
Abstract
Striatal dopamine (DA) release has been shown during behavioural tasks, but the relative contribution of motor, reward, and cognitive components is unclear. Dopamine release was quantified using [11C]-raclopride in two studies using a triple-scan approach, comprising active task, motor control, and rest. In the first, bolus radiotracer was delivered during a sequential motor learning paradigm; in the second, a spatial planning task, bolus plus constant infusion was applied. [11C]-raclopride binding potentials (BPNDs) in striatal functional subdivisions were compared across conditions. [11C]-raclopride BPND was significantly reduced in active task compared with rest in both the sensorimotor and associative striatum in both studies, because of differences between rest and motor control conditions. In both regions, the motor control BPND fell between the rest and active task in the planning study, but the difference between motor control and active task conditions was not significant. No such changes were observed in the limbic striatum. Using rigorous methodology, this study validates earlier evidence that striatal DA release occurs during behavioural challenges. Increased DA release during movement was reliably detected in the sensorimotor and associative striatum, supporting use of the functional subdivision model in humans. No additional DA release was observed specific to the cognitive component of either task.
Introduction
It is well established that radiotracer [11C]-raclopride ([11C]RAC) is sensitive to pharmacological challenges of the dopamine (DA) system (Laruelle, 2000). More controversial is the question whether behavioural challenges might induce sufficient changes of DA to be measured with [11C]RAC. Ten years ago, increases in striatal DA release were detected in humans during performance of a video game, through use of the D2/3 DA receptor radiotracer [11C]RAC and positron emission tomography (PET) imaging (Koepp et al, 1998). Using this technique, many studies have since reported increases in DA release during performance of behavioural tasks (reviewed in A Egerton
Performance of cognitive tasks generally involves several component processes, such as motor output, attention, and executive functions including reward and planning. Dopamine release during the task is often compared with an ‘at rest’ baseline measurement; it is therefore unclear whether increases in DA hypothesised to occur, for example, during a specific cognitive process are separable using [11C]RAC from increases in DA arising from inherent additional components of the task.
Motor aspects of task performance have a recognised role in DA release; DA has been widely implicated in the sequencing and control of motor and visuomotor coordination (Graybiel, 1995). Dopamine release during motor sequence execution in humans is supported by evidence in primates that activation of DA neurons occurs during repetitive movement sequences (Magarinos-Ascone et al, 1992). Human striatal DA release has been shown in several [11C]RAC PET studies during performance of motor tasks, including foot extension/flexion (Ouchi et al, 2002); sequential finger movement (Goerendt et al, 2003); and finger opposition (Badgaiyan et al, 2007).
Importantly, increases in striatal DA may also be observed using [11C]RAC PET when motor output is not required. For example, increases in DA release are reported after placebo administration under several conditions (Fuente-Fernandez et al, 2001; Strafella et al, 2006); and in reward tasks (Schultz, 1998). Task-induced DA release may result from any of a range of cognitive processes, such as learning, memory, attention, movement; indeed, striatal DA is regarded as central to the integration of cognitive and motor processes (Frank, 2005). Use of appropriate sensorimotor control tasks is therefore vital to dissect out the specific cognitive component of interest, to show that increases in DA are specific to an individual task component. Recent evidence suggests this may be possible: increases in striatal DA release have been reported during a set-shifting executive task compared with a nonset-shift control task where task stimuli and required motor output were equivalent across conditions (Monchi et al, 2006a).
Anatomic projections to and from the striatum are organised as discrete cortico—striato—thalamic networks in which different types of information are processed (Middleton and Strick, 2000). Haber et al (2000) examined these connections in macaques and concluded that the ventral midbrain provides an interface for information flow of motor, cognitive, and limbic processing between different striatal regions. In an extension of this organisational description, Martinez et al (2003) studied the human striatum according to Haber and colleagues’ classification into its ‘functional subdivisions’; and proposed the sensorimotor striatum (SMS; comprising dorsolateral putamen) to be implicated in motor output; the associative striatum (AST; caudate and ventrolateral putamen) in cognition; and the limbic striatum (LS; ventral) in drive, motivation, and reward (Martinez et al, 2003). Thus, task-induced changes in DA may be related to regional functional specialisation. Reviewing the location of reported effects in previous [11C]RAC imaging studies generally supports such a functional dissociation (see A Egerton
This study aimed to investigate striatal DA release during two tasks, to dissect out the motor and cognitive components intrinsic to the tasks. In the first, sequential motor learning was compared with a motor control condition and to a rest condition. In the second paradigm, spatial planning was compared with motor control and rest conditions in a larger sample and applying the more recently developed technique of bolus infusion for radiotracer delivery. In both paradigms, a triple-scan approach (active task, motor control, and rest) was adopted to control for the effect of motor output, and thus to allow dissociation of movement from either sequential learning or spatial planning.
A motor sequence learning task was selected for the first experiment because the role of the striatum in motor sequence learning is well recognised: in nonhuman primates, dopaminergic lesions of the striatum impair learning of motor sequences (Matsumoto et al, 1999), and striatal neurons are activated during procedural learning (Miyachi et al, 2002). Evidence for a role in humans comes from a recent study, which showed increased [11C]RAC displacement in the anterior body of the caudate nucleus and dorsomedial putamen during sequential motor learning compared with a motor control condition (Badgaiyan et al, 2007).
For the second experiment, the Tower of London (TOL) was the spatial planning task selected because it is a motor task, which can be administered at levels of difficulty, which require planning or no planning, both with an absence of any explicit reward component. The effects of spatial planning on DA release have not previously been investigated using [11C]RAC PET, although several studies emphasise a link between striatal DA markers and planning. For example, spatial planning has been associated with D2 receptor availability in the striatum of patients with Huntingdon's disease (Lawrence et al, 1998) and presynaptic DA availability in Parkinson's disease (Cheesman et al, 2005). Previous work by this group also showed that spatial planning accuracy, measured using the TOL task, is positively correlated with D2 receptor availability in the caudate and putamen of healthy postmenopausal women (Reeves et al, 2005). This relationship is particularly apparent when solving difficult problems, and motor output may also contribute. Further, percentage increases in [11C]RAC binding potential (BPND) after DA depletion positively correlate with worsening of TOL performance (Mehta et al, 2005). On the basis of these previous results, it was predicted that increases in DA release would occur during the spatial planning (TOL) task. Administration of [11C]RAC by bolus plus constant infusion negated potential blood flow influences on findings (Carson et al, 1997).
Aims
This paper describes two experiments conducted with common aims: to investigate the relative contribution to DA release of the motor and cognitive elements of a behavioural task through use of a control condition, and to examine these effects across the functional subdivisions of the striatum.
Based on the findings described above, the following hypotheses were formulated:
Dopamine release in the SMS would relate to the motor components of the task.
Dopamine release in the AST would relate to the cognitive components of the task.
Dopamine release in the LS would not significantly differ across conditions.
Dopamine release in the motor control condition would be intermediate between that observed in the active task and rest conditions.
Methods
Sample
Experiment 1
Six healthy, right-handed men (age range 35 to 45 years) were recruited by public advertisement. Screening involved a full medical and psychiatric history. Exclusion criteria were as follows: age <35 years, presence of current or previous psychiatric or neurological illness, including drug or substance abuse; history of any cerebrovascular accident; use of any medications, which may have affected DA activity. All participants provided written informed consent before inclusion. The study was approved by the Research Ethics Committees for the Hammersmith Hospital.
Experiment 2
Ten healthy right-handed men (age range 35 to 70 years) were recruited from a volunteer database collated from consenting volunteers to previous studies run at the Institute of Psychiatry, KCL. Screening was as per Experiment 1. The study was approved by the Research Ethics Committees for the Hammersmith Hospital and the South London and Maudsley NHS Trust.
In both cases, the Administration of Radioactive Substances Advisory Committee granted permission to administer [11C]RAC.
Design
Experiment 1 Sequential Motor Learning
The active task condition involved learning new sequences of key presses. Each sequence was eight moves long and was learned by trial and error. Movements were paced by a tone, which sounded every 2500 ms (participants wore headphones). After each pacing tone, participants pressed one of the keys, in an attempt to identify the order of the sequence. If the correct key was pressed, a high-pitched tone immediately sounded to give positive feedback. An incorrect movement was followed by a low-pitched tone. By pressing keys and receiving feedback, participants would gradually be able to learn the correct eight-move sequence, indicated by a burst of short high-pitched tones. The process was repeated during the duration of the scan. During the motor control (overlearned fixed sequence) condition participants were asked to repeatedly produce the same eight-move fixed sequence, which involved touching the keys in ascending numerical order. Movements were similarly paced—every 2500 ms—to match the movement performed during the learning paradigm.
Experiment 2 Spatial Planning
An adapted version of the one-touch Tower of London test of spatial planning was used (Owen et al, 1995). Participants viewed a computer touchscreen, on which were displayed two arrangements of coloured balls in pockets, one in the upper and one in the lower half of the screen. From left-to-right the pockets could hold three, two, or one of the balls, respectively; and each arrangement consisted of a red, green, and blue ball. At the start of each new trial, participants were requested to consider the minimum number of moves needed to rearrange the balls on the bottom half of the screen in order that they match the arrangement of the balls on the top half. They provided their solution by touching the relevant number from a selection of coloured boxes labelled 1 to 6 at the bottom of the touch-sensitive screen. Participants completed five blocks of the task, each comprising a total of 16 trials. Two trials in each block required participants to show the solution to the problem: after having selected the coloured box with their numerical solution, the command ‘Show me’ appeared across the touchscreen. Participants then touched the balls to select them, and touched the screen again to indicate their destination, until the task was completed. The final position of the balls was varied such that the minimum number of moves to solution was predominantly three, four, or five moves. The motor control task replicated all aspects of the experimental condition but replaced ‘difficult’ (three, four, and five moves) trials with ‘easy’ (one or two moves) problems, which can be solved by visual matching alone. As in Experiment 1, the motor control condition was matched to the active task condition in terms of type, pattern, and number of movements; and additionally in the TOL, for nonspecific task requirements such as visual activation.
Positron Emission Tomography Scanning
Procedure
Participants were asked to abstain from caffeine for 12 h before scanning. Volunteers were each scanned on three occasions, at least 1 week apart. The order of scan conditions was randomised for all participants. During the rest scan, participants lay at rest in the quietened scanner suite. In all cases head movement was minimised by use of a moulded head rest and straps.
Radiotracer Delivery
Experiment 1
[11C]RAC was administered as an intravenous bolus injection of 120 MBq, given over 20 secs and scans were acquired over 60 mins. Scans were acquired on a ECAT EXACT HR++ PET tomograph (CTI/Siemens 966, Knoxville, TN, USA).
Experiment 2
[11C]RAC was administered as a bolus injection followed by a constant rate infusion with a κbol=70 mins (Watabe et al, 2000). The bolus-infusion method enables a state of equilibrium to be reached, which avoids potential artefacts introduced through blood flow change during the scan (Carson et al, 1997). Scans were acquired on a Siemens ECAT 962 PET camera (CTI) in three-dimensional mode over 70 mins. Total administered activity was 370 MBq per scan. An increase in DA release is inferred in such a paradigm when a decrease in the [11C]RAC BPND is observed; after competition between [11C]RAC and endogenous DA at D2/3 receptors (Laruelle, 2000). The sampling period for calculation of BPND, once [11C]RAC equilibrium had been established, was 35 to 70 mins from a 70 mins dynamic scan collected over 22 serial time frames.
Image Analysis
[11C]RAC kinetic modelling was performed using a simplified tissue reference model; cerebellar activity was the reference input function. Simplified tissue reference model allows estimation of radiotracer BPND and the relative rate of radiotracer delivery normalised to the cerebellum (R1). Head movement was corrected using a frame-by-frame realignment procedure previously described (Montgomery et al, 2006). Nonattenuated corrected images were used for realignment to provide additional information by reducing the influence of redistribution of radiotracer producing erroneous realignments (Dagher et al, 1998). The nonattenuated corrected image was denoised using a level 2, order 64 Battle Lemarie wavelet filter (Turkheimer et al, 1999). A mutual information algorithm (Studholme et al, 1997) was applied for frame realignment to a single frame acquired 35 mins postinjection, in which there was a high signal-to-noise ratio. Transformation parameters were applied to the corresponding attenuation-corrected dynamic images (using frame 15 as a reference frame), to generate a frame-by-frame corrected dynamic image.
Region of Interest Analysis
Region of interest templates for striatal and cerebellar areas were defined on a magnetic resonance scan positioned in standard Montreal Neurologic Institute space (Hammers et al, 2003) and then normalised into individual native space for each scan using a RAC template. Three striatal subregions were examined bilaterally, based on previously described criteria (Martinez et al, 2003): LS; AST; and SMS. The ROI templates described above were then placed on parametric images of BPND (and R1 in the case of Experiment 1), to obtain regional values of the parameters of interest. The BPND was calculated as the ratio of specifically bound (striatum) radioligand to that of nonspecifically bound (cerebellum). Both cerebellar and striatal regions were sampled from the whole dynamic scan in Experiment 1; and from time frames 15 to 22 in Experiment 2.
Voxelwise Analysis
Voxel-based analyses were used to confirm the findings of the ROI analysis in Experiment 2 only, as the few participants in Experiment 1 precluded meaningful application of a voxel-based approach. Statistical parametric mapping 5 was used to perform voxelwise comparisons across the three test conditions to detect any localised striatal changes not detected using ROI. Parametric BPND images were created using dynamic images of mean activity in frames 15 to 22 (35 to 70 mins); activity at each voxel was divided by the cerebellar reference region activity for that image; and 1 subtracted to give BPND at each voxel. Binding potential images were normalised into standard Montreal Neurological Institute space with a RAC template, coregistering with the full dynamic image (1 to 22 frames). Smoothing was performed using an isotropic Gaussian kernel (full width half maximum=8 mm). Analyses were restricted to BPND values>1.0, so as to limit analysis to the striatum only. A full factorial model was applied with a height threshold of
Statistical Analysis
Statistical analyses were performed using the Statistical Package for Social Sciences, version 12.0 (SPSS Inc., Chicago, IL, USA). Nonparametric correlations were used as appropriate for ordinal or nonnormally distributed data. A repeated-measures approach was used to establish whether the main effect of ‘condition’ was significant, over the three scans, in each of the subregions (sensorimotor, associative, limbic).
Experiment 1
Friedman's (nonparametric equivalent of repeated-measures design) and Wilcoxon's signed-rank (nonparametric equivalent of paired
Experiment 2
Repeated-measures analysis of variance were applied to investigate the main effect of condition and the interaction of condition × side; linear contrasts were derived from within-subjects tests. Paired
Results
Sample Characteristics
Experiment 1
Mean age: 39 years (s.d.=4; range=35 to 45); all right handed. During motor learning, mean number of new sequences learned: 16 (s.d.=5; range=10 to 24); mean number of errors: 415 (s.d.=28; range=390 to 462). Age was not correlated with new sequences learned (
Experiment 2
Mean age of the sample was 53 years (s.d.=9; range=39 to 68). All were right handed. Current pharmacotherapy applied for only one subject (10%) who was taking a lipid-lowering drug. Tower of London accuracy was high both offline (mean=89.5%; s.d.=10; range=75 to 100) and during the scan (mean=89.0%; s.d.=4; range=81 to 95). In scanner planning accuracy did not correlate with age (
Region of Interest
Experiment 1
Mean BPND values for SMS, AST, and LS at rest and during the motor control and motor learning (active task) conditions are shown in Table 1. The Friedman's test detected a significant effect of condition in the SMS and AST. Wilcoxon's signed-rank test was used to estimate, in these regions as a whole and by side, the relative significance of the change between paired conditions. Binding potential was significantly decrease during sequential motor learning compared with rest in the SMS as a whole (
Mean BPND across task conditions in the sequential motor learning task (Experiment 1)
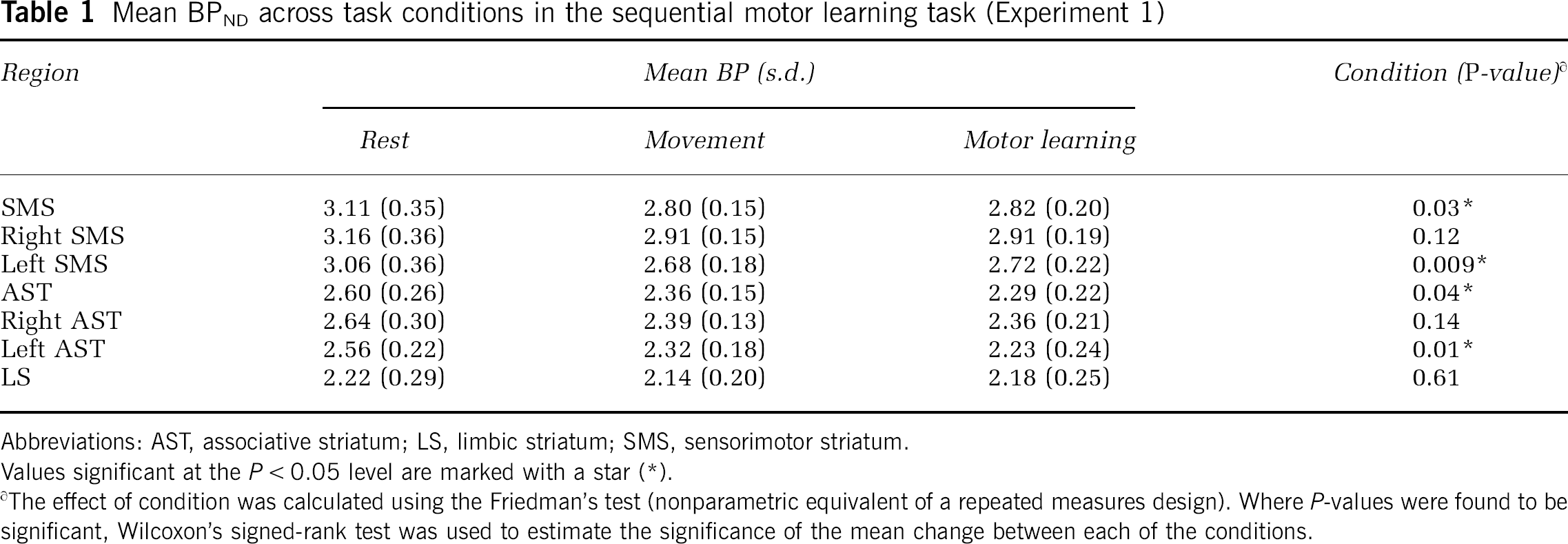
Abbreviations: AST, associative striatum; LS, limbic striatum; SMS, sensorimotor striatum.
Values significant at the
∂The effect of condition was calculated using the Friedman's test (nonparametric equivalent of a repeated measures design). Where
No significant differences were detected in [11C]RAC BPND in the sequential motor learning compared with the motor control condition in any region. There was no significant difference in R1 across conditions (Friedman's test;
Experiment 2
Mean BPND at rest and during the motor control and spatial planning (active task) conditions are shown in Figure 1 and Table 2. Repeated-measures analysis of variance analyses conducted for each of the three subregions (SMS, AST, LS) are also shown in Table 2, detailing the main effect of condition and the interaction of side × condition. Linear contrasts were significant for both SMS (F(2,18)=14.15;
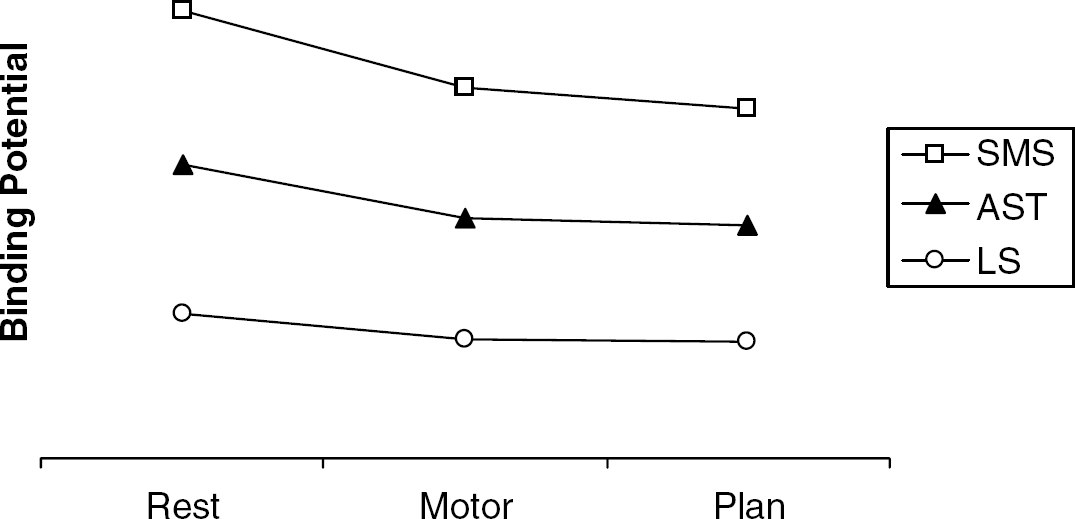
Mean raclopride binding potential (BPND) values for sensorimotor, associative, and limbic striatum across the three test conditions of the Tower of London task. Linear contrasts were significant for sensorimotor (F(2,18)=14.15;
Repeated measures analyses by region comparing mean BPND values during the rest, motor control, and planning (active task) conditions of the Tower of London task (Experiment 2)
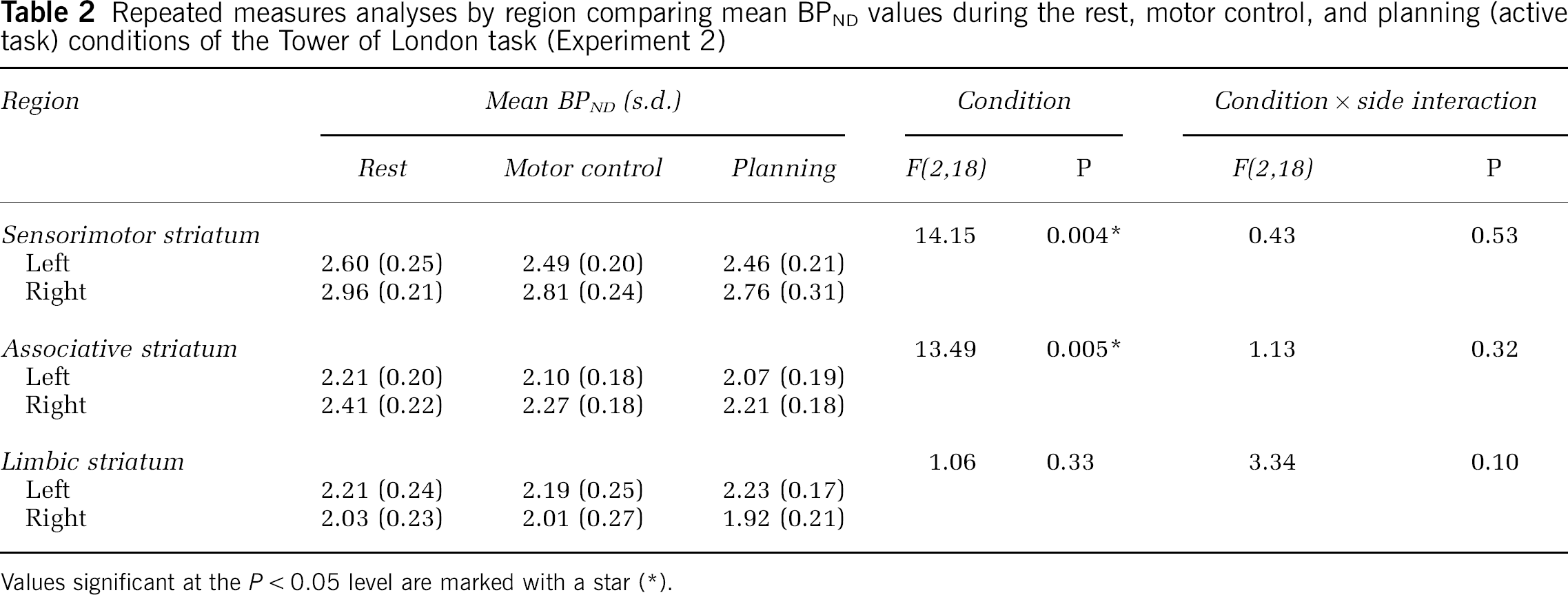
Values significant at the
There was no significant interaction of condition and side in any region.
Paired
Paired
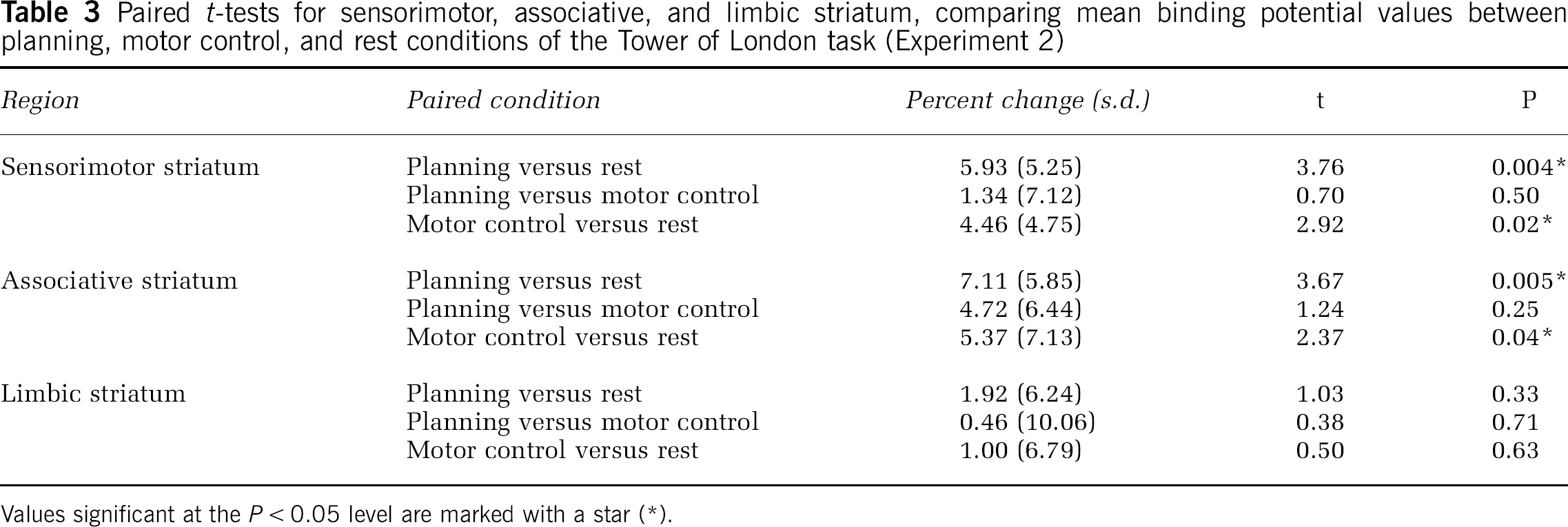
Values significant at the
Voxelwise Analyses
The planning condition compared with the rest condition generated four significant clusters of reduced [11C]RAC BPND (Figure 2; Table 4). A large right-sided cluster was located in the AST; of the three left-sided clusters, two were located in the SMS, and one in the AST. In the reverse condition—comparing the rest condition to planning—there were no significant clusters. Motor control versus rest analysis revealed one right-sided cluster, which was smaller in extent, and which had its peak in the AST with some extension into the SMS (Figure 3; Table 4). In the reverse condition—comparing the rest condition to motor control—there were no significant clusters. There were no significant clusters in either direction in the planning versus motor control analysis.
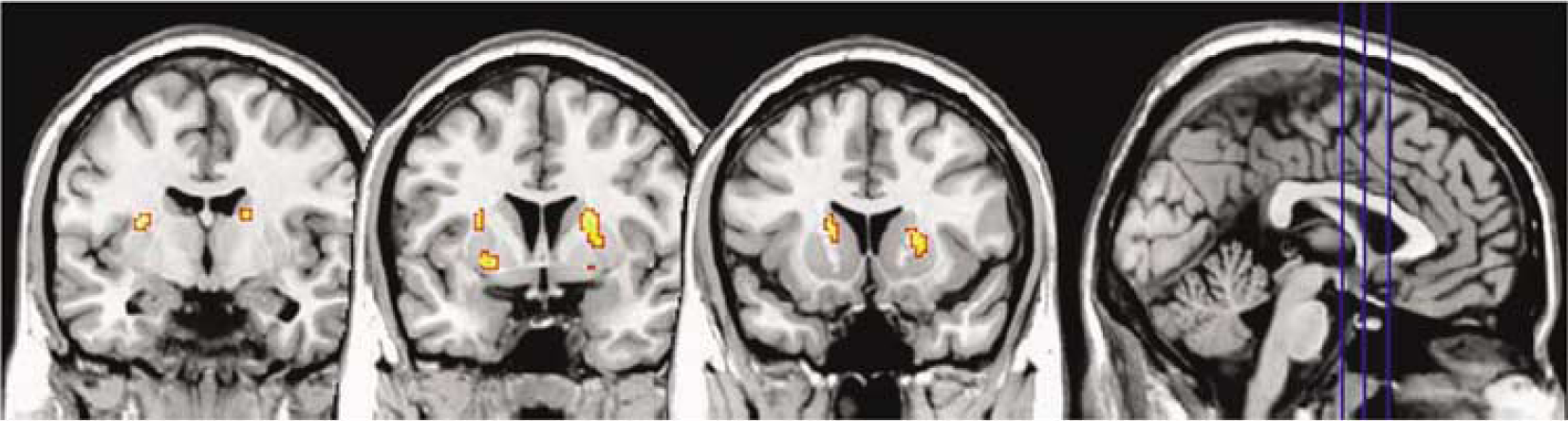
Coronal sections from the statistical parametric map of BPND change in the planning versus rest condition overlaid on a canonical image in stereotaxic space. Right side image corresponds to right side brain. The leftmost section (postcommisural;
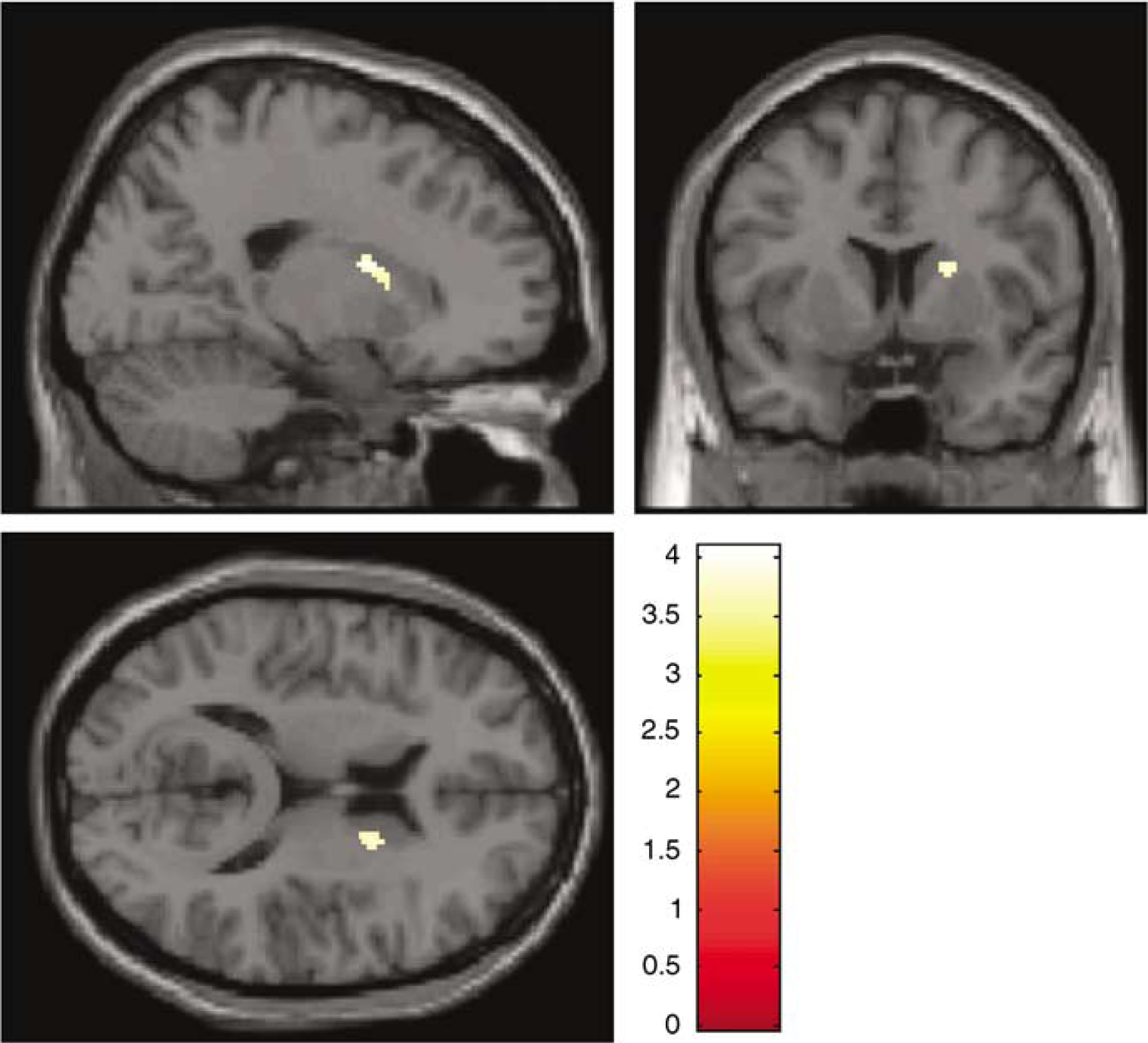
Sagittal, coronal, and axial sections of the statistical parametric map of BPND change overlaid on a canonical image in stereotaxic space. The displayed cluster shows the significant area of decreased raclopride binding potential in the right SMS and AST in the motor control compared with rest comparison ((
SPM analysis showing all clusters yielded at voxelwise significance level 0.001 (Experiment 2)
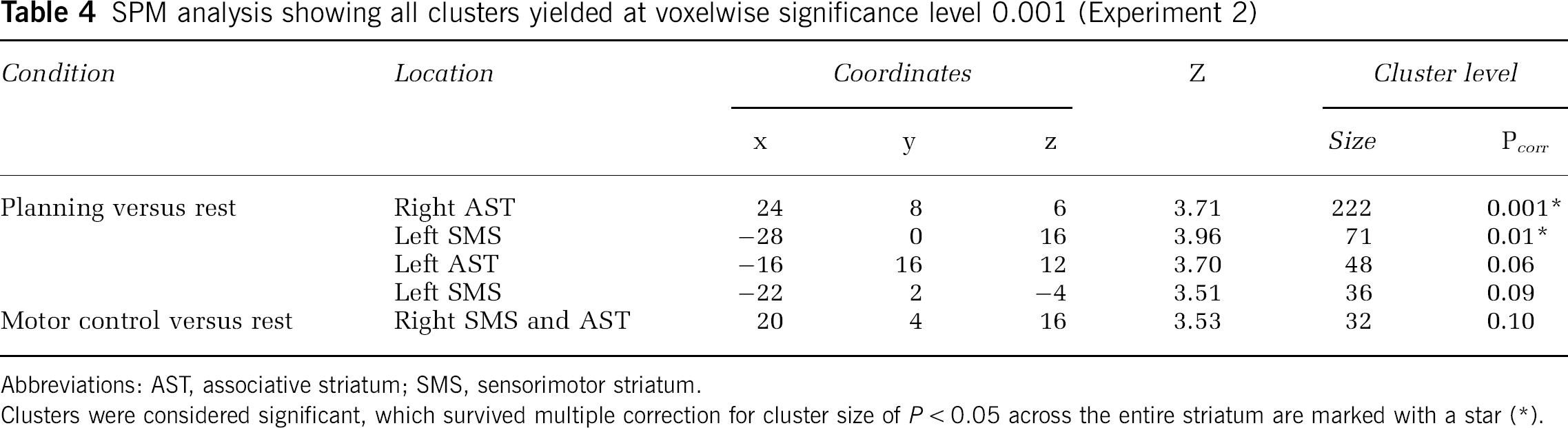
Abbreviations: AST, associative striatum; SMS, sensorimotor striatum.
Clusters were considered significant, which survived multiple correction for cluster size of
Discussion
This study replicates previous work by our group showing that DA release during behavioural tasks can be detected using PET, and validates the finding using state of the art techniques. Using robust methodology, this study has showed that significant increases in striatal DA levels occurred during two behavioural tasks. During both sequential motor learning and spatial planning paradigms, DA levels increased in the SMS in both the active task and motor control conditions relative to rest. The
Dopamine release occurred in the SMS during all conditions with a movement component across the sequential motor learning and spatial planning paradigms. Striatal DA release during motor sequence learning has been investigated only once previously using [11C]RAC PET administered by bolus plus constant infusion (Garraux et al, 2007). Compared with an oculomotor control task, finger sequence learning produced significant decreases in striatal [11C]RAC BPND. However, no motor output was required during the control task and the effects of learning could not be quantified because of the absence of a rest scan.
As in the study presented here, changes in [11C]RAC BPND during sequential motor output have been reported relative to a baseline condition where no equivalent motor output was required (Goerendt et al, 2003; Garraux et al, 2007). However, whereas increased displacement of [11C]RAC during motor sequence learning has been reported relative to a motor control condition (Badgaiyan et al, 2007), there was no significant difference in [11C]RAC BPND between motor control and motor sequence learning tasks in this study. There are several possible explanations for this discrepancy in findings: first, the motor sequence learning and motor control tasks differed between studies; it may be that the learning task used by Badgaiyan et al (2007) was more effective in stimulating DA release relative to control. Second, the small sample size (
In summary, DA release elicited by sequential motor learning was not clearly dissociable from that elicited by motor components. This finding was replicated in the spatial planning study, which was conducted in a larger sample and using more recently developed PET techniques (bolus plus constant infusion). Again, significant DA release was showed in the SMS in the motor control condition compared with rest, with no significant additional DA release because of the planning component alone when movement inherent to the task was subtracted out. These findings uphold the first hypothesis of the study, namely: DA release in the SMS was because of the motor components of the task.
The second hypothesis predicted that DA release in the AST would relate to the cognitive components of the task; this was not upheld in either Experiment 1 or Experiment 2. Thus, in both the sequential learning and spatial planning paradigms, whereas there was a significant effect of condition on DA release in AST (Tables 1 and 2),
The AST finding from Experiment 2 is superficially consistent with previous research that showed increased DA release occurring in the AST during planning compared with rest (Monchi et al, 2006b). During that paradigm, planning a set-shift gave rise to increased DA release in both the caudate nucleus and anterior putamen, which correspond functionally to the AST. However, in this study there was no evidence that DA release occurred because of planning alone, when separated from the inherent movement component of the task. The finding is therefore at odds with that of Monchi et al (2006a) who showed that increases in striatal DA release occurred during a set-shifting executive task compared with a nonset-shift control task in which task stimuli and movement were equivalent (i.e., movement was controlled). Although methodological factors may underlie the disparity between the results of Monchi
With respect to the TOL task, numerous studies attest to its utility and sensitivity to variations in dopaminergic function. Superior performance on the TOL necessitates successful planning, execution, monitoring, and revision of a sequence of actions using working memory (Owen, 1997) and there is significant evidence for involvement of striatal DA in planning performance: there is impairment in patients with mild Parkinson's disease (Owen et al, 1995), and modulation of performance (Lange et al, 1992) occurs after withdrawal from dopaminergic medication. Furthermore, there is evidence that TOL performance is associated with increased 18F-fluorodopa uptake in the caudate (AST) in participants with Parkinson's disease (Cheesman et al, 2005). Despite this body of evidence it remains possible that the control and task conditions were not adequately distinct in terms of planning requirements. Counter to this explanation are two findings showing increased blood flow in the caudate with increasing TOL task difficulty (Dagher et al, 1999) and work from this group showing striatal [11C]RAC BPND correlates with planning accuracy, but only for difficult problems and not easier problems (Reeves et al, 2005). In addition, PET and functional magnetic resonance imaging studies comparing easy with difficult problems have revealed clear differences in brain activation patterns (Dagher et al, 1999).
The observation that significant reductions in [11C]RAC BPND occurred in both the SMS and in the AST in motor control condition compared with rest indicates that movement is not specific to the SMS functional subdivision in humans. This finding is in keeping with the feed-forward model proposed by Haber et al (2000), which describes information flow from the AST to the motor striatum, and integration of signals across these and other nodes in the network. The findings reported here support this model, rather than specificity of motor activity to the SMS alone.
The third hypothesis, that DA release in the LS would not significantly differ across conditions, was upheld in both experiments. In the functional subdivision model, DA release in the LS is thought to particularly arise in response to emotional or rewarding stimuli. The tasks used here did not involve any explicit reward, and therefore no increased DA release in the LS was expected.
Lastly, the linear trend analysis used all data available and showed that significant negative slopes existed across active task-motor control-rest conditions in the spatial planning study, providing some support for the final hypothesis. However, the more direct paired comparisons suggested no change occurred between active task and motor control conditions.
Limitations
A limitation of the study is the size of the mean reduction in BPND values observed across the conditions. The percentage change values derived in the SMS across planning and rest conditions (5.93%) and across motor control and rest conditions (4.46%; Table 3) are in the order of the 5% changes observed with test—retest reliability analyses. The high standard deviation values for these changes are to be expected given the small regions in which an effect is being sought; yet they prompt a need for some caution in interpretation. Thus, it cannot be ruled out that these findings may have occurred by chance. However, through the inclusion of 10 participants, the spatial planning study was powered (0.8) to detect changes of 5% between two conditions. Furthermore, given that the change in DA release was in the hypothesised direction (i.e., planning>movement>rest), more confidence can be invested in these being true findings. To test this, analyses were rerun comparing BPND across regions by original (random) order of scan: that is, Scan1 versus Scan2 versus Scan3. For all regions there were no significant changes by scan number (analyses not shown). This adds weight to the proposition that the findings reported here represent true differences in DA release.
A related issue is that of the multiple comparisons conducted during statistical analysis; correction for multiple comparisons was not performed here, but all analyses were hypothesis led.
Conclusions
This study provides evidence of demonstrable DA release occurring in the human striatum during two behavioural tasks, which was driven primarily by the motor component of each task. These results validate the findings of studies that were performed (e.g., (Koepp et al, 1998; Larisch et al, 1999; Goerendt et al, 2003) before the introduction of techniques such as head movement correction or bolus infusion. Further, the incorporation of the striatal functional subdivisions into the design of this study enabled the identification of the SMS as an area integral to movement, but also showed a role for the AST in movement. These findings are in keeping with the feed-forward striatal functional subdivision model, and support future application of the model in humans. Finally, applying state of the art methods, additional DA release specific to the cognitive component of the planning task was only detected in a regression analysis utilising the resting BPND measurements, but not when comparing the cognitive and motor conditions directly. This pattern of findings raises the question whether DA release during nonrewarding cognition can reliably be imaged using current PET techniques.
Footnotes
Acknowledgements
Many thanks to all participants, and to the radiographers at the Hammersmith site for their valuable help in completing this study.
Dr Julia M Lappin, Dr Suzanne J Reeves, and Dr Mark Coulson report no competing interests. Dr Alice Egerton is funded by GlaxoSmithKline through Imperial College and reports no competing interests. Dr Mitul A Mehta reports research support from Pfizer, GlaxoSmithKline, and Evotec and has acted as a consultant for GlaxoSmithKline and Evotec. MAM reports no conflicts of interest. Professor Paul M Grasby has served as an occasional consultant to GlaxoSmithKline, Merck, and Pfizer and reports no competing interests.