
Abstract
Current thrombolytic therapy for acute ischemic stroke with tissue-type plasminogen activator (tPA) has clear global benefits. Nevertheless, evidences argue that in addition to its prohemorrhagic effect, tPA might enhance excitotoxic necrosis. In the brain parenchyma, tPA, by binding to and then cleaving the amino-terminal domain (ATD) of the NR1 subunit of N-methyl-
Introduction
Tissue-type plasminogen activator (tPA) remains the only approved therapy for acute ischemic stroke. Initially described as a component of the fibrinolytic system, a number of studies have later shown a role of this serine protease in synaptic plasticity-related processes and neurodegenerative pathologies, including acute stroke (Benchenane et al., 2004; Melchor and Strickland, 2005; Samson and Medcalf, 2006). It is also established that exogenous tPA can cross both the intact and the injured blood—brain barrier to reach the brain parenchyma (Benchenane et al., 2005a, 2005b; Harada et al., 2005). Thus, both endo- and exogenous tPAs are capable of influencing brain functions and dysfunctions.
The precise function of tPA in the brain is nevertheless surrounded by some uncertainty. One prominent idea is that tPA acts extracellularly to stimulate the elongation of axons and induces synaptic plasticity (Baranes et al., 1998; Neuhoff et al., 1999). In addition, a growing number of observations are showing that tPA can impact neurotransmission in a more direct manner. For instance, tPA/plasmin axis may control the maintenance of the late phase of long-term potentiation through the local activation of proBDNF (brain-derived neurotrophic factor) into mature BDNF (Pang et al., 2004). A number of independent studies have highlighted a role for tPA on the modulation of glutamatergic neurotransmission through a direct action on the function of N-methyl-
Potentiation of NMDAR is well known to play a key role in memory- and learning-related processes (Bliss and Collingridge, 1993). Conversely, overactivation of NMDARs may also lead to excitotoxic neuronal death under certain circumstances (Choi et al., 1987; Sattler and Tymianski, 2000). Although a number of independent reports are a clear evidence of a relationship between tPA and NMDAR signaling, the molecular basis of this interaction needs to be further investigated and elucidated. The present report combines a molecular and cellular approach to determine the molecular background of the interaction of tPA with the NR1 subunit of NMDAR. Moreover, we provide new data to explain how other thrombolytic agents, especially the plasminogen activator derived from the saliva of the vampire bat Desmodus rotundus, desmoteplase (DSPA), in contrast to tPA, might not exert neurotoxic effects.
Materials and methods
Materials
Horse serum and fetal bovine serum were obtained from Life Technologies (Cergy Pontoise, France). Human recombinant tPA was purchased from Boehringer Ingelheim (Paris, France). Antibodies against the NR1 subunit (sc-9058) and 6 × His were purchased from Santa Cruz Biotechnology (Heidelberg, Germany) and Qiagen (Courtaboeuf, France), respectively. The tPA substrate spectrozyme XF-444 was purchased from American Diagnostica (Greenwich, CT, USA). Tenecteplase (TNK) and TNKS478A were kindly provided by Professor G. Bu (Department of Pediatrics, Cell Biology and Physiology, Washington University School of Medicine, St Louis Children's Hospital, St Louis, MO, USA). Reteplase and DSPA were made available by PAION Deutschland GmbH (Aachen, Germany); reteplase was used after the removal of tranexamic acid, an excipient in the marketed preparation.
Cell Culture
Neuronal cortical cultures were prepared from fetal mice (E15 to E16) (Buisson et al., 1998). Dissociated cortical cells were resuspended in Dulbecco's modified Eagle's medium supplemented with 5% fetal bovine serum, 5% horse serum, and 2 mmol/L glutamine and plated in 24-well dishes previously coated with poly-
Excitotoxicity
Slowly triggered excitotoxicity was induced at 37°C by a 24 h exposure to 10 μmol/L NMDA in Dulbecco's modified Eagle's medium supplemented with glycine (10 μmol/L). Neuronal death was assessed by phase-contrast microscopy and quantified by measurement of lactate dehydrogenase release by damaged cells into the bathing medium.
Surface Plasmon Resonance Studies
Binding of tPA, DSPA, TNK, TNKS478A, and reteplase to recombinant ATD-NR1 (rATD-NR1) was studied using surface plasmon resonance (SPR) on Biacore 3000. Recombinant ATD-NR1 was immobilized on the CM biosensor chip using N-hydroxysuccinimide/N-ethyl-N′-(dimethylaminopropyl)carbodiimide chemistry, according to the manufacturer's specifications (Biacore, Uppsala, Sweden). Binding of the test compounds was measured by injecting a defined concentration over the sensor chips that were, or were not, previously covalently absorbed with the wild-type rATD-NR1. All SPR experiments were carried out in triplicate.
Calcium Videomicroscopy Analysis
Primary cultures of cortical neurons were loaded in the presence of HEPES-buffered saline solution containing 5 μmol/L of fura-2/AM plus 0.1% pluronic F-127 (30 mins, at 37°C; Molecular Probes, Leiden, the Netherlands) and incubated for an additional 30 mins in HEPES-buffered saline solution. Experiments were performed at 22°C on the stage of a Nikon Eclipse inverted microscope equipped with a 75 W Xenon lamp and a Nikon × 40, 1.3 numerical aperture epifluorescence oil-immersion objective. Fura-2 (excitation: 340 and 380 nm, emission: 510 nm) ratio images were acquired with a CCD camera (Princeton Instrument, Trenton, NJ, USA), and digitized (256 × 512) using Metafluor 4.11 software (Universal Imaging Corporation, Cherter, PA, USA).
Construction of His-Tagged Recombinant Amino-Terminal Domain/Leucine-Isoleucine-Valine-Binding Protein-Like Domain
The region of the NR1 subunit encoding amino acids 19 to 371 for NR1-1a corresponding to the amino-terminal domain (ATD) was produced in bacterial cultures (Escherichia coli, M15 strain) and purified as described previously (Fernandez-Monreal et al., 2004).
Enzymatic Assay
Thrombolytics were incubated in the presence of a fluorogenic substrate (Spectrozyme XF444; 5 μmol/L) alone or in combination with selected peptides. The reaction was carried out at 25°C in 100 mmol/L HEPES (pH 8.0) containing 150 mmol/L NaCl and 0.01% Tween-80 in a total volume of 100 μL. The amidolytic activity of tPA was measured as the change in fluorescence emission at 440 nm (excitation at 360 nm).
Immunoblotting
Protein samples were resolved on SDS-polyacrylamide gel and transferred onto a polyvinylidene difluoride membrane. Membranes were blocked with 5% dried milk in Tris-buffered saline containing 0.05% Tween-20 and incubated with primary antibodies. After incubation with the corresponding peroxidase-conjugated secondary antibody, proteins were visualized with an enhanced chemiluminescence ECL Plus immunoblotting detection system (Perkin Elmer-NEN, Paris, France).
Molecular Modeling
The 3D model of the ATD-NR1 used in this study has been previously described (Fernandez-Monreal et al., 2004). For the kringle 2 (K2) model, a molecule of the crystal structure (PDB code 1PML) was selected (Padmanabhan et al., 1994). The 3D model of the tPA catalytic domain in active form was modeled on the basis of the microplasmin-staphylokinase-microplasmin complex crystal structure (Parry et al., 1998) by homology modeling. Alignment and model were built and optimized using Model Tool Server and Modeller (Douguet and Labesse, 2001).
To perform docking of K2 onto ATD-NR1, Dock (Cherfils et al., 1991) and FTDock (Katchalski-Katzir et al., 1992) programs were used. The generated K2/ATD-NR1 complexes were handpicked graphically and selected models with lesser energy using the CHARMM program (Brooks et al., 1983) (CHemistry and HARvard Molecular Mechanism) in vacuum. The final complexes were evaluated on the basis of the K2/ATD-NR1 interaction energy.
Peptide Synthesis
Peptide 1 (AQNPSAQA), peptide 2 (AQNPSAQAGGDGD), and peptide 3 (LKNRRL) were synthesized by solid-phase methodology on Fmoc-Ala-HMP, Fmoc-Asp (OtBu)-HMP, and Fmoc-Leu-HMP resins, respectively, using a 433A Applied Biosystems peptide synthesizer (Applera-France, Courtaboeuf, France) and the standard Fmoc procedure as described previously (Leprince et al., 1998). The synthetic peptides were purified by reversed-phase high-performance liquid chromatography (purity was >99.5%) and characterized by MALDI-TOF mass spectrometry on a Voyager DE-PRO spectrometer (Applera-France).
Results
Kringle 2 Domain of tPA is Critical for its Binding to the Amino-Terminal Domain of NR1
We sought to characterize the mechanism by which tPA interacts with the NR1 subunit of NMDAR, the critical step for the potentiation of excitotoxic neuronal death (Fernandez-Monreal et al., 2004; Nicole et al., 2001). Real-time binding reactions were recorded by SPR assays (Hashimoto, 2000), with a His-tagged recombinant form of the ATD of NR1 (amino acids 19 to 371 of NR1; 39,875 Da) termed rATD-NR1 immobilized on a hydrophobic biosensor chip surface performed with increasing concentrations of tPA (1.5 nmol/L to 3 μmol/L, random order of injection). We found a dose-dependent binding of the serine protease to rATD-NR1 (Figure 1, N=3) and the rate constants were determined from the recorded sensorgrams, giving a steady-state affinity (KD) of 213 nmol/L. Studies were then performed according to kinetic models including a one-site model (Langmuir), a two-sites model (parallel reactions), and a two-sites model with docking (linked reactions). Based on statistical analyses including t-test and χ2-values, the SPR data did not fit reliably to a 1:1 Langmuir binding model (χ2=1.79). Therefore, a two-sites model was tested and found to yield a strong prediction of the actual experimental data (KD1=84 pmol/L and KD2=24 nmol/L) (Figure 1). This model can also be promoted by a conformational change based mechanism with KD∗=(kd1/ka1) × (kd2/ka2)=39.8 nmol/L, χ2=0.72.
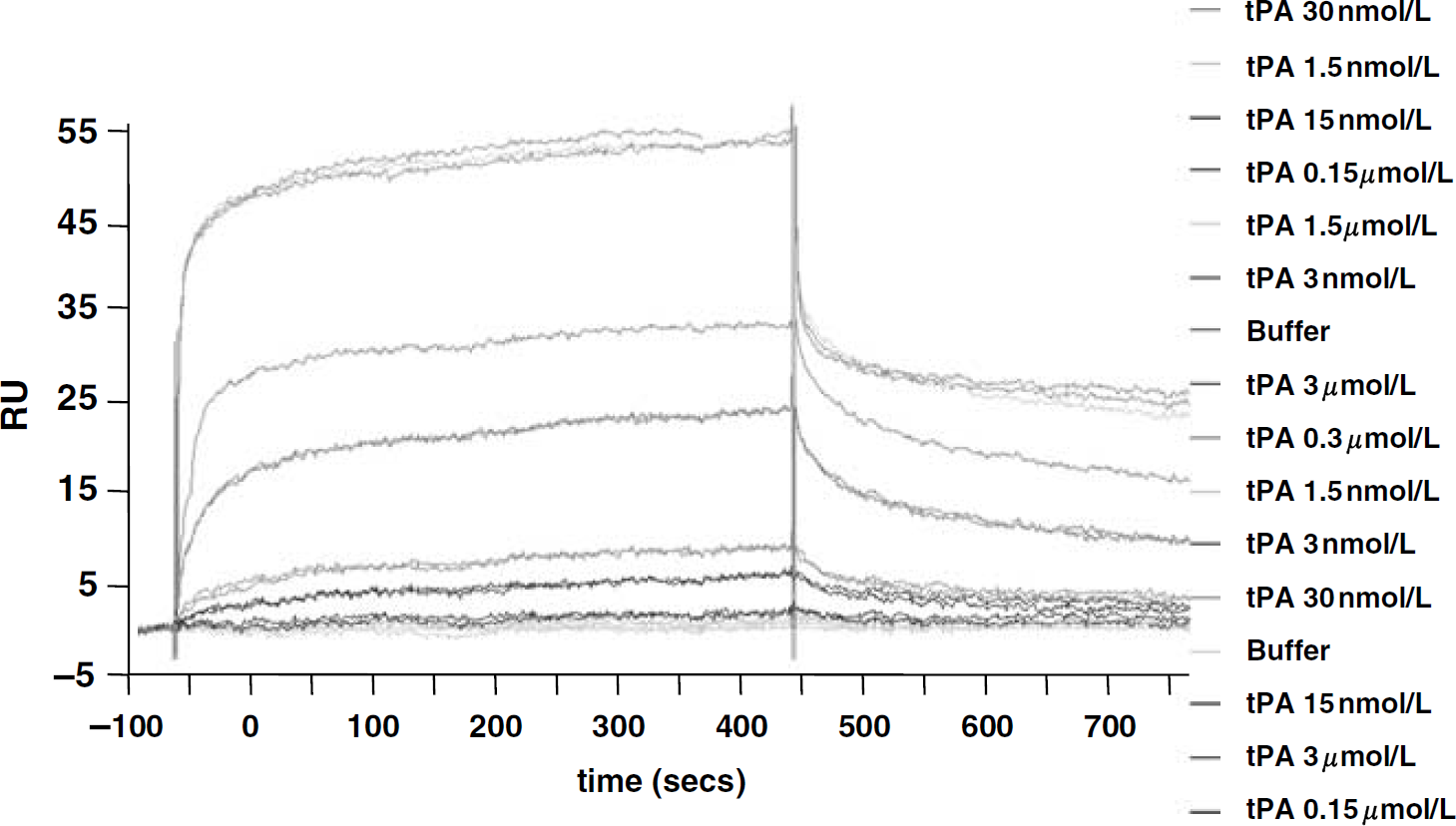
Tissue-type plasminogen activator (tPA) binds NR1 by a two-sites kinetic model. Surface plasmon resonance analysis showing tPA binding to rATD-NR1 in a dose-dependent manner. RU, response units; rATD-NR1, amino-terminal domain of NR1 subunit.
Then, the tPA-related molecules (Supplementary Figure 1), TNK, catalytically inactive TNK (TNKS478A), reteplase, and DSPA, were tested for their ability to bind rATD-NR1 by using SPR as above. Tenecteplase (Figure 2A) is a tPA-derived molecule with specific point mutations in the catalytic triad that confer enhanced fibrin specificity, resistance to plasminogen activator inhibitor, and a prolonged half-life. Wild-type TNK bound to rATD-NR1 with characteristics similar to those of tPA (KD=205 nmol/L) and with a reliable fit of the SPR data to a two-sites kinetics (KD1=138 nmol/L and KD2=43.1 pmol/L; KD∗=3.17 nmol/L; Figure 2A, N=3). In contrast, TNKS478A bound to rATD-NR1 with characteristics closer to a one-site model (Langmuir), suggesting that the catalytic domain of tPA/TNK plays a key role in the two-sites binding to NR1 (Figure 2B). The plasminogen activator derived form the saliva of the vampire bat Desmodus rotundus, DSPA, failed to bind with the rATD-NR1 (Figure 2C). These data suggest that the K2 domain of tPA, which is absent in DSPA, is also critical in mediating binding with the NR1 subunit of NMDAR. Consistent results were obtained with reteplase, a plasminogen activator genetically engineered as the K2 domain and the catalytic domain of tPA (Lapchak, 2002). Reteplase (0.03 μmol/L to 1 μmol/L) bound rATD-NR1 with a pattern similar to that of tPA and TNK, although with a lower apparent affinity (KD=1.64 μmol/L versus 213 and 205 nmol/L for tPA and TNK, respectively; Figure 2D, N=3). These data are in agreement with cleavage assays performed using rATD-NR1 (Figure 3, N=3) or native NR1 (Fernandez-Monreal et al., 2004; Nicole et al., 2001) as a substrate, showing that tPA, TNK, and reteplase can cleave NR1, whereas TNKS478A and DSPA cannot (Figure 3). In consequence, these data suggest sequential binding of the K2 domain (first step) and the catalytic domain of tPA to NR1 (second step).
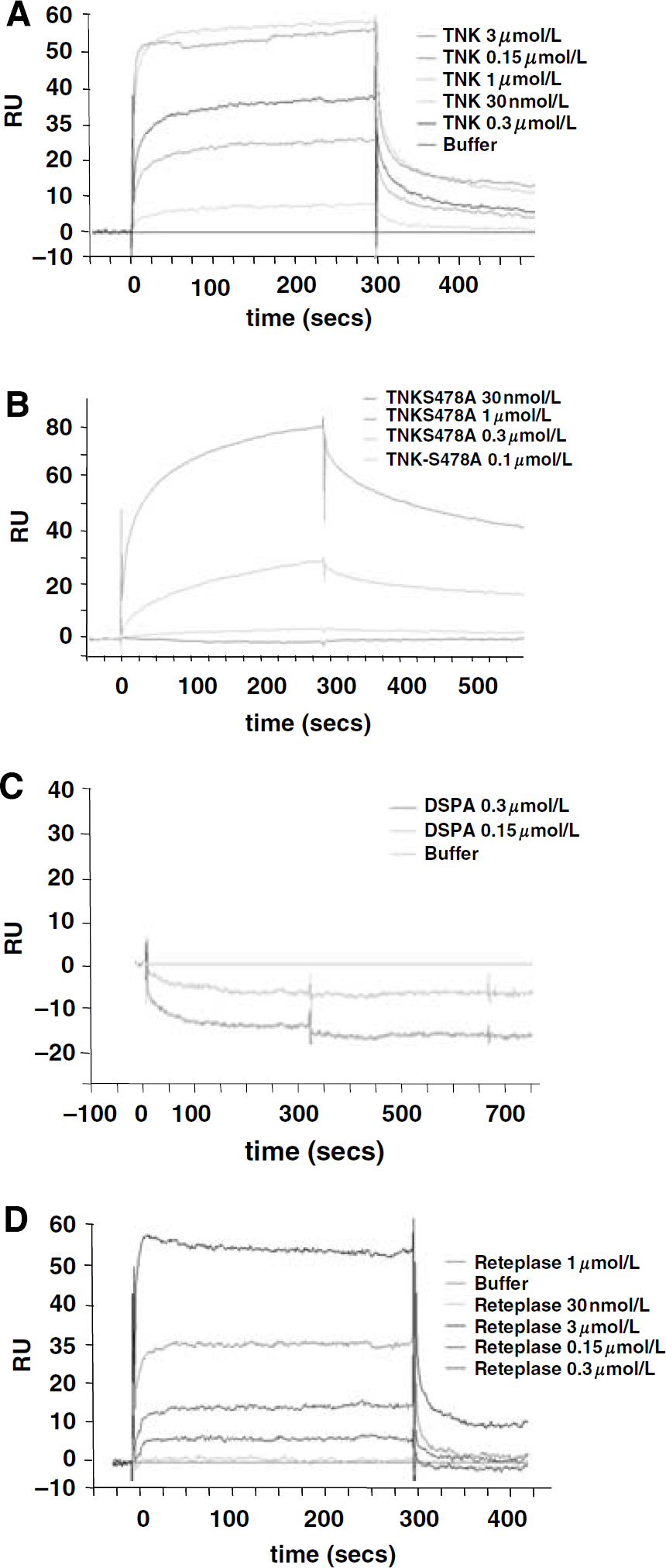
Molecular requirements of tissue-type plasminogen activator binding to ATD-NR1. Surface plasmon resonance response analysis of tenecteplase (TNK) (

Both kringle 2 and catalytic domains of tissue-type plasminogen activator (tPA) are required for cleavage of NR1. Immunoblots performed from rATD-NR1 previously incubated with tPA (0.3 μmol/L), tenecteplase (TNK) (0.3 μmol/L), TNKS478A (0.3 μmol/L), desmoteplase (DSPA) (0.3 and 3 μmol/L), or reteplase (0.3 μmol/L) for 2 h at 37°C. rATD-NR1, amino-terminal domain of NR1 subunit.
Based on the above data, a 3D model for the interaction of both K2 and the catalytic domains of tPA with NR1 was generated (Figure 4A). We propose that tPA forms a ternary complex with two NR1 subunits in a two-step reaction. Firstly, the K2 domain interacts with the hydrophobic pocket of one ATD-NR1 (Figure 4A), and secondly, the catalytic domain binds to R260 (Fernandez-Monreal et al., 2004) of the second ATD-NR1. The data imply a coordinated spatial arrangement of two NR1 subunits, a notion supported by the recent demonstration that native NR1 subunits of NMDARs form dimers under physiologic conditions (Meddows et al., 2001). Furthermore, rATD-NR1 molecules seem to adopt a dimeric conformation that can be identified when subjecting the recombinant protein to SDS-polyacrylamide gel electrophoresis under nonreducing conditions. Compared with reducing conditions, a significant decrease in the 40 kDa band (monomeric rATD-NR1) was observed, accompanied by the appearance of a new band at a higher molecular weight indicative of the generation of dimeric forms of the recombinant proteins (Figure 4B).
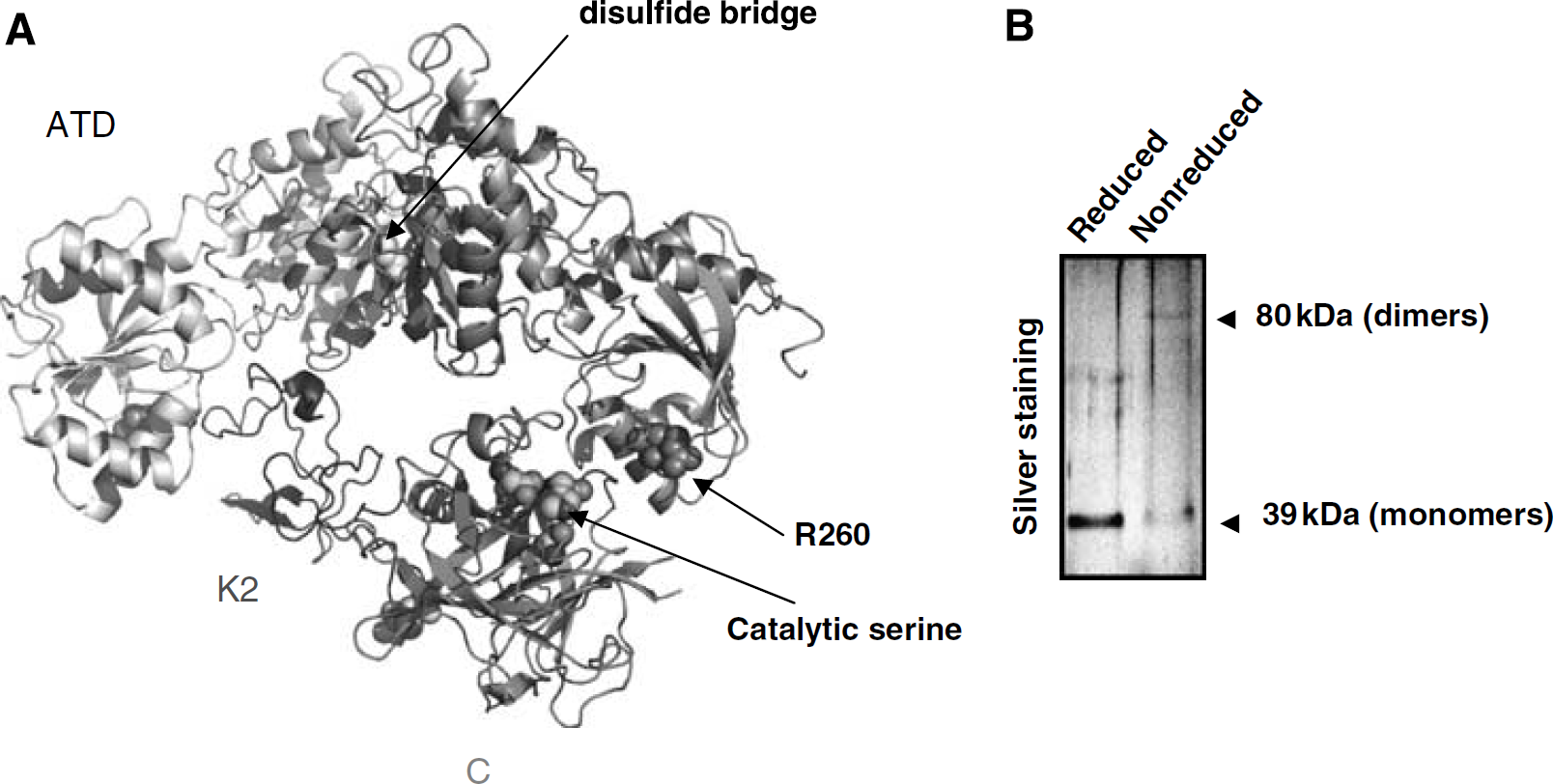
Three-dimensional model of the interaction of tissue-type plasminogen activator (tPA) with ATD-NR1. (
Peptides of the Kringle 2 Domain of tPA Prevent tPA-Induced Cleavage of NR1 without Affecting its Proteolytic Activity
Since the K2 domain of tPA contains a lysine-binding site, two lysine analogues, ε-amino caproic acid and tranexamic acid, both administered at 3 μmol/L, were tested for their ability to prevent tPA-induced cleavage of rATD-NR1. As shown in Figure 5A, both ε-amino caproic acid and tranexamic acid abolished tPA-induced cleavage of rATD-NR1 without affecting tPA proteolytic activity (assessed by its sustained ability to cleave its specific fluorogenic substrate Spectrozyme; Figure 5B). Docking experiments indicated that the K2 domain of tPA can interact with the hydrophobic pocket of rATD-NR1. In the proposed model, the interaction energy of each K2 residue with rATD-NR1 was calculated and revealed several K2 regions capable of a strong interaction with rATD-NR1. On the basis of this study, three peptides were synthesized (Figure 5C). Peptides 1 and 3 correspond to the sequences of two loops identified directly as interacting regions. Peptide 2 is a C-terminally elongated form of peptide 1 to which the sequence D-G-D was added using a G-G linker. For all three peptides, conservation of their 3D conformations as well as their interaction potency was verified by energy minimization calculation. Interestingly, peptides 1 and 3 did not influence the cleavage of rATD-NR1 by tPA, whereas peptide 2 completely prevented the cleavage (Figure 5D). Importantly, none of these peptides were able to influence the catalytic activity of tPA on its conventional substrate, Spectrozyme (Figure 5E). These data are in agreement with the proposed 3D model, confirming the importance of the K2 domain of tPA in the interaction with NR1 and the direct involvement of the amino acids located at positions 270 to 274 of the K2 domain of tPA.
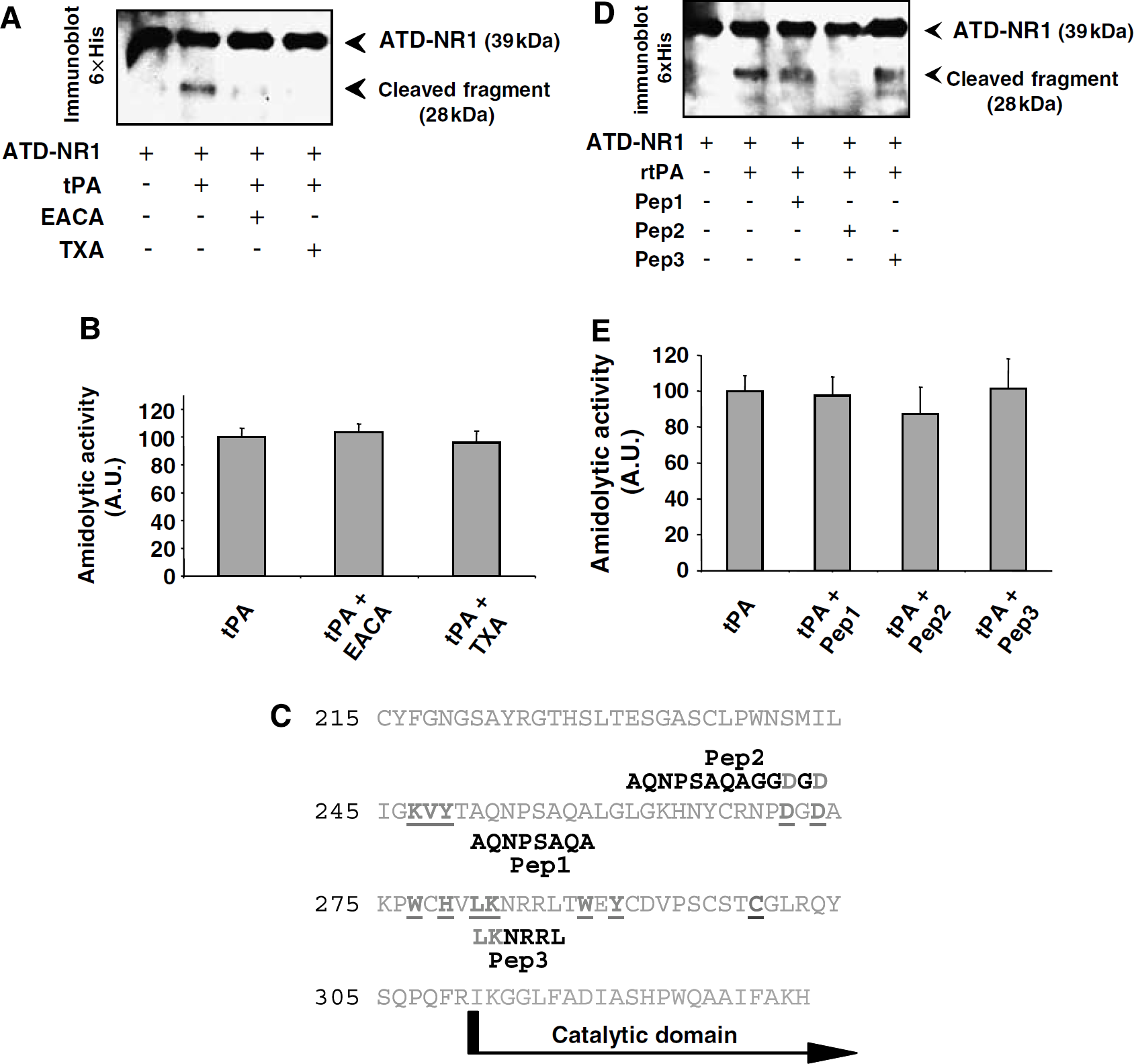
Lysine analogs and peptide mimicking part of the kringle 2 domain of tPA prevent tPA-induced cleavage of ATD-NR1. (
In Contrast to tPA, DSPA does not Promote NMDA Receptor-Mediated Signaling
Although both tPA and DSPA display thrombolytic properties, one binds and cleaves NR1 (tPA) whereas the other does not (DSPA). Based on these observations, their respective influence on glutamatergic neurotransmission was investigated in pure primary cultures of mouse cortical neurons (at 14 days in vitro) challenged by the ionotropic glutamatergic agonist NMDA (10 μmol/L). N-methyl-
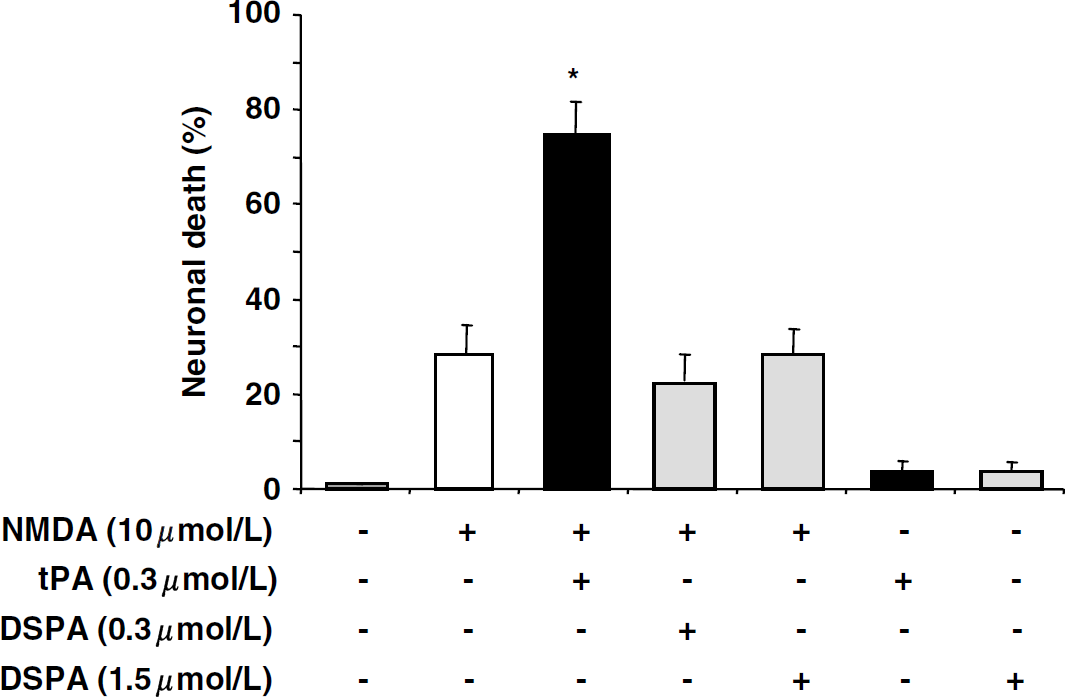
In contrast to tissue-type plasminogen activator (tPA), desmoteplase (DSPA) does not promote N-methyl-
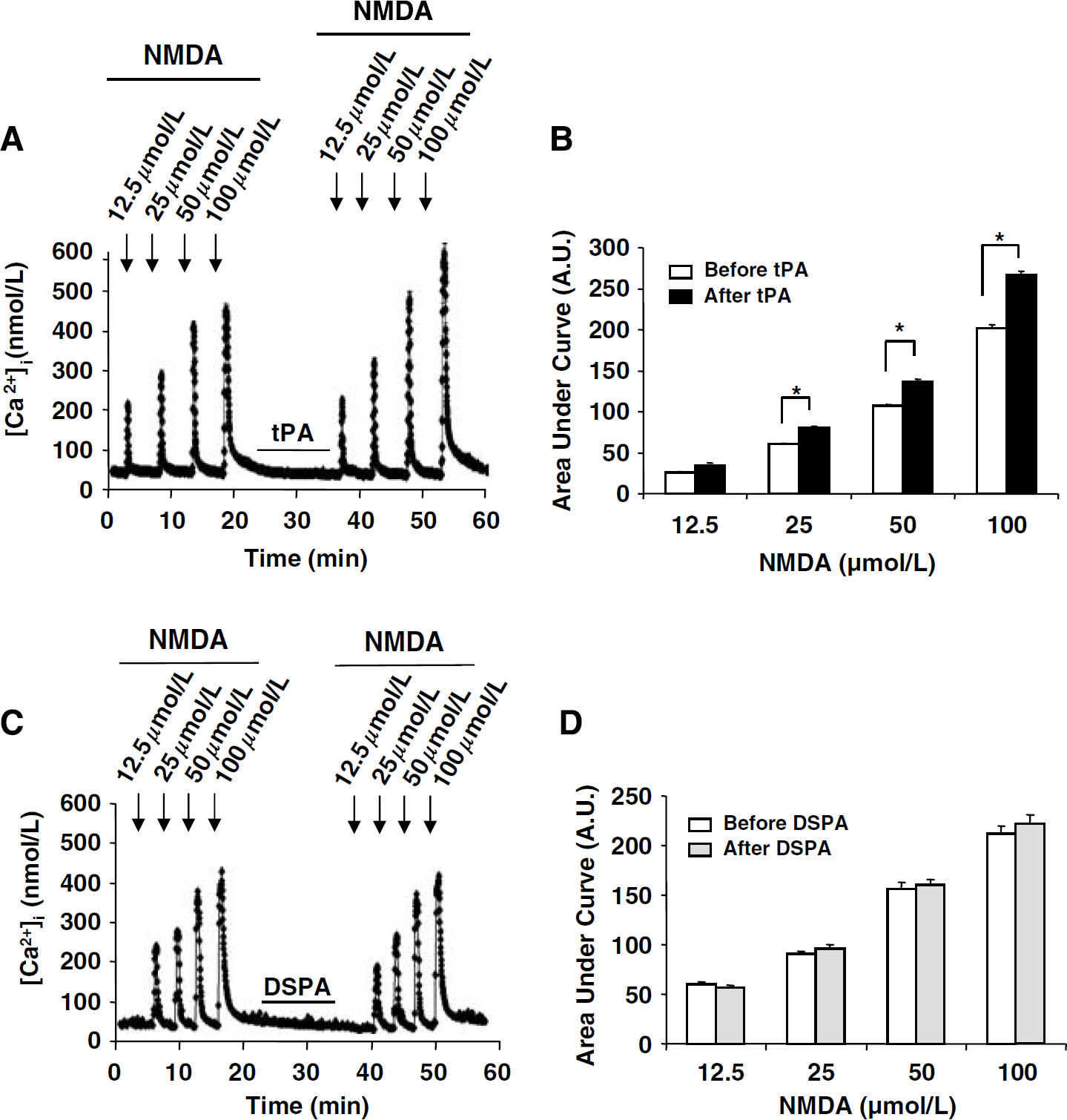
In contrast to tissue-type plasminogen activator (tPA), desmoteplase (DSPA) does not promote N-methyl-
Discussion
Stroke is the third leading cause of death and the leading cause of adult morbidity in industrialized nations. As a consequence, stroke constitutes a major social and economic burden to society. Early reperfusion, using intravenous administration of tPA within 3 h of stroke onset (The National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group, 1995), is currently the only therapeutic intervention in ischemic stroke. Although its efficacy cannot be questioned, seminal investigations in animal models on the mechanism of neuronal death have pointed out that tPA may increase the infarct volume by promoting excitotoxic cascades (Tsirka et al., 1996; Benchenane et al., 2004; Kaur et al., 2004). In addition, others and we have shown that when injected intravenously, exogenous tPA can cross both intact and injured blood—brain barrier to reach the brain parenchyma (Benchenane et al., 2005a, 2005b; Harada et al., 2005). Indeed, even in the absence of any brain injury, exogenous tPA can be detected in the brain parenchyma as early as 45 mins after its intravenous injection over 15 mins, whereas the vascular compartment is cleared within 1 h (Benchenane et al., 2005a). Importantly, although this vascular tPA invading the brain parenchyma is not toxic by itself, it occurs at levels sufficient enough to aggravate excitotoxic injuries in vivo (Nicole et al., 2001; Benchenane et al., 2005a, 2007; Armstead et al., 2006), suggesting that even short exposures to tPA can promote brain damages. Although a number of independent laboratories have reported, by using either in vitro systems or in vivo models, that tPA may promote NMDAR signaling (for review, see Benchenane et al., 2004; Samson and Medcalf, 2006), one of the questions that remains to be elucidated is to assess whether tPA can form a stable and direct complex with NMDAR. Based on our previous publications showing that tPA can cleave the NR1 subunit of NMDAR in primary cultures of cortical neurons in the absence of plasminogen (Fernandez-Monreal et al., 2004; Nicole et al., 2001), we have further addressed this issue by using a set of complementary approaches, including SPR. Our results suggest that tPA can form a two-sites stable and direct complex with ATD-NR1 (KD=24 nmol/L).
The second issue was to determine the molecular requirements of this interaction. Our results suggest that although several agents have roughly comparable thrombolytic advantages, some might not carry proexcitotoxic risks. Indeed, in contrast to tPA, TNK, and reteplase, DSPA does not bind to NR1. Accordingly, our data show that the K2 domain of tPA is critical for binding with NR1. This might be a feature common to other tPA-mediated actions, as it was recently reported that the K2 domain of tPA is also required for the interaction and cleavage of latent platelet-derived growth factor CC at Arg231 (Fredriksson et al., 2005).
We have previously identified Arg260 of NR1 as the sensitive cleavage site necessary for subsequent potentiation of NMDAR signaling by tPA (Fernandez-Monreal et al., 2004). Based on the influence of the K2 domain of tPA, our 3D model of the interaction of tPA with NR1 suggests that tPA binds first with one NR1 subunit through its K2 domain leading to the presentation of its catalytic domain to Arg260 of a second NR1 subunit before promoting cleavage. These data argue regarding the existence of a dimer of NR1, as previously shown in native receptors (Meddows et al., 2001). Moreover, the experiments show that no more than 50% of the NR1 subunit can be cleaved by tPA.
Another important point of our present investigations is that peptides that mimic the K2 domain of tPA containing the tPA lysine-binding site can be used to prevent both the interaction and cleavage of NR1 by tPA without affecting the proteolytic activity of tPA per se.
In conclusion, here we provide the first description of the molecular requirements by which the serine protease tPA, previously characterized as a neuromodulator (Benchenane et al., 2004), can indeed modulate NMDAR signaling. The present data may lead to the generation of compounds that will allow thrombolysis devoid of secondary neurotoxic effects. Such molecules may be able to antagonize binding of the tPA K2 domain to NR1 or, conversely, plasminogen activators that are unable to bind NR1. The data presented here also suggest a molecular explanation for the absence of neurotoxic effects of DSPA in several in vitro and in vivo studies (Liberatore et al., 2003; Reddrop et al., 2005; Lopez-Atalaya et al., 2007), a feature likely to contribute to an increased tolerance and safety of the use of thrombolytic drugs currently employed in clinical situations (Furlan et al., 2006; Hacke et al., 2005). Although the safety of DSPA (90 and 125 μg/kg) administered in the 3 to 9 h time-window after the onset of stroke symptoms was confirmed in a recently completed phase III trial (DIAS-2), there was a surprising lack of clinical efficacy. As a first approximation, this was thought to relate to the unusually high placebo response rate of about 50% (16th European Stroke Conference in Glasgow). Meanwhile, an in-depth analysis has shown that a major reason for the similar clinical outcome across the different dose groups including placebo seems to lie in a high percentage of DIAS-2 patients lacking visible occlusion in major cerebral arteries. A recent press release from companies developing the compound announced that a new clinical phase III trial with DSPA in acute ischemic stroke is being planned.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.