
Abstract
Xenobiotic efflux pumps at the blood—brain barrier are critical modulators of central nervous system pharmacotherapy. We previously found expression of the ligand-activated nuclear receptor, pregnane X receptor (PXR), in rat brain capillaries, and showed increased expression and transport activity of the drug efflux transporter, P-glycoprotein, in capillaries exposed to PXR ligands (pregnenolone-16α-carbonitrile (PCN) and dexamethasone) in vitro and in vivo. Here, we show increased protein expression and transport activity of another efflux pump, multidrug resistance-associated protein isoform 2 (Mrp2), in rat brain capillaries after in vitro and in vivo exposure to PCN and dexamethasone. The phase-II drug-metabolizing enzyme, glutathione S-transferase-π (GSTπ), was found to be expressed in brain capillaries, where it colocalized to a large extent with Mrp2 at the endothelial cell luminal plasma membrane. Like Mrp2, GSTπ protein expression increased with PXR activation. Colocalization and coordinated upregulation suggest functional coupling of the metabolizing enzyme and efflux transporter. These findings indicate that, as in hepatocytes, brain capillaries possess a regulatory network consisting of nuclear receptors, metabolizing enzymes, and efflux transporters, which modulate blood—brain barrier function.
Introduction
Pharmacotherapy of central nervous system (CNS) disorders is critically limited by the brain capillary endothelium that constitutes the blood—brain barrier. Although originally thought to be a passive physical barrier, we now know that the blood—brain barrier is a complex, dynamic structure with selective active components. The molecular basis of the selective barrier is a group of ATP-driven efflux transporters, such as P-glycoprotein and multidrug resistance-associated proteins (Mrps). Acting as the ‘first line of defense,’ drug efflux transporters limit xenobiotics, including a large number of CNS drugs, from penetrating into the brain. Thus, one strategy to improve drug delivery to the brain and increase drug levels in the CNS is to manipulate the expression and function of drug efflux transporters at the blood—brain barrier.
In liver, the ligand-activated nuclear receptor, pregnane X receptor (PXR, NR1I2), regulates the expression of a number of target genes involved in xenobiotic metabolism and efflux (Francis et al, 2003). Phase-I metabolizing enzymes regulated by PXR include CYP3A, CYP2B, CYP2Cs, and CYP7A (Wang and LeCluyse, 2003); phase-II enzymes include several UDP-glucuronosyltransferases (UGTs), sulfotransferases (SULTs), as well as glutathione S-transferases (GSTs) (Kliewer et al, 2002). Efflux transporters regulated by PXR include Mrp2, Mrp3, Oatp2, BSEP, and P-glycoprotein (Geick et al, 2001; Johnson et al, 2002; Kast et al, 2002; Kullak-Ublick and Becker, 2003; Teng et al, 2003). The pregnane X receptor is also expressed in nonhepatic tissues. We recently showed PXR expression in rat brain capillaries as well as PXR regulation of P-glycoprotein at the blood—brain barrier (Bauer et al, 2004). Exposing isolated rat brain capillaries to the PXR ligands, pregnenolone-16α-carbonitrile (PCN) or dexamethasone, increased protein expression and transport function of P-glycoprotein. Brain capillaries from rats dosed with PCN and dexamethasone also showed upregulation of P-glycoprotein expression and function. Using transgenic mice expressing human PXR (hPXR), we determined the consequences of increased P-glycoprotein expression at the blood—brain barrier and showed that hPXR activation in vivo tightened the barrier to a CNS-acting P-glycoprotein substrate, reducing the drug's central effect (Bauer et al, 2006).
In the present study, we have identified two additional targets at the blood—brain barrier for PXR regulation: (1) Mrp2, an ATP-driven efflux transporter that handles xenobiotics, xenobiotic phase-II metabolites, and endogenous metabolites (Kruh and Belinsky, 2003; Leslie et al, 2005) and (2) GSTπ, a phase-II metabolizing enzyme. We show that the PXR ligands, PCN and dexamethasone, increased the expression of Mrp2 in brain capillary membranes in vitro and in vivo. Mrp2 protein expression was also upregulated by chenodeoxycholic acid (CDCA), which is a high-affinity ligand for farnesoid X receptor (FXR), another ligand-activated nuclear receptor (Gnerre et al, 2004). We also present the first evidence for the expression and PXR regulation of GSTπ in brain capillaries. These findings suggest that, as in liver, a regulatory network of nuclear receptors, metabolizing enzymes, and efflux transporters is present at the blood—brain barrier. The elements of this network may provide targets to improve CNS pharmacotherapy.
Materials and methods
Chemicals
Pregnenolone-16α-carbonitrile, dexamethasone, CDCA, Texas Red (sulforhodamine 101 free acid), leukotriene C4 (LTC4), and DAPI (4′-6′-diamidino-2-phenylindole) were purchased from Sigma (St Louis, MO, USA). Rifampin was from Spectrum Chemical and Laboratory Products (Gardena, CA, USA). Mouse monoclonal M2III-6 antibody to Mrp2 and human polyclonal Mrp4 antibody were from Alexis-Axxora (San Diego, CA, USA), GSTπ antibody was from Stressgen (Victoria, Canada), Na+/K+-ATPase antibody was from Upstate Biotechnology (Lake Placid, NY, USA), GAPDH (glycerylaldehydephosphate dehydrogenase) and β-actin antibodies were from Abcam (Cambridge, MA, USA). PSC833 was a generous gift from Novartis (Basel, Switzerland); GF120918 was a generous gift from GlaxoSmithKline (Research Triangle Park, NC, USA). All other chemicals were of analytical grade and were obtained from commercial sources.
Animals, Dosing, and Tissue Isolation
Animals used were male, retired breeder Sprague-Dawley rats (500 to 600 g; Taconic, Germantown, NY, USA), male, retired breeder Wistar wild-type rats (500 to 600 g; Charles River Laboratories, Wilmington, MA, USA), male, retired breeder Wistar TR− rats lacking Mrp2 (500 to 600 g), male CB6F1 wild-type mice (25 to 35 g; Charles River Laboratories), and CB6F1 hPXR transgenic mice (25 to 35 g). The numbers of animals used are given in the figure legends. The animal-housing and -dosing protocols were approved by the Institutional Animal Care and Use Committees of NIEHS and the University of North Carolina and were in accordance with the NIH guidelines.
For in vitro experiments, rats were euthanized by CO2 inhalation and decapitated; brains were taken immediately for capillary isolation. For in vivo studies with rats, animals were dosed daily for 3 days with PCN (10, 25, or 50 mg/kg in corn oil) or dexamethasone (5 mg/kg in corn oil) by intraperitoneal injection; control animals received corn oil alone. For in vivo studies with mice, animals were dosed daily for 3 days with 50 mg/kg rifampin in 0.1% agarose by oral gavage (4 μL/g of 0.1% agarose, 12.5 μg/μL of 0.1% agarose; agarose should be maintained at 37°C to keep it in the liquid state); controls received agarose alone. Twenty-four hours after the last dosing, animals were euthanized by CO2 inhalation and brains were taken immediately for capillary isolation. Livers and kidneys were removed, snap-frozen in liquid nitrogen, and stored at −80°C until use.
Capillary Isolation
Brain capillaries were isolated as described previously (Bauer et al, 2007, 2006; Hartz et al, 2004, 2006). Animals were decapitated and brains were immediately put in ice-cold phosphate-buffered saline (PBS) (2.7 mmol/L KCl, 1.46 mmol/L KH2PO4, 136.9 mmol/L NaCl, 8.1 mmol/L Na2HPO4 supplemented with 5 mmol/L
Western Blotting
Tissue was homogenized in lysis buffer containing complete protease inhibitor (Roche, Mannheim, FRG). Samples were first centrifuged at 10,000 g for 30 mins, supernatants were then centrifuged at 100,000 g for 90 mins. Pellets (crude plasma membranes) were resuspended in buffer containing protease inhibitor and protein concentrations were determined. Western blots were performed using NuPage (Invitrogen, Carlsbad, CA, USA) electrophoresis and blotting system. After blocking, membranes were incubated overnight with antibody to Mrp2 (1:50), Mrp4 (1:500), GSTπ (1:500), Na+/K+-ATPase (1:10,000), GAPDH (1:1,000), or β-actin (1:1,000). Membranes were then washed and incubated for 1 h with the corresponding horseradish-peroxidase-conjugated ImmunoPure secondary antibody (1:15,000; Pierce, Rockford, IL, USA). Proteins were detected using SuperSignal West Pico chemoluminescent substrate (Pierce). Bands were visualized with a Bio-Rad Gel Doc 2000 gel-documentation system (Bio-Rad, Hercules, CA, USA).
Immunohistochemistry
Isolated rat brain capillaries adhering to glass coverslips were fixed for 15 mins with 3% paraformaldehyde/0.2% glutaraldehyde at room temperature. After washing with PBS, capillaries were permeabilized for 30 mins with 0.1% (v/v) Triton X-100 in PBS and blocked with PBS containing 1% BSA. Then, capillaries were incubated for 1 h at 37°C with antibody to Mrp2 (1:100), Mrp4 (1:100), or GSTπ (1:500). After washing (PBS and 1% BSA), capillaries were incubated for 1 h at 37°C with the corresponding Alexa Fluor 488- or 568-conjugated secondary IgG (all 1:1,000; Molecular Probes, OR, USA). Nuclei were counterstained for 15 mins with 1 μg/mL propidium iodide or DAPI. Negative controls for each treatment were processed without primary antibody. Immunofluorescence was visualized by confocal microscopy (Zeiss LSM 510 META inverted confocal microscope, × 40 water immersion objective, NA=1.2) and confocal images of 15 to 20 capillaries per treatment were acquired. To quantitate Mrp2 expression, luminal membrane Mrp2 immunofluorescence for each capillary was measured using ImageJ software (version 1.29). A 10 × 10 grid was superimposed on each image and measurements of capillary luminal plasma membrane Mrp2 immunofluorescence were taken along intersecting grid lines. The fluorescence intensity for each capillary was the mean of all measurements.
Luminal Texas Red Accumulation
Capillaries were transferred to incubation chambers with coverslip bottoms and incubated for 1 h at room temperature in PBS with 2 μmol/L Texas Red (sulforhodamine 101 free acid) or Texas Red plus 0.3 μmol/L LTC4, a compound known to inhibit Mrps (Miller et al, 2002, 2000; Russel et al, 2002), or with 1 mmol/L NaCN, a metabolic inhibitor. For each treatment group, confocal images (Zeiss LSM 510 META inverted confocal microscope, × 40 water immersion objective, NA=1.2) of 10 to 15 capillaries were acquired and luminal fluorescence intensity was measured using Zeiss imaging software, as described previously (Bauer et al, 2007; Hartz et al, 2004; Miller et al, 2000). Specific luminal Texas Red fluorescence was taken as the difference between total luminal Texas Red fluorescence and fluorescence in the presence of LTC4/NaCN.
Reverse Transcription-Polymerase Chain Reaction
Total RNA was isolated using TRIzol reagent (Invitrogen) and purified using the RNeasy Mini kit (Qiagen, Valencia, CA). Reverse transcription of total RNA was performed using the GeneAmp kit according to the manufacturer's protocol (Applied Biosystems, Foster City, CA, USA). Reverse-trancripted products were used for polymerase chain reaction (PCR) of rat Mrp4 (forward: 5′-TAAAATGGACACTGAACTAGC-3′, reverse: 5′-AATGGTGAGAACAGTGCA-3′, 35 cycles), rat FXR (forward: 5′-GACTGGTACTCTCCTGGACTCTATG-3′, reverse: 5′-CCTCATTCACTGTCTGATCCGCATG-3′, 40 cycles), and rat GSTπ (forward: 5′-GGAGGAGGTGGTTACCATAGATGTC-3′, reverse: 5′-GCAGGTCCAGCAAGTTGTAATCTGC-3′, 35 cycles). All primers were screened for specificity by using the PubMed BLAST database and were custom-synthesized by Qiagen. The PCR products were separated by agarose gel electrophoresis.
Statistical Analysis
Data are reported as mean±s.e.m. Appropriate Student's t-test was used to evaluate the statistical differences between controls and treated groups. Differences between means were considered to be statistically significant when P<0.05.
Results
Mrp2 Expression and Transport Function
Using a specific antibody (M2III-6), we previously immunolocalized the efflux pump, Mrp2, to the luminal plasma membrane of brain capillaries from pig, rat, and killifish (Miller et al, 2002, 2000). In brain capillaries from rats not expressing Mrp2 (TR− rats), no immunoreactivity was observed (Miller et al, 2000). Here, we confirm the functional expression of Mrp2 in isolated rat and mouse brain capillaries. Mrp2 protein expression was detected in western blots of plasma membranes isolated from rat brain capillaries (Figure 1A). Compared with renal brush-border membranes, Mrp2 expression levels were low in brain capillary membranes (compare signal strength and protein loading in Figure 1A). Indeed, although we could detect Mrp2 protein in isolated brain capillary membranes, we could not detect any signal in western blots of whole-brain homogenate, capillary-depleted brain homogenate, and brain capillary lysates from rat (data not shown). The ability of the antibody to specifically detect Mrp2 was verified in experiments with membranes from liver and brain capillaries of wild-type and TR− rats (Figure 1B). Note that in both tissues, expression of P-glycoprotein was not altered when Mrp2 was deleted.
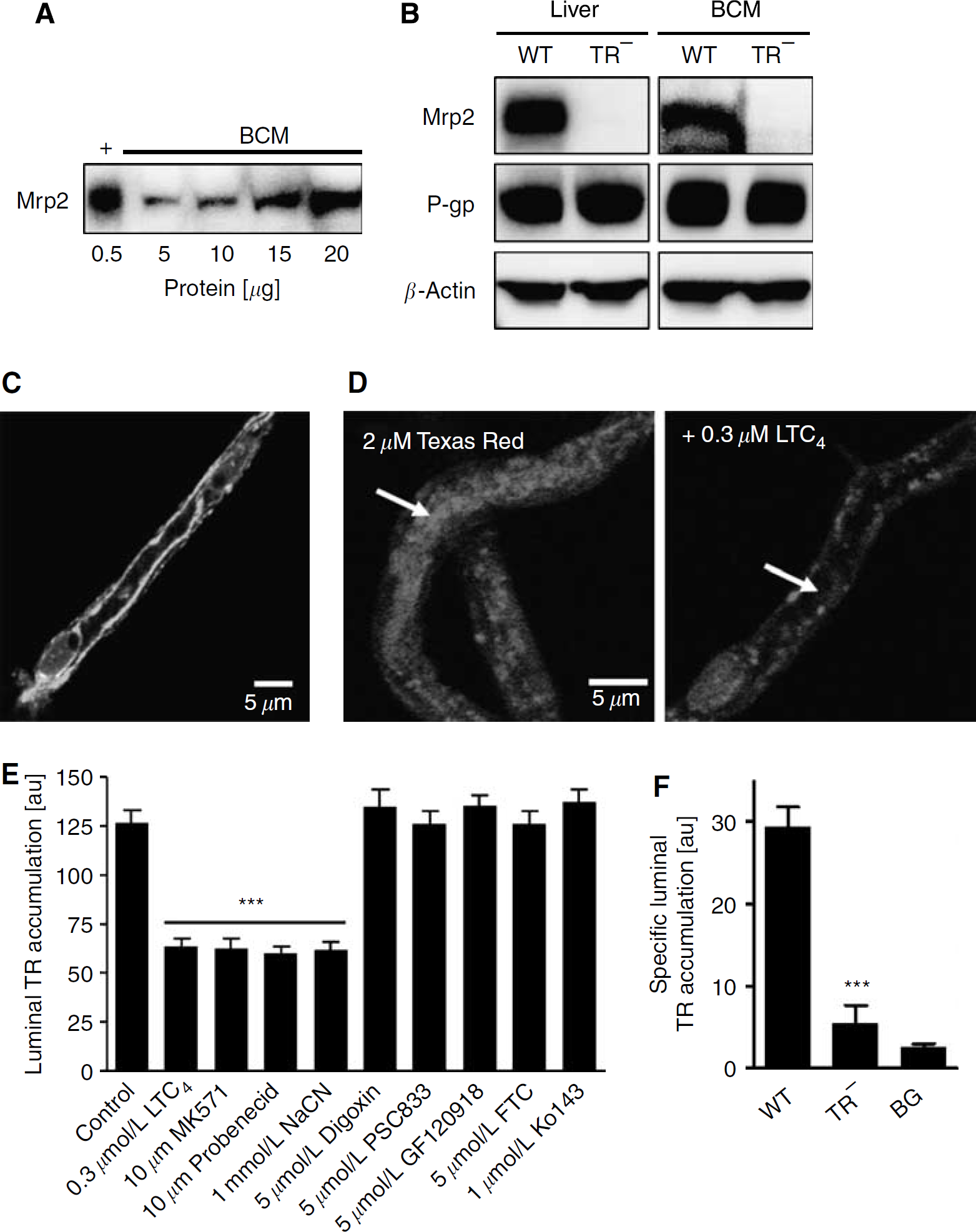
Multidrug resistance-associated protein isoform 2 (Mrp2) is functionally expressed in rat brain capillaries. (
Expression of Mrp2 at the blood—brain barrier is a matter of controversy (Loscher and Potschka, 2005b). Mrp2 is an integral plasma membrane protein. This may explain the inability of other investigators to detect Mrp2 in samples of total-brain homogenate, brain capillaries, and isolated endothelial cells from cow and rat (Johnson et al, 2006; Zhang et al, 2000) and why we could detect Mrp2 protein only in brain capillary plasma membranes. Consistent with Mrp2 expression in brain capillaries and a role in efflux transport, we detected immunoreactive Mrp2 protein in the luminal membrane of isolated capillaries by immunostaining (Figure 1C). Negative controls, that is, capillaries that were not exposed to primary antibody, showed no immunofluorescence (data not shown).
Isolated rat brain capillaries can be maintained in simple physiologic saline for up to 8 h without loss of viability or transport activity (Bauer et al, 2004; Hartz et al, 2004). Previous experiments suggested that Mrp2-mediated transport in isolated brain capillaries could be assayed using confocal microscopy to measure luminal accumulation of Texas Red (sulforhodamine 101 free acid), a fluorescent organic anion (Miller et al, 2002, 2000). We have extended those initial studies to validate a specific functional assay for Mrp2. Figure 1D shows representative confocal images of rat brain capillaries after 1 h incubation (steady state) in medium with 2 μmol/L Texas Red (control, left image) or 2 μmol/L Texas Red plus 0.3 μmol/L LTC4, an Mrp inhibitor (right image). The image of control capillaries shows high accumulation of Texas Red within the luminal space; luminal accumulation was greatly reduced in the capillaries exposed to LTC4. Quantitation of steady-state luminal Texas Red accumulation showed that the organic anion transport inhibitors, LTC4, MK571, and probenecid and the metabolic inhibitor, NaCN, reduced luminal Texas Red levels by about 50% (Figure 1E). In contrast, inhibitors of transport mediated by P-glycoprotein (PSC833, digoxin, GF120918), breast cancer resistance protein (GF120918, FTC, Ko143), and Oatp2 (digoxin) were without effect. Luminal Texas Red fluorescence remaining after inhibition of transport probably reflects passive diffusion and nonspecific binding of the dye to the tissue (Bauer et al, 2007, 2006; Hartz et al, 2006). Finally, Figure 1F shows that LTC4-sensitive luminal accumulation of Texas Red was essentially absent in capillaries from TR− rats (note that luminal fluorescence in TR− rats was close to background fluorescence, indicating lack of transport). Thus, the difference between total luminal Texas Red fluorescence and fluorescence in the presence of LTC4 or NaCN represents Mrp2-mediated transport; it provides a measure of specific Mrp2 transport activity.
Mrp2 Regulation in Rat
We previously showed that exposing brain capillaries from rat and mouse to PXR ligands for 6 h increased the expression and transport activity of the drug efflux transporter, P-glycoprotein (Bauer et al, 2004, 2006). Figure 2A shows that exposing isolated rat brain capillaries to the rodent-specific PXR ligand, PCN, for 6 h also increased Mrp2 immunoreactivity measured in isolated capillary membranes. Immunoreactivity for another plasma membrane protein, Na+/K+-ATPase, used as the loading control, was not increased. Pregnenolone-16α-carbonitrile exposure also increased Mrp2-mediated transport. Steady-state luminal Texas Red fluorescence increased significantly in capillaries exposed for 6 h to 5 to 10 μmol/L PCN (Figure 2B). Only the specific, that is, LTC4- and NaCN-sensitive, component of Texas Red accumulation was affected. With 5 to 10 μmol/L PCN, specific transport had nearly doubled (Figure 2C). This increase was abolished by actinomycin D (Figure 2D), suggesting it was dependent on transcription.
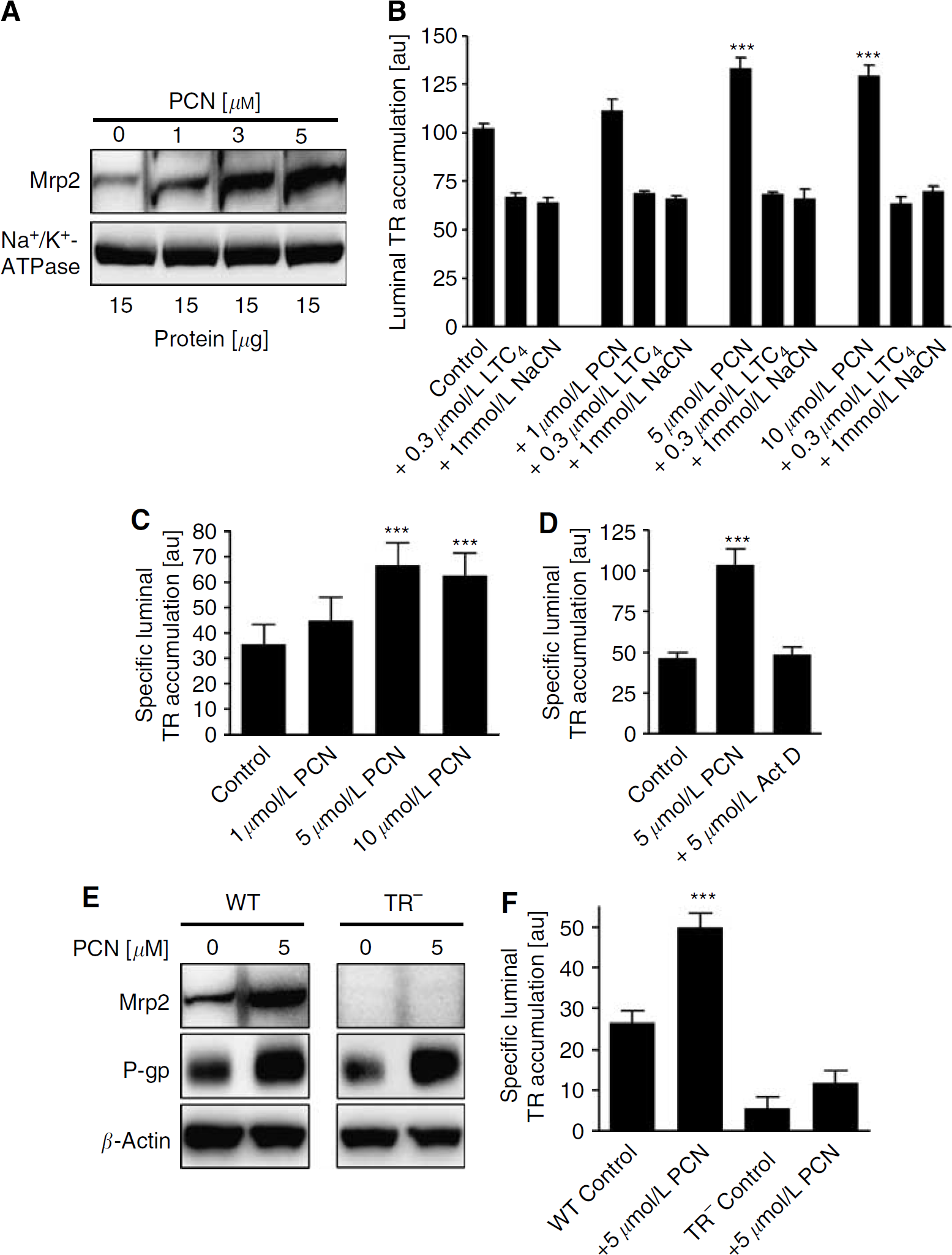
Pregnenolone-16α-carbonitrile (PCN) upregulates multidrug resistance-associated protein isoform 2 (Mrp2) in isolated rat brain capillaries from wild-type but not TR− rats. (
Parallel experiments were performed with brain capillaries from TR− rats and wild-type controls. In the wild-type rats, western blots showed that the expression of P-glycoprotein and Mrp2 increased with PCN exposure (Figure 2E, left panel). The increase in P-glycoprotein expression confirms previous results (Bauer et al, 2004). In TR− rats, the expected increase in P-glycoprotein expression was evident, but no Mrp2 signal was seen in either control or PCN-exposed capillaries (Figure 2E, right panel). Consistent with the protein expression data, PCN exposure nearly doubled the specific luminal accumulation of Texas Red in capillaries from wild-type rats (Figure 2F). In contrast, specific Texas Red accumulation was very low in capillaries from TR− rats and was not significantly increased by PCN exposure. Dexamethasone, a glucocorticoid commonly used to treat CNS inflammation, is also a PXR ligand, and we recently showed in rat brain capillaries that dexamethasone increased protein expression of Mrp2 in vivo (Bauer et al, 2004; Kast et al, 2002; Kliewer et al, 1998; Pascussi et al, 2000). Here, we confirm our previous results and show that dexamethasone increased Mrp2 protein and specific Texas Red accumulation in capillary lumens (Supplementary Information). Taken together, these in vitro experiments indicate specific increases in Mrp2 expression and transport activity in brain capillaries exposed to the PXR ligands, PCN and dexamethasone.
To determine the effect of PXR ligands on Mrp2 expression in vivo, we dosed rats with 50 mg/kg PCN or 5 mg/kg dexamethasone daily for 3 days by intraperitoneal injection. On day 4, we isolated brain capillaries and measured Mrp2 protein expression and Texas Red transport. Livers and kidneys were also removed and crude plasma membranes were isolated for western blot analysis. In agreement with previous studies (Bauer et al, 2004; Demeule et al, 1999; Salphati and Benet, 1998), Mrp2 expression increased in plasma membranes from kidneys and livers of animals dosed with PCN or dexamethasone (Figure 3A). Importantly, Mrp2 expression also increased in membranes isolated from rat brain capillaries (Figure 3A), a finding that confirms our initial results (Bauer et al, 2004). Capillaries isolated from PCN- or dexamethasone-treated rats also showed significantly increased (about 1.6-fold) luminal membrane Mrp2 immunofluorescence compared with controls (Figure 3B and 3C). Quantitation of luminal membrane Mrp2 immunofluorescence revealed a 66% increase in the staining intensity of PCN-exposed capillaries and a 64% increase of dexamethasone-exposed capillaries (97.5±2.8 U for control capillaries versus 161.6±3.1 and 159.6±3.9 U for PCN- and dexamethasone-exposed capillaries, respectively; P<0.001). Consistent with this, PCN and dexamethasone dosing increased specific Texas Red accumulation in capillary lumens by 131%±9% and 126%±8%, respectively (Figure 3D).
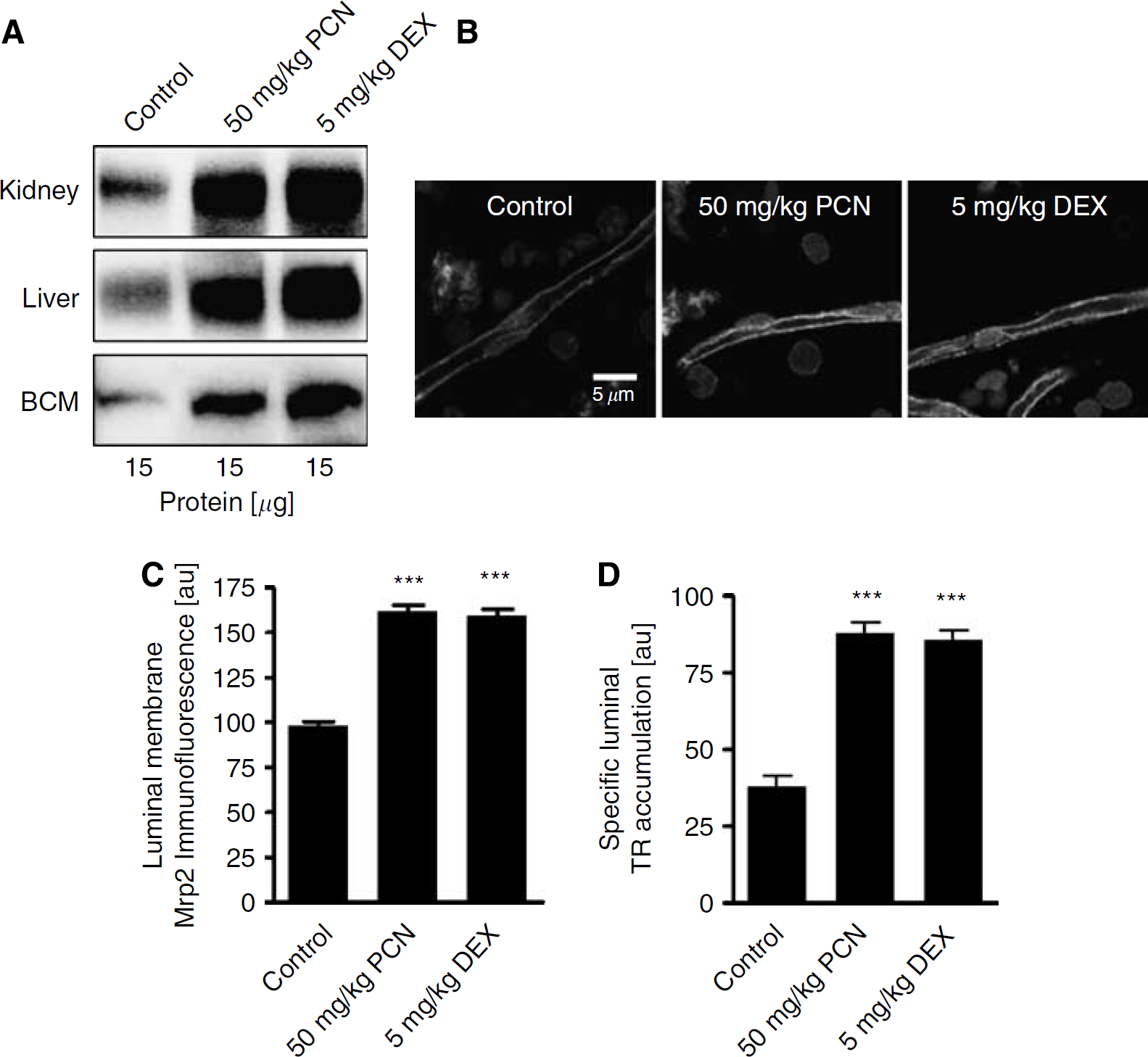
Pregnenolone-16α-carbonitrile (PCN) and dexamethasone (DEX) dosing upregulates multidrug resistance-associated protein isoform 2 (Mrp2) expression in vivo. (
Mrp2 expression in liver is known to be regulated by two other nuclear receptors, the constitutive androstane receptor (CAR) and FXR (Kast et al, 2002). Using reverse transcription (RT)-PCR, we detected CAR mRNA in liver, as reported previously (Maglich et al, 2002), in brain homogenate and brain capillaries (data not shown). In addition, we detected the expression of FXR mRNA in brain capillaries, brain homogenate, choroid plexus, kidney, and liver (positive control, Figure 4A). Consistent with FXR expression and function, a 6 h exposure of brain capillaries to the high-affinity FXR ligand, CDCA (Gnerre et al, 2004), increased Mrp2 protein expression in capillary plasma membranes (Figure 4B); a maximal effect was observed with 25 μmol/L CDCA. In agreement with western blotting, luminal membrane Mrp2 immunofluorescence increased significantly with both 15 and 25 μmol/L CDCA (Figure 4C). In isolated capillaries, CDCA also increased specific luminal Texas Red accumulation in a concentration-dependent manner (Figure 4D). Luminal fluorescence increased by 35%±17% with 5 μmol/L CDCA, 90%±17% with 15 μmol/L CDCA, and 122%±18% with 25 μmol/L CDCA. Taken together, these findings indicate that blood—brain barrier Mrp2 expression is also regulated by FXR.
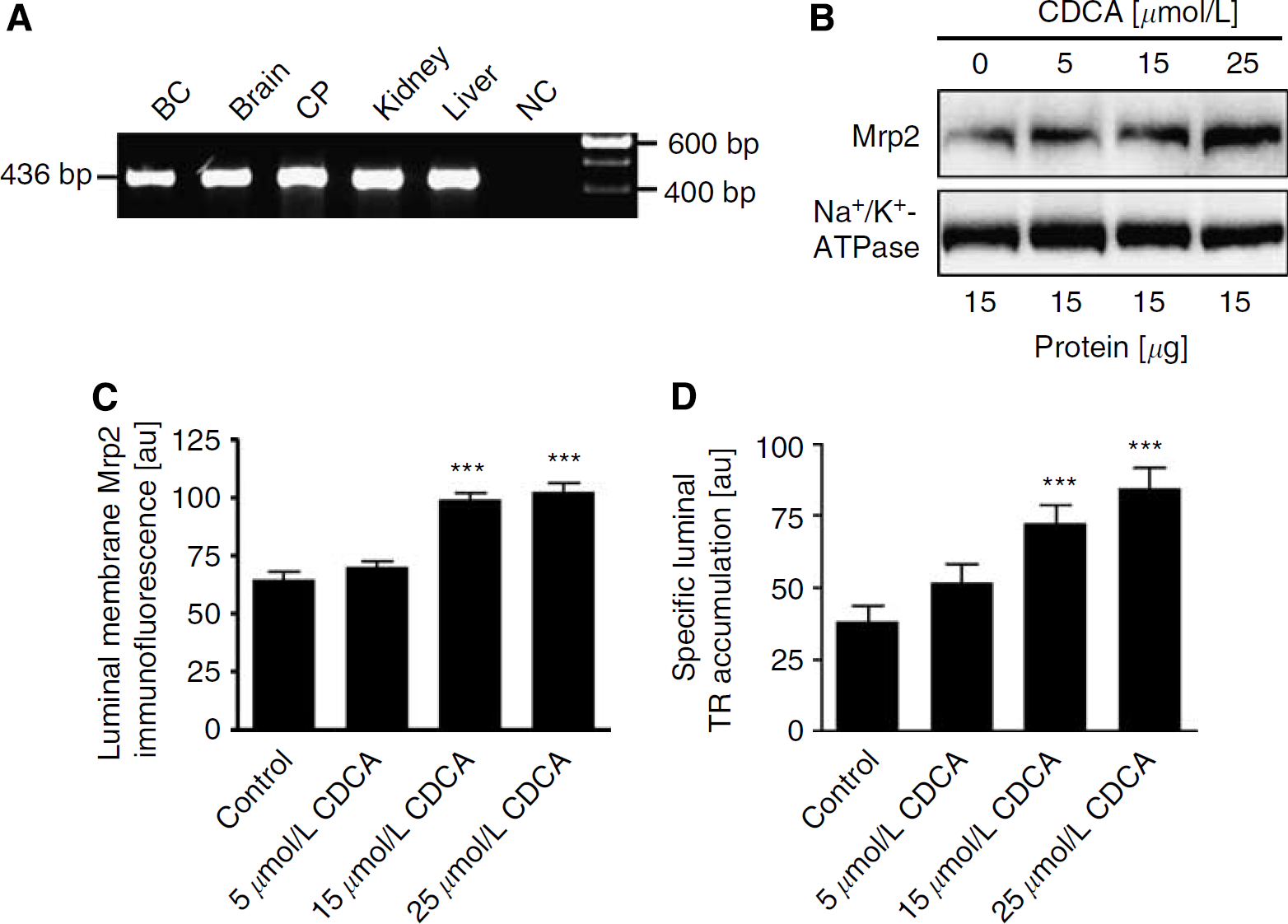
Chenodeoxycholic acid (CDCA) upregulates multidrug resistance-associated protein isoform 2 (Mrp2) in isolated rat brain capillaries. (
Lack of Mrp4 Regulation
Recently, Leggas et al (2004) showed the expression of another ABC-C family transporter, Mrp4, at the blood—brain barrier of mice. Using RT-PCR, we detected Mrp4 mRNA expression in multiple rat tissues, including isolated rat brain capillaries, brain homogenate, choroid plexus, kidney, and liver (Figure 5A). By western blotting, we detected Mrp4 protein in isolated capillary membranes, but not in capillary lysate, indicating enrichment of the transporter in the membrane (Figure 5B). Consistent with this and with previous findings (Zhang et al, 2004), we found Mrp4 immunofluorescence in both luminal and abluminal membranes (Figure 5C). Mrp4 staining was also observed in blood cells trapped within capillary lumens, which was expected based on previous studies (Jedlitschky et al, 2004). To determine whether Mrp4 expression was regulated by PXR, we dosed rats daily for 3 days with 50 mg/kg PCN or 5 mg/kg dexamethasone by intraperitoneal injection and isolated membranes from brain capillaries and kidneys (positive control for Mrp4 expression). Neither PCN nor dexamethasone dosing increased Mrp4 protein in kidney or brain capillary membranes (Figure 5D), indicating differential transcriptional regulation of Mrp2 and Mrp4.
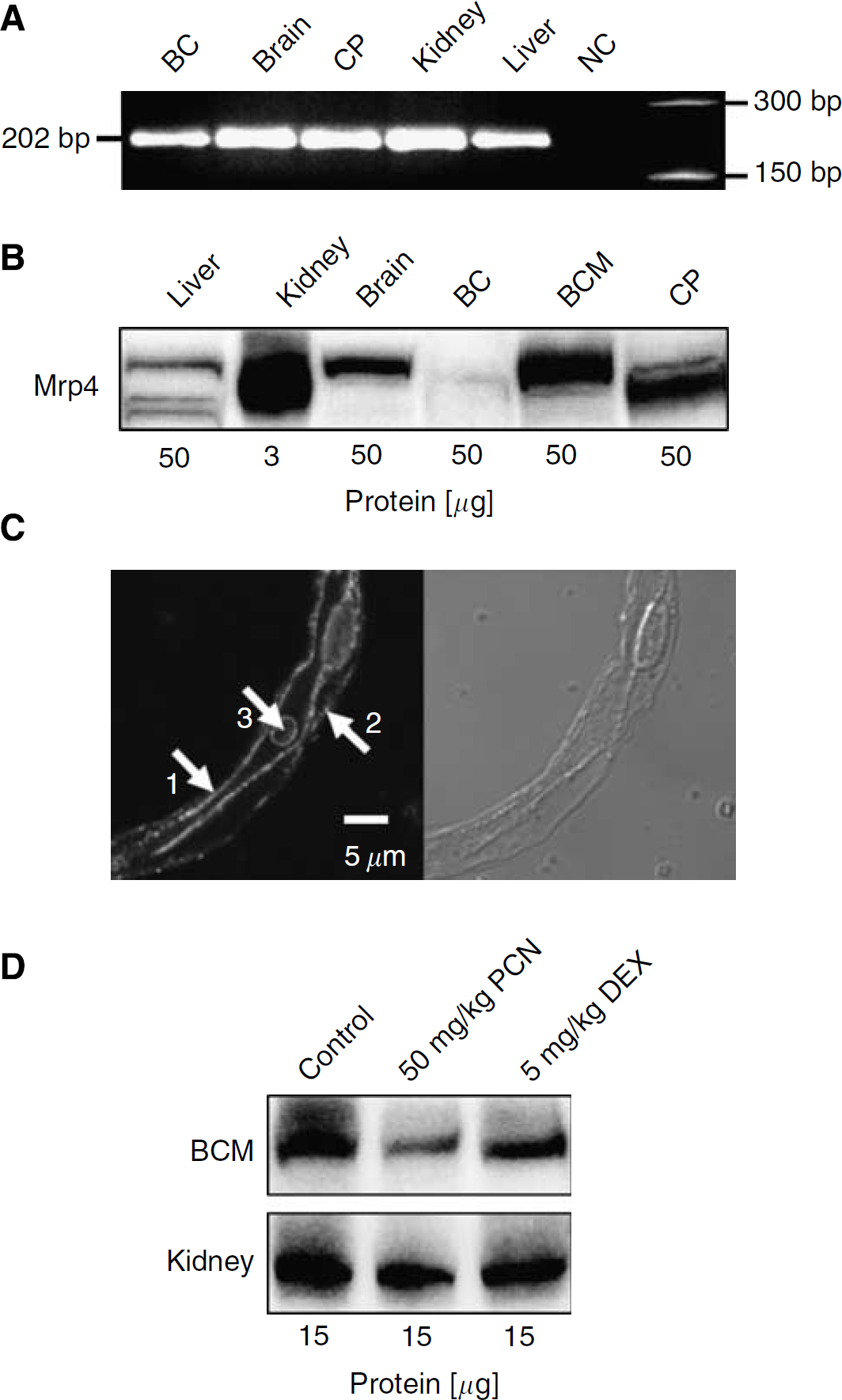
Multidrug resistance-associated protein isoform 4 (Mrp4) is not upregulated by pregnenolone-16α-carbonitrile (PCN) or dexamethasone. (
Glutathione S-Transferase-π Expression and Regulation
Several studies suggest metabolism-coupled elimination of xenobiotics by Mrp1 or Mrp2 in combination with the phase-II conjugating enzyme, GST (Depeille et al, 2004; Leslie et al, 2004; Smitherman et al, 2004). We detected mRNA and protein expression of GST isoform π in isolated rat brain capillaries (Figure 6A and 6B). In the western blot, the GSTπ signal was roughly equivalent in brain homogenate, total-brain capillary lysate, and brain capillary membranes (Figure 6B). In contrast, the signal for the cytosolic marker protein, GAPDH, decreased from brain homogenate to capillary lysate to brain capillary membranes. This suggests the association of a substantial fraction of total GSTπ with cellular membranes. To explore this possibility further, isolated brain capillaries were immunostained for both GSTπ and Mrp2. Figure 6C shows GSTπ staining (red) extended over cytoplasm and the luminal membrane of capillaries. As expected, Mrp2 staining (green) was only at the luminal plasma membrane. Overlaying red GSTπ and green Mrp2 immunofluorescence produced a merged image showing substantial colocalization (yellow) of the two proteins in the luminal plasma membrane (Figure 6C).
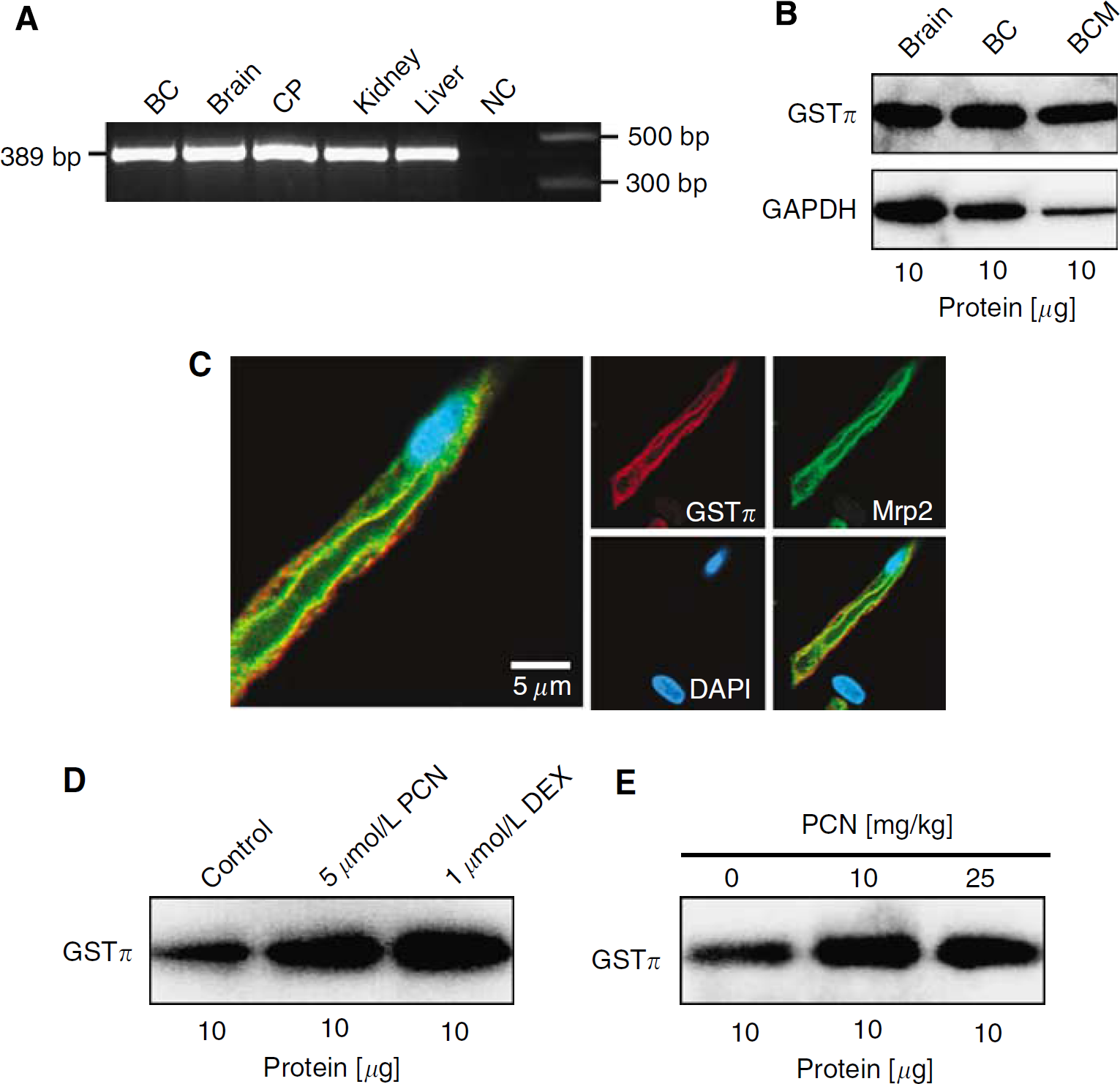
Glutathione S-transferase-π (GSTπ) is expressed in isolated rat brain capillaries and upregulated by pregnenolone-16α-carbonitrile (PCN) and dexamethasone. (
In liver, GSTπ expression is transcriptionally regulated by PXR (Kliewer et al, 2002). To determine whether GSTπ expression in brain capillaries was also regulated by PXR, we exposed capillaries to PCN and dexamethasone for 6 h, isolated capillary membranes, and measured GSTπ expression by western blotting. Figure 6D shows that exposure to 5 μmol/L PCN or 1 μmol/L dexamethasone increased GSTπ expression. Consistent with these in vitro results, GSTπ expression was also increased in capillary membranes isolated from rats dosed with 10 and 25 mg/kg PCN (Figure 6E).
Rifampin Effects in Human Pregnane X Receptor Transgenic Mice
The ligand-binding domain of PXR exhibits substantial species variation and this is reflected in the differences in ligand specificity of species (Kliewer et al, 2002; Moore et al, 2002). For example, PCN activates rodent PXR, but not hPXR. Conversely, rifampin is a potent ligand for hPXR, but not for rodent PXR. To show that PXR activation at the blood—brain barrier could potentially play a role in humans, we recently used a transgenic mouse expressing hPXR. We showed that in vivo dosing with rifampin, an hPXR ligand, increased P-glycoprotein expression at the blood—brain barrier and decreased methadone antinociception (Bauer et al, 2006). In these mice, the dose of rifampin used (50 mg/kg daily for 3 days) resulted in plasma levels that were equivalent to those found in patients undergoing a course of rifampin therapy. Figure 7 shows that this rifampin-dosing schedule also increased the expression of Mrp2 and GSTπ in brain capillary membranes from hPXR transgenic mice. As before (Bauer et al, 2006), P-glycoprotein expression was increased by rifampin. As in rat (Figure 5D), Mrp4 expression in mouse brain capillaries was not affected by PXR activation (Figure 7).
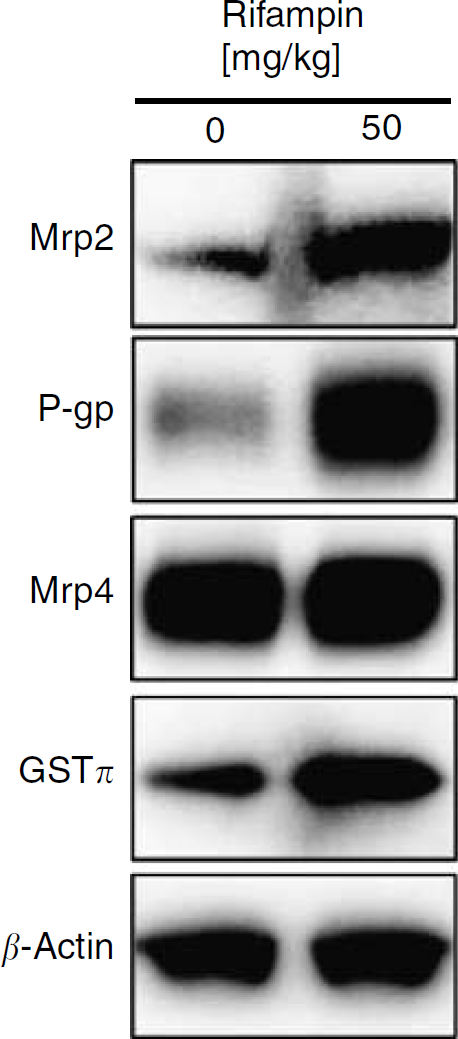
Rifampin upregulates multidrug resistance-associated protein isoform 2 (Mrp2) and P-glycoprotein in human pregnane X receptor (hPXR) transgenic mice in vivo. Western blots of the transporters, Mrp2, P-glycoprotein (P-gp), Mrp4, and the metabolizing enzyme, GSTπ, in brain capillary membranes of hPXR transgenic mice dosed with 50 mg/kg rifampin daily for 3 days by oral gavage; controls received vehicle alone. β-Actin was used as the loading control.
Discussion
We recently showed expression of the ligand-activated nuclear receptor, PXR, in rodent brain capillaries and showed that in vitro or in vivo exposure of brain capillaries to PXR ligands increased protein expression and transport function of the drug efflux transporter, P-glycoprotein (Bauer et al, 2004, 2006). One consequence of increased P-glycoprotein expression was tightening of the selective blood—brain barrier, as evidenced by reduced methadone antinociception (Bauer et al, 2006). The present study extends previous findings to other PXR targets and introduces two new concepts for the blood—brain barrier: (1) coordinated regulation of xenobiotic metabolism and excretory transport and (2) regulation through a network of ligand-activated nuclear receptors.
We show here that PXR regulates Mrp2 expression at the blood—brain barrier in rats and hPXR transgenic mice. In wild-type rats, Mrp2 immunoreactivity increased in membranes of brain capillaries that were exposed to the PXR ligands, PCN and dexamethasone. In agreement with previous studies, dosing rats with PCN or dexamethasone increased Mrp2 expression in kidney and liver (Bauer et al, 2004; Demeule et al, 1999; Salphati and Benet, 1998). Importantly, Mrp2 protein expression also increased in brain capillary membranes isolated from rats dosed with PCN and dexamethasone. A similar increase in Mrp2 immunoreactivity was found in brain capillary membranes from hPXR transgenic mice dosed with the hPXR ligand, rifampin. Consistent with these findings, an assay that measures Mrp2 transport function in intact rat capillaries showed that in vitro or in vivo exposure to PXR ligands significantly increased Mrp2 transport activity. In contrast, no increases were found when blots were probed for Mrp4.
In these in vitro and in vivo experiments with rats, the two drugs were equally effective when dexamethasone levels were an order of magnitude lower than PCN levels. We saw the same pattern in our previous study of P-glycoprotein regulation in rat brain capillaries (Bauer et al, 2004). These observations are interesting in that they could suggest the involvement of the glucocorticoid receptor in dexamethasone induction of P-glycoprotein and Mrp2. Indeed, using RT-PCR, we found a weak signal for the glucocorticoid receptor in rat brain capillaries (B. Bauer et al, unpublished data). Acting through both PXR and the glucocorticoid receptor, dexamethasone has been previously shown to regulate CYP3A4 in hepatocytes (Pascussi et al, 2000).
In our experiments, actinomycin D, an inhibitor of transcription, blocked the PCN-induced increase in Mrp2 transport activity, suggesting transcriptional activation (present study). However, with RT-PCR, we could detect at best only a weak signal for Mrp2 mRNA in capillaries, even though we found a strong signal in liver (data not shown). Similar observations have been made by several other groups (Berezowski et al, 2004; Sugiyama et al, 2003; Zhang et al, 2000), suggesting a low level of Mrp2 mRNA in brain capillaries. Thus, from the present results, it is not fully established to what extent PCN increases Mrp2 protein through transcriptional, posttranscriptional, translational, and/or posttranslational mechanisms. In rat liver, PCN treatment greatly increases Mrp2 protein with little or no effect on Mrp2 mRNA, and again it is not clear to what extent each mechanism contributes (Johnson et al, 2002; Jones et al, 2005). Certainly, as we used total membrane fractions for western blotting, increased Mrp2 levels reflect newly synthesized protein, suggesting that increased translation is involved. However, the exact mechanism(s) underlying the increase in Mrp2 protein levels remains to be determined.
Previously, Leslie et al (2004) showed plasma membrane colocalization of Mrp1 and the phase-II drug-metabolizing enzyme, GSTπ, in the H69AR cell line and found that functional GSTπ was required for Mrp1-mediated transport. In the present study, we immunolocalized GSTπ to the cytoplasm and luminal plasma membrane of brain capillary endothelial cells; to a large extent, GSTπ colocalized with Mrp2 in the membrane. Moreover, consistent with regulation by PXR, GSTπ protein expression increased in membranes from rat brain capillaries exposed to PCN or dexamethasone, in capillary membranes from rats dosed with PCN and in hPXR transgenic mice dosed with rifampin. Indeed, GSTπ and Mrp2 upregulation occurred in parallel, suggesting a coordinated regulation of phase-II metabolism and phase-III efflux transport. Membrane colocalization and coordinated regulation suggest, for the first time, the coupling of a metabolizing enzyme (GSTπ) with an efflux transporter (Mrp2) at the blood—brain barrier.
The role of metabolizing enzymes as an element of the blood—brain barrier is essentially unexplored. There is some evidence for the expression of CYP1A and CYP2B isoforms in cultured brain capillary endothelial cells (Chat et al, 1998), and the present results indicate GSTπ protein expression in rat and mouse brain capillaries. Although a comprehensive survey of enzyme gene expression has yet to be completed, we have obtained evidence for mRNA expression of both phase-I and phase-II enzymes in rat brain capillaries (Cyps 1a2, 2b, 3a23, 7a1, 27a1, GSTπ, GSTα, Sult1a1, Sult1a2, Ugt1a1; B. Bauer et al, unpublished data). We speculate that metabolizing enzymes, in addition to transporters, could contribute to blood—brain barrier function as a ‘second line of defense’ in two ways: (1) converting xenobiotics that escaped the transporter-based barrier into substrates for efflux transporters including Mrp1, 2, and 4 and (2) converting waste products of CNS metabolism into substrates that are more easily eliminated from the brain and subsequently pumped into the blood.
In the present study, we show that the nuclear receptor, FXR, is expressed in isolated rat brain capillaries. Consistent with this, CDCA, a high-affinity FXR ligand (Gnerre et al, 2004), increased both Mrp2 protein expression in plasma membranes of isolated brain capillaries and specific luminal Texas Red fluorescence in capillary lumens. Recently, a regulatory network of nuclear receptors, metabolizing enzymes, and efflux transporters was described in hepatocytes (Rosenfeld et al, 2003). In this network, nuclear receptors, such as PXR, FXR, and CAR, sense xenobiotics, for example, drugs, dietary constituents, and environmental pollutants, and coordinately activate the transcription of multiple target genes. These include phase-I and phase-II metabolizing enzymes as well as phase-III efflux transporters. Thus, in liver, nuclear receptors act as ‘xenobiotic sensors,’ orchestrating a coordinated response to potentially harmful xenobiotics. At present, it is not clear to what extent such regulatory networks coordinate transport and metabolism at the blood—brain barrier. In addition to PXR and FXR, we now have preliminary evidence for mRNA expression of other ligand-activated nuclear receptors in brain capillaries, including CAR, liver X receptor, vitamin D receptor, and glucocorticoid receptor (B. Bauer et al, unpublished data). Therefore, it is possible that these receptors participate in an analogous, complex regulatory network at the blood—brain barrier. Receptor activation may lead to the upregulation of efflux transporters and metabolizing enzymes, such as P-glycoprotein, Mrp2, and GSTπ (present study; Bauer et al, 2004). Increased metabolism and efflux transport could then contribute to tightening of the barrier and increased protection on the one hand, but impaired CNS pharmacotherapy on the other. For example, PXR and FXR activation by endogenous ligands could explain increased Mrp2 expression in focal brain regions of epileptic patients (Loscher and Potschka, 2005a). An understanding of this regulatory network at the blood—brain barrier, including mechanisms modulating drug metabolism and efflux, could lead to novel strategies to improve pharmacotherapy of numerous CNS disorders, such as epilepsy, Alzheimer's disease, or brain cancer.
Footnotes
Acknowledgements
We thank Dr Daniel Bow and Dr Kim Brouwer (School of Pharmacy, University of North Carolina, NC, USA) for providing TR− rats and Dr Rosalinde Masereeuw for providing the Mrp4 primer sequences.
None.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.