
Abstract
Background
Secreted frizzled-related protein 1 (sFRP1), a negative regulator of the Wnt signaling pathway, is frequently inactivated in human gastric cancer. Genetic variants in the 3′ untranslated region (UTR) of the gene may influence the strength of miRNA binding and the regulation of mRNA transcription, affecting the individual's cancer risk. This study aims to investigate the impact of variants in the 3′ UTR of sFRP1 on the gastric cancer susceptibility in a Chinese population.
Patients and methods
The association between 2 sFRP1 gene variation loci (rs1127379 and rs10088390) with minor allele frequency more than 0.1 in the 3′ UTR and gastric cancer risk was assessed in a case-control study including 419 gastric cancer cases and 571 healthy controls. PCR-restriction fragment length polymorphism analysis was used for genotyping; the odds ratio and 95% confidence interval were calculated to estimate the relative risk.
Results
Compared with the AA genotype, the GG genotype of rs1127379 was significantly associated with a reduced risk of gastric cancer overall. In the subgroup analysis, the protective effect of the GG genotype was also found for noncardia cancer and intestinal gastric cancer. Furthermore, haplotype analysis showed that the Ars1127379 Grs10088390 haplotype conferred a risk effect for gastric cancer.
Conclusions
Genetic variants at the sFRP1 gene may be involved in gastric tumorigenesis, especially in noncardia and intestinal gastric cancer. Further prospective studies with different ethnicities and large sample sizes are needed to confirm our findings.
Introduction
Gastric cancer (GC) is the fourth most common cancer and the third leading cause of cancer-related death worldwide. According to GLOBOCAN estimates, a total of 951,600 new cases and 723,100 deaths occurred in 2012 (1). Although a steady decline in GC incidence and mortality rates has been observed in both more and less developed countries, the incidence rates are very high in Eastern Asia, such as Japan and China (35.4 in males and 13.8 in females per 100,000). The mechanism of gastric carcinogenesis is still not clear. It has been generally acknowledged that Helicobacter pylori infection, tobacco smoking, alcohol consumption, and low intake of fresh fruits and vegetables play important roles in the risk of GC. However, exposed to the same environment, only a small proportion of people eventually develop GC. Recent studies have focused on the association between genetic variants and GC risk, and genome-wide association studies (GWAS) have identified several GC susceptibility loci, including 1q22, 3q13.31, 5p13.1, 8q24.3 and 10q23 (2–4). However, these single-nucleotide polymorphisms (SNPs) are often located in the noncoding or intergenic region with unknown biological relevance and GWAS often focus on peak signals, while signals with relatively moderate levels may be neglected. In this regard, a gene-based analysis strategy could play an important role in exploring cancer susceptibility, and more efforts should be directed at the variants around genes/regions already implicated in carcinogenesis, especially those in functional regions.
The Wnt signaling pathway is an evolutionarily highly conserved signal transduction system and appears to be one of the most relevant cellular pathways involved in cancer stem cell growth and differentiation (5). Secreted frizzled-related proteins are glycoproteins that contain an N-terminal domain homologous to the cysteine-rich domain (CRD) of the frizzled family of Wnt receptors and a COOH-terminal domain with some homology to netrins (6). The CRD of secreted frizzled-related protein 1 (sFRP1) competes with the frizzled receptors for Wnt binding, modulating the signal (7). Meanwhile, sFRP1 is the evolutionarily conserved target of the Hedgehog-GLI signaling pathway. Hedgehog is secreted from differentiated epithelial cells to induce sFRP1 expression in mesenchymal cells, which keeps differentiated epithelial cells away from the effects of canonical Wnt signaling. CpG hypermethylation of the sFRP1 promoter during chronic persistent inflammation and aging could break down the Hedgehog-dependent Wnt signal inhibition and lead to the occurrence of gastrointestinal cancers, such as GC and colorectal cancer (8).
The sFRP1 gene was proposed to lie at 8p11.21, and the 3′ end of the sFRP1 mRNA was found to be alternatively spliced (6). sFRP1 gene expression is silenced by aberrant methylation of the promoter region in several cancers and loss of expression was shown to correlate with lymph node metastases and increased mortality in patients with breast cancer (9) and patients with GC (10). Recently, the 3′ untranslated region (UTR) of its mRNAs was found to be the target of several microRNAs, and microRNAs function posttranscriptionally by interacting directly with the 3′ UTRs of mRNAs to repress their expression by translational inhibition, mRNA degradation, or both. For instance, miRNA27a could inhibit sFRP1 expression and lead to the accumulation of -catenin (11), and miRNA206 suppressed GC cell growth and metastasis through targeting the sFRP1 gene (12). These observations led to the hypothesis that genetic variation in the 3′ UTR of sFRP1 affecting the expression and/or function of sFRP1 may influence the GC risk. In this study, 2 SNPs (rs1127379 and rs10088390) in the 3′ UTR of sFRP1 with minor allele frequency (MAF) more than 10% were selected, and the association between these SNPs and GC susceptibility was evaluated in a case-control study with 419 GC cases and 517 cancer-free controls in an eastern Chinese population.
Material and methods
Study participants
A total of 419 GC patients and 571 cancer-free healthy controls recruited from among genetically unrelated ethnic Han Chinese were enrolled from January 2008 to July 2012 in Nanjing City, Jiangsu Province. All patients had histopathologically confirmed primary GC. Intestinal and diffuse subtypes were identified by the criterion of Lauren's classification; unclassified cases were those with a mixed type or not available for classification. Gastric cardia cancers were defined as tumors located within 20 mm distal to the gastroesophageal junction (13). The 571 controls were enrolled to participate in the study from among hospital visitors who came to the health examination clinic for an annual checkup at the same hospital (Jiangsu Provincial Hospital of Traditional Chinese Medicine) during the same period. A 3-5 mL venous blood sample was collected from each study participant after they had signed an informed consent form. The study was approved by the institutional review board of Jiangsu Provincial Hospital of Traditional Chinese Medicine.
SNP selection and genotyping
A public SNP database (http://www.ncbi.nlm.nih.gov/) was used to select SNPs with MAF>10% in the HapMap HCB population (HapMap Genome Browser release #28) in the 3′ UTR of the sFRP1 gene, and 2 SNPs (rs1127379 and rs10088390) were eligible for inclusion.
Genotyping was performed by the PCR-restriction fragment length polymorphism (RFLP) method. The primers for rs1127379 were sense: 5′-GAA AGT TGA CAT GGG TGG GG-3′ and antisense: 5′-TGC AGA GAA TTA AGA ATC CGG C-3′, which generated a 237-bp product. The PCR product was then digested by Bsp1286I (Bio-Rad Laboratories) and separated on a 2% agarose gel stained with SYBR® Safe DNA Gel Stain (Invitrogen). The G allele resulted in 2 fragments of 81 bp and 156 bp, and the A allele produced 1 fragment of 237 bp. The primers for rs10088390 were sense: 5′-GGA CGG ATC AGG AGA CCT TG-3′ and antisense: 5′-TGC TAG TCA GGG TGA ATT GTG-3′. The amplified product (177 bp) was then digested by DdeI (Bio-Rad Laboratories). The C allele produced 2 fragments of 137 bp and 40 bp, while the G allele resulted in a single 177-bp fragment. To validate the accuracy of the PCR-RFLP genotyping method, 10% of the samples were randomly selected to confirm the assays by the same method, and the results of repeated assay should be completely consistent with the original assay.
Statistical analyses
The Hardy-Weinberg equilibrium was tested by a goodness-of-fit chi-square test to compare the observed genotype frequencies with the expected ones among the control group. Differences in the distribution of demographic characteristics, selected variables and genotypes of the sFRP1 variant between GC cases and controls were evaluated using a 2-sided chi-square test. Logistic regression analyses were employed to estimate crude and adjusted odds ratios (ORs) and 95% confidence intervals (CIs) for the association between sFRP1 genotypes and risk of GC by age and sex. The PHASE 2.0 program was used to infer haplotype frequencies based on the observed sFRP1 genotypes. The Quanto program (version 1.2.3, University of Southern California) was employed to calculate the power of the OR on the GC risk. A p value less than 0.05 was considered statistically significant. Analyses were performed with the Statistical Analysis System software (version 9.1.3; SAS Institute).
Results
A total of 419 GC cases and 571 cancer-free controls were enrolled. The mean ages of patients and controls were 59.9 ± 11.3 and 53.7 ± 13.4 years, respectively. The proportion of male and female subjects was 65.5% and 34.5% for GC cases, and 64.1% and 35.9% for controls. Smokers accounted for 39.1% of GC cases and 30.6% of healthy controls. Drinkers of alcohol accounted for 22.9% of GC cases and 14.7% of healthy controls. Of the 419 GC patients, 299 (71.4%) were defined as having noncardia cancer and 109 (26.0%) as having cardia GC according to disease location; the data of 11 patients (2.6%) were not available for classification. According to the WHO classification, the histopathological features were classified as tabular adenocarcinoma (154 cases, 36.75%), mucinous adenocarcinoma (26 cases, 6.21%), signet ring cell carcinoma (31 cases, 7.4%), and adenocarcinoma not otherwise specified (208 cases, 49.73%). For the convenience of data operation, we divided the histological types into intestinal GC (321, 76.6%), diffuse GC (73, 17.4%), and unclassified GC (25, 6%) according to Lauren's classification.
The genotype distributions of sFRP1 rs1127379 and rs10088390 in the GC cases and controls are shown in Table I. The observed genotype frequencies for the 2 polymorphisms were both in Hardy-Weinberg equilibrium in the controls (p = 0.602 and p = 0.604, respectively). The rs1127379 genotype frequencies were 42.7% (AA), 46.3% (AG) and 11.0% (GG) in the GC cases and 37.1% (AA), 46.1% (AG) and 16.8% (GG) in the cancer-free controls; the difference was statistically significant (χ2 = 7.652, p = 0.022). Logistic regression analysis revealed that the GG homozygote genotype was associated with a significantly reduced risk of GC (adjusted OR = 0.53, 95% CI 0.35-0.81) compared with the AA wild-type homozygote, while the recessive model (GG vs. AA/AG) showed that the GG genotype was also in significant association with GC (adjusted OR = 0.55, 95% CI 0.37-0.81). No significant association was detected between rs10088390 genotypes and risk of GC (adjusted OR = 1.07, 95% CI 0.81-1.43 for CG vs. CC; adjusted OR = 1.12, 95% CI 0.77-1.64 for GG vs. CC). We further evaluated the genetic effects of the 2 loci on GC cases according to the 2 confirmed histological subtypes (intestinal and diffuse) and found that the protective effect of the rs1127379 GG genotype remained significant in intestinal GC cases (adjusted OR = 0.53, 95% CI 0.34-0.84) but not in cases of the diffuse type (p>0.05) compared with the wild-type AA genotype (Tab. II). When the GC cases were divided into noncardia and cardia cancer, we found that the protective effect was significant in the noncardia cases (adjusted OR = 0.52, 95% CI 0.33-0.82) but not in the cardia GC cases (p>0.05) (Tab. III).
sFRP1 gene polymorphisms and the risk of gastric cancer
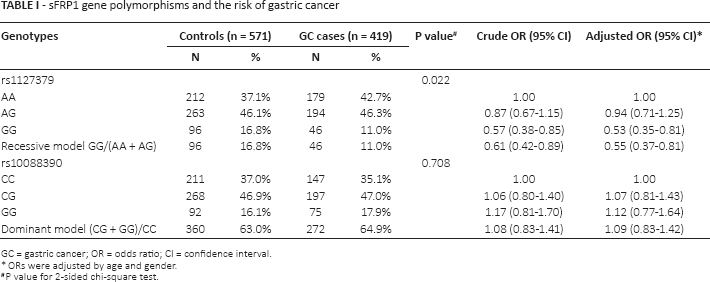
GC = gastric cancer; OR = odds ratio; CI = confidence interval.
ORs were adjusted by age and gender.
P value for 2-sided chi-square test.
Gene polymorphisms of sFRP1 and the risk of different histological types of gastric cancer
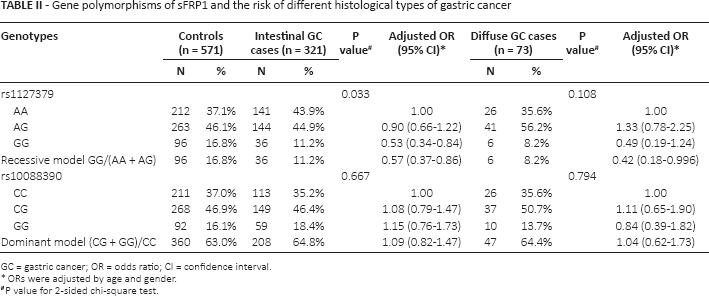
GC = gastric cancer; OR = odds ratio; CI = confidence interval.
ORs were adjusted by age and gender.
P value for 2-sided chi-square test.
Gene polymorphisms of sFRP1 and the risk of gastric cardia and noncardia cancers
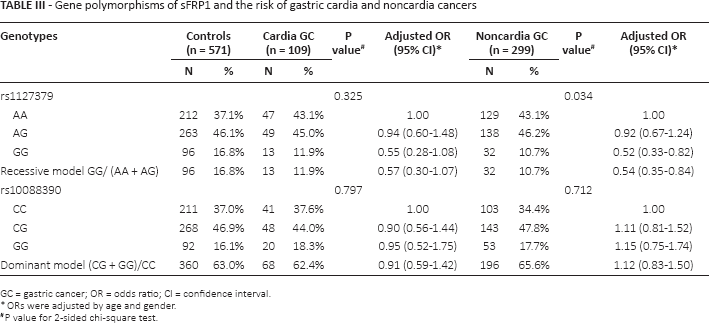
GC = gastric cancer; OR = odds ratio; CI = confidence interval.
ORs were adjusted by age and gender.
P value for 2-sided chi-square test.
In the stratified analysis (Tab. IV), the protective effect of the rs1127379 GG genotype remained significant in men, drinkers and smokers, and both younger and older patients. In addition, we did not find any obvious effect for the rs10088390 on risk of GC in different strata (data not shown).
Stratified analyses of SNP rs1127379 genotypes between gastric cancer cases and controls
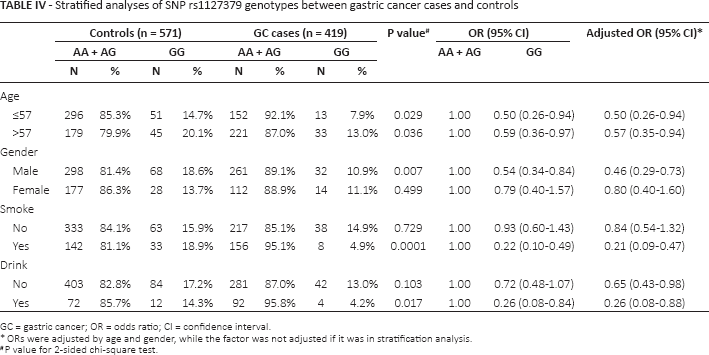
GC = gastric cancer; OR = odds ratio; CI = confidence interval.
ORs were adjusted by age and gender, while the factor was not adjusted if it was in stratification analysis.
P value for 2-sided chi-square test.
Using the PHASE 2.0 program, we inferred 4 haplotypes on the observed genotype data. The Ars1127379 Grs10088390 haplotype showed a risk effect for GC compared with the most common haplotype Ars1127379 Crs10088390; the OR was 2.54 (95%CI 1.76-3.67) (Supplementary Table S1 - Haplotype distributions of rs1127379 and rs10088390 among GC cases and controls. Available online at www.tumorijournal.com).
Based on our current sample size, the statistical power was calculated by the Quanto software. We found that the statistical power reached 91.66% for OR = 0.53 and 88.66% for OR = 0.55. The correspondence parameters were G allele frequency (0.4530) for rs1127379, the estimated GC prevalence (31.8/100,000) in China (14), and recessive mode of inheritance. For rs10088390, based on the G allele frequency of 0.3958 and the same GC prevalence, the statistical power for OR = 1.07 was 7.56%, for OR = 1.12 it was 12.27%, and for OR = 1.09 it was 9.17%. Although the statistical power for rs10088390 was low, the higher statistical power for rs1127379 demonstrated that the sample size was adequate to evaluate the association between rs1127379 and GC risk.
Discussion
The canonical Wnt/β-catenin pathway regulates a variety of elementary cellular functions such as proliferation, polarity, adhesion and motility during development, differentiation and adult tissue homeostasis (15). Aberrant signaling in the Wnt, Hedgehog and epidermal growth factor pathways are responsible for the GC stem cell phenotype, which are believed to be the only cells that live long enough to accumulate the multiple genetic changes required for transformation (16). sFRP1 is a target gene of the Hedgehog pathway and is involved in cross-talk between the Hedgehog pathway and the Wnt pathway; it can positively or negatively regulate tumorigenesis and tumor progression. Reduced sFRP1 levels and transcripts have been frequently found in breast (9), bladder (17) and gastric neoplasms (10). Our analysis based on TCGA (The Cancer Genome Atlas, http://cancergenome.nih.gov) data further revealed that the sFRP1 mRNA expression level in gastric adenocarcinomas was significantly lower than that in adjacent normal tissues (Supplementary Figure 1 - sFRP1 mRNA expression analysis between 32 paired gastric adenocarcinoma and adjacent normal tissues based on TCGA data. Available online at www.tumorijournal.com), suggesting a potentially vital role of sFRP1 in gastric carcinogenesis. Epigenetic alteration of sFRP1 was reported to play an important role in gastric carcinogenesis. Methylation of sFRP1 was inversely correlated with gene expression (18). As a tumor suppressor, hypermethylation of sFRP1 was frequently found in tumors compared with adjacent normal tissue (19). No genetic variants have been reported so far as being associated with GC. In the present study, rs1127379 located in the 3′ UTR of the sFRP1 gene was identified to be significantly associated with an altered risk of GC in a Chinese population. Furthermore, we provided important clues that genetic variants in sFRP1 might also be involved in the susceptibility to GC and highlight the potential pleiotropic effect of sFRP1 variants on multiple cancers.
Several studies have reported the association of SNPs in sFRP1 with bone mineral density (BMD) (20), bone mineral content (21), and the risk of bladder cancer (22). Four SNPs (rs921142, rs4736965, rs10106678 and rs7832749) in sFRP1 were associated with bone density, and 2 SNPs (rs3242 and rs16890444) were related to lumbar spine BMD values. The T allele in SNP rs3242 was shown to confer an increased risk of early-onset bladder cancer and to influence the genotype-dependent microRNA capacity on sFRP1 mRNA. However, those SNPs had a relatively low frequency in the Asian population (MAF<10%) or in the intron regions and were not eligible in the present research. In our study, the G allele in SNP rs1127379 with an allele frequency of 45.3% in individuals of Asian descent showed a protective effect for GC compared with the more common A allele (54.7%). Although the decrease in GC risk is modest (approximately 0.53-fold in GG homozygotes), the contribution of rs1127379 to the GC incidence in Asian populations is extremely important. The 3′ UTR is, however, an area for binding sites of proteins and microRNAs that affect the stability and transport of the mRNA. According to the web-based SNP analysis tool, SNPinfo (http://snpinfo.niehs.nih.gov/snpinfo/snpfunc.php), the base change of A to G may alter binding to mature hsa-miR-1182, hsa-miR-216a or hsa-miR-220c to its target mRNA (23). Functional studies reported that miR-1182 can inhibit GC proliferation and migration by targeting the ORF1 of hTERT (24), while miR-216a can regulate the JAK/STAT pathway by targeting JAK2, contributing to pancreatic carcinogenesis and tumor growth (25). Based on the evidence above, we can draw the conclusion that the base change of A to G for rs1127379 may modify the transcriptional regulation of sFRP1 and then alter sFRP1 protein expression by influencing the miRNA binding.
Several methodological issues and limitations of this study need to be discussed. First of all, in a case-control setting, inherent selection bias cannot be completely excluded. Meanwhile, in our study the mean age of GC cases was significantly higher than that of the healthy controls, and age itself may be an independent risk element for GC. However, the odds value of the GG genotype of rs1127379 was not influenced after adjusting for age, indicating that potential confounding factors related to age were minimal. Secondly, fresh tumor tissues of the GC cases were not obtained in this study and gene expression related to different genotypes of rs1127379 could not be analyzed in the same population with the SNP association study. Thirdly, only 2 potentially functional SNPs of sFRP1 were investigated, which did not cover all variants in the sFRP1 gene, especially for those minor allele frequencies less than 5%. Finally, epigenetic alteration in gastric carcinogenesis was not taken into account in this study.
In conclusion, our case-control study revealed that SNP rs1127379 in the 3′UTR of sFRP1 was associated with gastric noncardia and/or intestinal cancer in a Chinese population. This provides further important evidence that genetic variants in the sFRP1 gene contribute to GC susceptibility. Larger well-designed studies with ethnically diverse populations are warranted to confirm our findings.
Footnotes
Abbreviations
Financial support: This work was supported by the Science Foundation for Youth of Nanjing University of Chinese Medicine (13XZR04); Natural Science Foundation of the Jiangsu Higher Education institution of China (14KJD330001); Research and innovation program of graduate students in Jiangsu higher education institutions (CXLX13_576); National Natural Science Foundation of China (81473458, 81473593 and 81171987); Research Fund for the Doctoral Program of Higher Education of China (20133234110007); and a project funded by the Priority Academic Program Development of Jiangsu higher education institutions (PAPD).
Conflict of interest: None of the authors has any conflict of interest related to this work.