent in the penumbra near the AAA. However, since RITC (plasma indicator) filled capillaries, the capillaries were somehow patent immediately after occlusion, implying occurrence of RBC sieving or plasma skimming. Such plasma passage became impeded within a few hours in the core. Day 1–2: Fragments of microvessels and tissue debris were observed, with abundant macrophages which phagocytosed RITC that had leaked from the vessels. Day 3: We recognized newly built vessels, mostly immature capillaries and reconstructed veins, around the marginal zone of the infarction. As shown in the Figure 1A and 1B (a rough sketch), straight capillaries that sprouted from the arteriolo-arteriolar cut end connected with dilated and tortuous vessels. Figure 1C shows revascularization of remodeled veins and a reactive part with many modular protrusions. Labeled RBC were flowing the capillaries and remodeled veins, with unsteady motions. Day 7: Revascularization similar to that reported by Tomita Y et al (JCBFM 2005; 25:858) was observed in cerebral ischemic tissue by 7 day after MCAO in mice. The ischemic core appeared to be liquefied in all 8 mice. In conclusion, angiogenesis was apparent in the marginal zone as early as day 3 after MCA occlusion in mice.
287. Feasibility of MRI-based measurements of cerebrovascular reactivity in an anesthetized animal model using a controlled cerebrovascular challenge
J.D. Winter1, S. Dorner2, J. Fisher3,4, G. Detzler1 and A. Kassner1,5
1Physiology and Experimental Medicine, The Hospital For Sick Children; 2Thornhill Research Inc., 3Department of Anesthesiology, University Health Network, University of Toronto; 4Department of Physiology; 5Department of Medical Imaging, University of Toronto, Toronto, ON, Canada
Objectives: Recent technological advances in the controlled manipulation of end-tidal partial pressure of CO2 (PetCO2) offer the ability to generate high-quality MRI-based maps of cerebrovascular reactivity (CVR).1 Using this novel methodology, researchers have demonstrated potential for identifying reduced CVR in patients with cerebrovascular disease.2 However, to better understand the basis for CVR abnormalities, we need controlled investigations in animal models. The purpose of this study was to demonstrate the feasibility and repeatability of MRI-based measures of CVR in an anesthetized animal model using standardized and repeatable PetCO2 manipulation.
Methods: We acquired blood-oxygen-level-dependent (BOLD) MRI data on a 1.5 T GE Signa MRI scanner using standard single-shot echo-planar imaging on three pigs (2 months of age). Imaging parameters included: echo time = 40 ms, repetition time = 2 s, field-of-view = 160 mm, resolution = 2.5 mm, slices = 16, slice thickness = 4.5 mm, volumes = 270 and acquisition time = 9 min. Animals were induced, orally intubated and mechanically ventilated. Anesthesia was maintained with a ketamine and midazolam infusion. The vascular stimulus consisted of four square-wave cycles of hypercapnia (∼55 mm Hg for 60 s) and normocapnia (∼40 mm Hg for 60 s). This was achieved using a computer-controlled gas delivery system in combination with a sequential gas delivery circuit (RespirAct™, Thornhill Research Inc., Toronto, Canada), which also continuously recorded PetCO2.1 To assess repeatability we acquired two data sets consecutively.
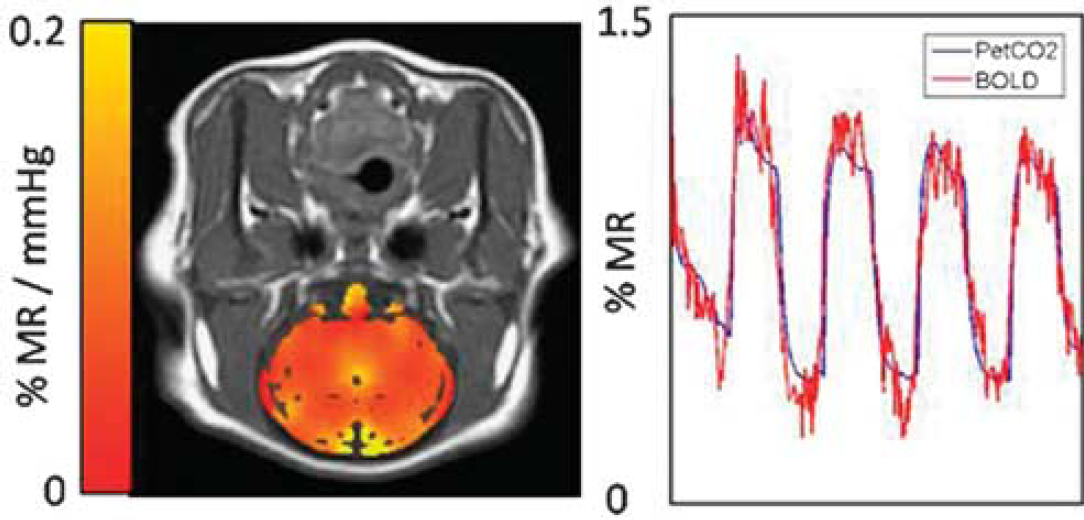
Data post-processing was performed offline using FSL and Matlab scripts. First, we matched the PetCO2 and BOLD MRI time courses by estimating the time delay between PetCO2 and whole-brain BOLD signal changes. Once in phase, we calculated CVR (% ΔMR signal/mm Hg CO2) on a pixel-by-pixel basis from the slope of the regression of % MR signal with PetCO2. To compare the test-retest CVR data, we automatically identified grey matter regions from anatomical scans and extracted mean grey matter CVR. We assessed repeatability using the coefficient of variation (CV).
Results:Figure 1 provides a representative CVR map and the BOLD MRI time series for all grey matter overlaid with the fitted PetCO2 model. Mean (±std. err.) grey matter CVR across all subjects was 0.064%±0.010%MR/mm Hg, and the mean CV was 8.9%±1.2%.
Conclusions: Using controlled PetCO2 manipulation we demonstrated the feasibility and repeatability of CVR measurements in a porcine animal model. To our knowledge, this is the first study to demonstrate CVR mapping in an anesthetized animal model using a controlled cerebrovascular stimulus, which may have implications for future animal studies investigating cerebrovascular impairment. Furthermore, we have illustrated the prospect of utilizing this CVR methodology in anesthetized human patients.
904. Regulation of vascular tone in the cerebral circulation is markedly impaired following cell-specific interference with PPARγ function
M. Modrick, S. Chrissobolis, C. Halabi, C. Sigmund and F. Faraci
University of Iowa, Iowa City, Iowa, USA
Objective: In vascular muscle would alter regulation of vasomotor tone in the cerebral circulation. Peroxisome proliferator-activated receptor-gamma (PPARγ) is a nuclear receptor that may play an important protective role in the vasculature. We examined the hypothesis that selective interference with PPARγ.
Methods: We studied basilar arteries from mice expressing a dominant negative mutation in human PPARγ (P467 L) under the control of the smooth muscle myosin heavy chain promoter (S-P467 L). We examined responses in vitro to stimuli that produce vasodilation via nitric oxide and cGMP (acetylcholine, nitroprusside), cAMP (forskolin), and potassium channels [cromakalim and potassium ion (K+)]. Because products of arachidonic acid metabolism may be endogenous activators of PPARγ, we also examined effects of arachidonic acid on vascular tone.
Results: In non-transgenic controls, all agonists produced dilation of the basilar artery. In S-P467 L mice, responses to acetylcholine, nitroprusside, forskolin, and activators of K+ channels were reduced by 40%–70% compared to controls (P<0.01). For example, vasodilation to K+ (10 mmol/L) was 67±6 and 22%±4% in control (n = 9) and S-P467 L (n = 6) mice, respectively (P<0.01). Arachidonic acid (1 μmol/L) dilated arteries from non-transgenic and S-P467 L mice by 63±3 and 20%±3%, respectively (P<0.01). Arteries from both groups responded similarly to the calcium channel agonist Bay K8644 and the calcium channel antagonist nifedipine and dilated fully to papaverine.
Conclusions: These data provide the first evidence that cell-specific interference with the function of PPARγ in vascular muscle produces marked, but selective, impairment of diverse vasodilator pathways. Thus, normal PPARγ function in vascular muscle plays an essential role in regulation of cerebral vascular tone.
951. VEGF rescues loss of myogenic tone induced by hypoxic-ischemia in neonatal rat middle cerebral arteries
S. Charles1, S. Butler1, J. Abrassart1, J. Williams1, B. Tone2, S. Ashwal2 and W. Pearce1
1Center for Perinatal Biology; 2Department of Pediatrics, Loma Linda University School of Medicine, Loma Linda, California, USA
Objectives: Whereas vascular injury undoubtedly contributes to cerebral infarct development following neonatal hypoxic-ischemic insults, the direct effects of such insults on cerebrovascular contractility remain poorly understood due largely to the technical challenges involved in such measurements. Nonetheless, better understanding of how hypoxic-ischemic insults compromise neonatal cerebrovascular contractility could greatly facilitate management of many NICU infants. In particular, it is important to know if neonatal ischemic vascular injury is irreversible, or if vascular function can be restored, and if so, how. The present study examines the hypothesis that neonatal ischemia compromises contractility by stimulating dedifferentiation of contractile smooth muscle, and that this process can be mitigated or reversed by treatment with the angiogenic growth factor, VEGF.
Methods: Sprague-Dawley rat pups at postnatal day 10 were anesthetized and the left common carotid was permanently ligated, as previously described.1 After a 90-min post-surgical recovery the pups were exposed to 8% oxygen for 90 min, then allowed to recover for 90 min at which time the cerebral arteries were harvested. The brains were stained with TTC to determine infarct volume, as previously described.1
Contractile characteristics of control and ischemic middle cerebral arteries were determined immediately after sacrifice, and after 48 h of organ culture. In organ culture, arteries were serum starved in DMEM in the presence or absence of 30 ng/ml VEGF. The contractility protocol measured in cannulated arteries the pressure-induced (myogenic) and potassium-induced changes in diameter at 20, 40, 60, and 80 mm Hg of transmural pressure. After contractility measurements, segments were sectioned and imaged following fluorescent immunohistochemical staining for smooth muscle α-actin and Myosin Light Chain Kinase (MLCK) to identify smooth muscle phenotypic changes.
Results: Before organ culture, ischemic injury completely ablated both stretch-induced and depolarization-induced contractility; at 20, 40, 60, and 80 mm Hg pressure-induced diameter differences averaged 39, 51, 58, and 75 μm in control arteries but only 5, 4, 2, and 0 μm in ischemic arteries. Organ culture with VEGF significantly improved contractility in the ischemic arteries; at 20, 40, 60, and 80 mm Hg diameter differences averaged 83, 79, 72, and 18 μm in control arteries and 45, 23, 4, and 1 μm in ischemic arteries. Fluorescent immunohistochemistry revealed that ischemia potently stimulated smooth muscle proliferation, as indicated by increased volume and density of staining for smooth muscle α-actin. VEGF treatment amplified this effect. Ischemia alone had little effect on MLCK expression, but ischemia plus VEGF dramatically enhanced MLCK expression, and by inference, the relative fraction of contractile smooth muscle cells.
Conclusion: The present results support the hypothesis that neonatal hypoxic-ischemia induces loss of vascular contractility via stimulation of contractile dedifferentiation in arterial smooth muscle. The data further suggest that this effect can be ameliorated by VEGF, which stimulates smooth muscle proliferation but also promotes expression of MLCK, a predominant regulatory contractile protein and marker for contractile, as opposed to proliferative, smooth muscle.