
Abstract

Commentary
Huberfeld G, Menendez de la Prida L, Pallud J, Cohen I, Le Van Quyen M, Adam C, Clemenceau S, Baulac M, Miles R. Nat Neurosci 2011;14:627–634.
The mechanisms involved in the transition to an epileptic seizure remain unclear. To examine them, we used tissue slices from human subjects with mesial temporal lobe epilepsies. Ictal-like discharges were induced in the subiculum by increasing excitability along with alkalinization or low Mg(2+). During the transition, distinct pre-ictal discharges emerged concurrently with interictal events. Intracranial recordings from the mesial temporal cortex of subjects with epilepsy revealed that similar discharges before seizures were restricted to seizure onset sites. In vitro, pre-ictal events spread faster and had larger amplitudes than interictal discharges and had a distinct initiation site. These events depended on glutamatergic mechanisms and were preceded by pyramidal cell firing, whereas interneuron firing preceded interictal events that depended on both glutamatergic and depolarizing GABAergic transmission. Once established, recurrence of these pre-ictal discharges triggered seizures. Thus, the subiculum supports seizure generation, and the transition to seizure involves an emergent glutamatergic population activity.
The mechanisms governing the transition of neural firing from the interictal state to the ictal state (ictogenesis) are largely unknown. However, mechanistic information about this transition is critical for several reasons. First, such details would enhance the accurate prediction of seizure occurrence from clinical and EEG data. Second, this information would help to identify factors that enhance or limit the generation of seizures. Third, knowledge about the interictal-to-ictal transition could aid the development of directed therapies.
Interictal spikes are brief paroxysmal discharges that arise from synchronous neuronal firing. In addition to denoting neuronal hyperexcitability, interictal discharges cause transient cognitive impairment (1) and comprise the electrical substrate for chronic epilepsy (2, 3). Interictal spikes are considered to be a predictor of the site of ictal onset, though scalp-recorded EEG in humans does not always localize the seizure-onset zone (4). Therefore, reduced systems can be used experimentally to study the role of interictal neuronal activity. In both animal models and organotypic cultures of epilepsy, interictal spikes develop prior to the onset of the first seizure and are considered to be a marker for epileptogenesis (5–7). However, the frequency of interictal spikes does not necessarily correlate with imminent seizure occurrence. That is, interictal spikes may indicate chronic excitability and epileptogenesis, but not the impending occurrence of an actual seizure. What, then, would be the usefulness of documenting the location and frequency of interictal spikes? Or, is there another electrophysiological marker that accurately presages ictal occurrence?
Huberfeld and colleagues approach these questions with a complex yet elegant series of studies using brain slices of resected hippocampal/limbic tissue from patients undergoing temporal lobectomy for refractory epilepsy. Brain slices containing hippocampus, presubiculum, subiculum, and entorhinal cortex were subjected to a convulsant stimulus (high potassium, increased HCO3− to alkalinize the perfusate, or low magnesium, or a combination of these agents). Each of these experimental conditions produced region-specific interictal discharges (IIDs) and ictal-like discharges; in addition, a third type of discharge was identified, which they named “pre-ictal discharges” (PIDs). PIDs represent a novel form of neuronal firing that directly precedes an electrographic seizure, thus representing a possible key to the interictal-to-ictal transition.
The primary finding of this article is that prior to the ictus, two distinct patterns of neuronal firing are discernible in extracellular recordings: IIDs and PIDs. These two firing patterns are best illustrated by a figure from the paper (Figure 1). IIDs and PIDs could be differentiated by their electrographic characteristics, onset site within the slice, and pharmacologic responsiveness, leading the investigators to postulate that different mechanisms underlie interictal versus pre-ictal activity. The physiological underpinnings of the two patterns were explored in this system using intracellular recordings and blockers of various aspects of excitatory and inhibitory neuro-transmission. Furthermore, presurgical intracranial EEG studies from the same patients demonstrate similar build-up of pre-ictal discharges in the subiculum, confirming the similarity of these two phenomena in situ and in vitro. Table 1 summarizes differences between these two firing patterns.
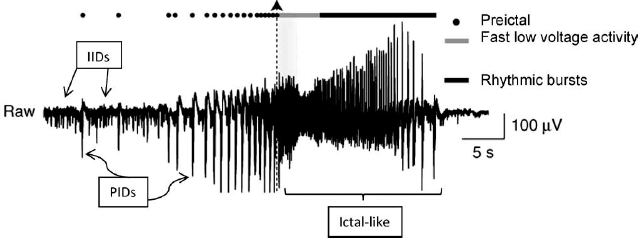
Electrographic recording from subiculum in brain slice from a patient with temporal -lobe epilepsy. Slice is superfused in 65-mM HCO3− and 8-mM K+. Recording shows interictal discharges (IIDs), build-up of pre-ictal discharges (PIDs, indicated by dots), and ictal-like discharges. Ictal-like discharges begin at dotted vertical line with arrowhead and are so named because an “ictal” event is by definition clinical, and this recording is from a brain slice. Ictal-like discharges consist of initial low voltage fast activity (gray line) followed by rhythmic bursts (solid black line). Reproduced in modified form with permission from Huberfeld, et al., Nat Neurosci 2011;14:627–634.
Differences Between Interictal and Pre-Ictal Discharges in Human Epileptic Tissue
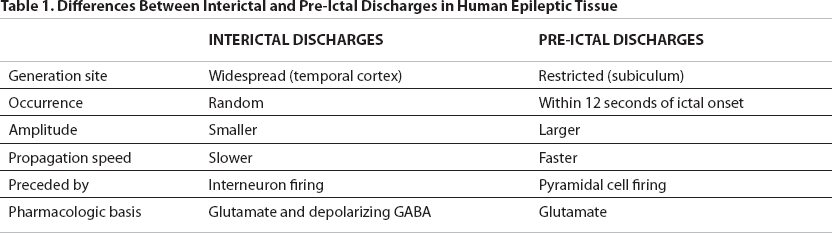
IIDs, recorded throughout limbic structures, have a random temporal distribution and do not increase in frequency as a seizure approaches. IIDs depend on both glutamatergic and depolarizing GABAergic activity, since they could be blocked by the NMDA receptor antagonist AP-5 or the GABA receptor blocker bicuculline.
PIDs, by contrast, have higher amplitudes, suggesting that they are produced by the firing of a larger number of neurons. PIDs are generated solely in the subiculum. They are mediated by glutamatergic activity only (they were not blocked by bicuculline); NMDA receptors are necessary for the initiation of PIDs but not their maintenance. PIDs progressively increase in amplitude as the ictal onset approaches (see Figure 1) and invariably precede an ictal event, as opposed to IIDs, which do not have a temporal correlation with electrographic seizures. The electrical seizure itself (low voltage fast activity, rhythmic bursts, or both, see Figure 1) comprises a separate event.
In summary, this report presents convincing physiological evidence supporting the existence of a pre-ictal state distinguishable from the interictal state, with a different physiological basis responsible for each state. The findings have major treatment implications, since interictal activity is typically resistant to suppression by antiseizure agents, which block seizures only (5). The demonstration of a pre-ictal phase in human epileptic tissue and the similar findings from in situ EEG recordings of the same patients add to the validity of this model. Epilepsy research has historically placed excessive reliance on investigations of epilepsy mechanisms in brain slices from normal animals. The severed connections in such preparations diminish their ability to truly mimic the chronic epileptic state. The recent trend to study epilepsy mechanisms in epileptic animals is a welcome advance (8). Despite the experimental challenges of studying human tissue in vitro (limited tissue availability, lack of adequate control tissue, etc.), such investigations are essential and worth the effort. Organotypic cultures, with intact circuitry, circumvent this problem and yet maintain the experimental advantages inherent in in-vitro preparations (5).
Some questions arise. Why does this preparation require chemoconvulsant stimuli to generate IIDs, PIDs, and electrographic seizures? Presumably, the tissue, although epileptic by virtue of the epilepsy surgery evaluation, differs when removed from its native environment, structurally or physiologically. Nevertheless, the fact that the network needs an extra “jolt” to exhibit seizure activity in vitro must be taken into account.
What is the role of interneurons in PIDs? GABAergic transmission is needed for IIDs but not for PIDs, so what accounts for the transient lack of inhibitory function during the pre-ictal state? Perhaps feed-forward inhibition is overwhelmed by excessive pyramidal cell firing. Future studies to elucidate the overall network structure and function (e.g., whether distinct networks mediate IIDs and PIDs) should provide clues to these questions.
Finally, what is the clinical importance of these results? The presence of a definable pre-ictal firing pattern that reliably precedes electrographic seizure activity, and can be induced by exogenous electrical stimulation of the subiculum in slices, establishes the existence of a novel, targetable phenomenon in human temporal lobe epilepsy. The use of human tissue satisfies the critique that mechanistically oriented in vitro studies of seizure generation usually use normal tissue exposed to a convulsant stimulus. Future work should evaluate the hypothesis that interictal activity is a marker for epileptogenesis, while pre-ictal discharges govern the emergence of intermittent seizures in a predisposed brain.