
Abstract
Recent studies have demonstrated the influence of clock genes in cell cycle regulation, cell proliferation, apoptosis and DNA damage recognition and repair. There is evidence suggesting the implication of clock genes in colorectal cancer (CRC) development and progression. The aim of this study is to evaluate the expression levels of clock genes in CRC and correlate them with patients' prognosis. Forty-two CRC samples (from 24 males and 18 females), their paired noncancerous tissues and 8 biopsies from healthy individuals were included. Quantitative real-time PCR was used to examine the expression levels of CLOCK1, BMAL1, PER1, PER2 and PER3 genes in all the samples. In the cancerous tissues CLOCK1 (p<0.0001) and BMAL1 (p<0.0001) expression levels were higher, while PER1 (p<0.0024) and PER3 (p<0.0001) expression levels were lower compared to matched healthy tissues. No difference was observed in the expression levels of PER2 (p=0.99). No correlation was found between clock gene expression and patients' clinicopathological characteristics or prognosis. The results suggest abnormal expression of CLOCK1, BMAL1, PER1 and PER3 genes in CRC but no correlation with patients' prognosis.
Introduction
The circadian rhythm is an internal timing system with a period of 24 hours permitting mammals to adjust to a variety of environmental modifications. In humans the central circadian oscillator is located in the suprachiasmatic nuclei of the brain in the anterior hypothalamus, whereas every single cell is believed to be a peripheral circadian clock (1). The circadian master coordinates the numerous peripheral circadian clocks directly through autonomic nervous system regulation and indirectly through hormonal modulation (2). Interestingly, recent studies demonstrate that the peripheral clocks can preserve their circadian activity without the appropriate control of the central pacemaker, suggesting the presence of an autonomous mechanism based on the interaction between cellular components (3).
The molecular mechanism of circadian regulation in the cellular level is based on a negative feedback model including a variety of circadian genes, mainly BMAL1, CLOCK, NPAS2, casein kinase 1 epsilon (CK1∊), period circadian protein homolog 1 (PER1), period circadian protein homolog 2 (PER2), cryptochrome 1 (CRY1) and cryptochrome 2 (CRY2), which interact in a continuous cycle of 24 hours. In particular, BMAL1, CLOCK and NPAS2 promote the formation of BMLA1/CLOCK and BMLA1/NPAS2 heterodimers, which bind to the promoters of the CRY and PER genes, activating their transcription, and to the promoter of the BMAL1 gene, inhibiting its transcription (4). CK1∊ is known to phosphorylate PER protein, resulting in its degradation by ubiquitination. CRY accumulates in the cytoplasm, promoting the formation of CK1∊/CRY/PER complexes, which seem to enter the nucleus and disrupt the BMAL1/CLOCK-associated transcriptional complex resulting in the inhibition of CRY and PER transcription (1).
Recent studies suggest that a variety of cell cycle mediators, tumor suppressor genes and oncogenes are regulated by circadian rhythm agents, which explains the observation that the circadian system is involved in the control of DNA repair, apoptosis and cell proliferation (5). In particular, PER1 and PER2 proteins inhibit the expression of c-myc, which is implicated in cyclin D1 expression and cell proliferation (6), whereas the tumor suppressor protein RORα is known to be a transcriptional target of the BMAL1/CLOCK complex (7). On the other hand, PER1 protein is believed to be implicated in DNA repair mechanisms while c-erbB-2 and p53 were suggested to act as downstream players for PER in terms of tumor progression (8). Finally, the CK1∊ kinase phosphorylates and destabilizes β-catenin, a molecule implicated in the Wnt pathway, which is known to play a critical role in the molecular pathogenesis of colorectal cancer (CRC) (9).
CRC is one of the most common malignancies and one of the leading causes of cancer-related death worldwide. Despite the better understanding of the molecular basis of the disease, surgical therapy remains the main therapeutic approach. Adjuvant chemotherapy is currently used for stage III and IV patients as well as high-risk stage II patients. It is clear though that new prognostic markers could be highly valuable especially for patients with stage II disease. There are data suggesting a clear implication of circadian gene mutations in the pathogenesis of CRC (10), while downregulation of BMAL1 in cell cultures seems to increase cell proliferation, decrease apoptosis and increase resistance of CRC cells to chemotherapy (11). Recent studies demonstrated that the expression of these genes in CRC may influence the prognosis of the disease (12–14). CRC tissues presented lower expression of PER1, PER2 and CRY2 compared to healthy samples while high expression of another circadian gene, known as the TIM gene, was correlated with poor prognosis (12). Low PER2 expression may be related to increased cancer invasiveness, advanced stage, higher grade and more intense Ki-67 staining (13). Increased BMAL1 expression and decreased PER1 and PER2 expression may be related to liver metastasis in patients with CRC (14).
The aim of this study is to compare the expression of CLOCK1, BMAL1, PER1, PER2 and PER3 between CRC patients and healthy matched controls and to correlate the expression of these circadian genes with clinical and pathological features of CRC patients.
Materials and Methods
Study subjects
Forty-two CRC samples (from 24 males and 18 females) and paired noncancerous tissues were obtained from patients who had undergone colectomy for cancer at Hippokrateio Hospital, Athens, Greece. Furthermore, 8 biopsies from healthy individuals were also included in the study. After surgical removal, the samples were stabilized with RNAlater (QIAGEN GmbH, Hilden, Germany). All tumors were sporadic, as assessed by personal and family histories. All patients underwent surgery following histological diagnosis at colonoscopic biopsy and had not received any neoadjuvant chemo- or radiotherapy before operation. The samples were surgically obtained between 9:00 and 12:00 am. The age of the patients, tumor site, tumor type, histological grade and TNM stage according to the American Joint Committee on Cancer (AJCC) classification were recorded. The study was approved by the Hospital Ethical Committee and all patients gave their written informed consent to the sample collection and analyses described in the present study.
RNA isolation, complementary DNA synthesis, and quantification
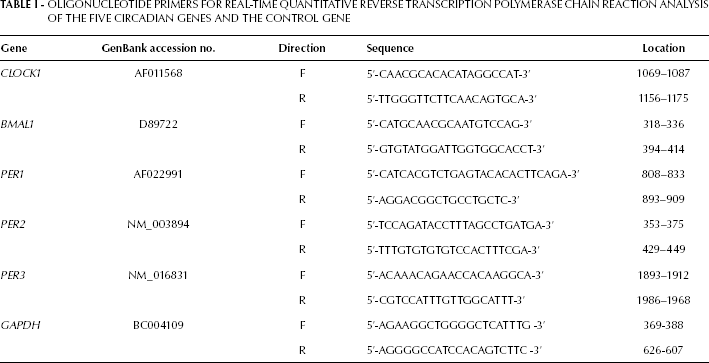
The isolation of total RNA was performed using the AllPrep DNA/RNA/Protein mini kit (Qiagen) according to the manufacturer's instructions. Hereafter, reverse transcription was performed by incubating 1 μg total RNA for 1 hour at 42°C in the presence of 500 μg/mL of Oligo dT 12-18, 10 mM deoxyribonucleotide triphosphates, 5 × first-strand buffer, 0.1 M dithiothreitol, and 200 U/mL of MMLV reverse transcriptase (Invitrogen, Darmstadt, Germany). Assessment and normalization of the CLOCK1, BMAL1, PER1, PER2 and PER3 mRNA levels was performed by adjusting the expression levels of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a reference gene. All specific sets of primers (15) were obtained from Applied Biosystems (Applied Biosystems, Foster City, CA, USA). The sequences of the forward and reverse primers are listed in Table I. Real-time PCR was performed in an ABI PRISM 7000 Sequence Detection System (Applied Biosystems) as follows: initial denaturation for 2 minutes at 50°C and 10 minutes at 95°C, followed by 40 cycles of PCR (95°C for 15 seconds and 60°C for 1 minute). Reactions were performed in duplicate using the SYBR Green PCR Master Mix (Applied Biosystems) according to the manufacturer's instructions. The analysis was randomly repeated to ensure the repeatability of the results. Data were analyzed using the comparative Ct method for the relative quantitation of results. The levels of gene expression were normalized after subtracting the Ct value of the GAPDH internal control from that of the examined genes' Ct value for samples (ΔCt = |Ctgene(samples) - CtGAPDH|). In order to compare the expression levels of each gene between the samples tested, the ΔΔCt value was determined using the formula (ΔΔCt = ΔCtgene(sample A) - ΔCtgene(sample B)). Then the relative level of each gene in cancer samples was compared with that in normal samples using the formula 2ΔΔCt. Post-amplification denaturation curves showed that the primer pairs generated single products.
Oligonucleotide Primers for Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction Analysis of the Five Circadian Genes and the Control Gene
Statistical analysis
Data are shown as mean ± SD. Differences in clock gene expression between CRC patients and controls were analyzed using the Mann-Whitney test. Correlations were calculated using Spearman's correlation coefficient. All p values were 2-tailed and p values less than 0.05 were considered significant. All calculations were performed using GraphPad version 3.00 (GraphPad Software Inc., San Diego, CA, USA).
Results
Table II summarizes the histopathological characteristics of the CRC samples. To address whether mRNA levels of CLOCK1, BMAL1, PER1, PER2 and PER3 were impaired in CRC cases, we compared the transcript levels of these 5 clock genes in patients having CRC with those of the paired nontumor tissues and the tissues from healthy individuals (Fig. 1). No significant differences in gene expression were observed between the paired nontumor tissues and tissues from healthy individuals. The expression levels of the circadian genes were categorized as low or high according to their median values. CLOCK1 and BMAL1 expression levels were higher in cancer than in healthy normal tissues (p<0.0001 and p<0.0001, respectively). PER1 and PER3 expression levels were higher in normal tissues (p=0.0024 and p<0.0001). PER2 expression levels were similar in cancer and normal tissues (p=0.99).
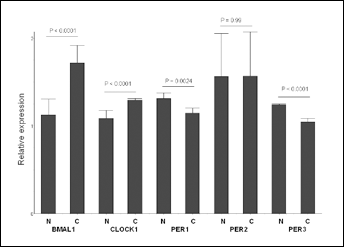
Comparison of circadian gene mRNA expression levels between colorectal cancer tissues (C) and normal tissues (N).
Histopathological Characteristics of Colorectal Cancer Samples at Diagnosis

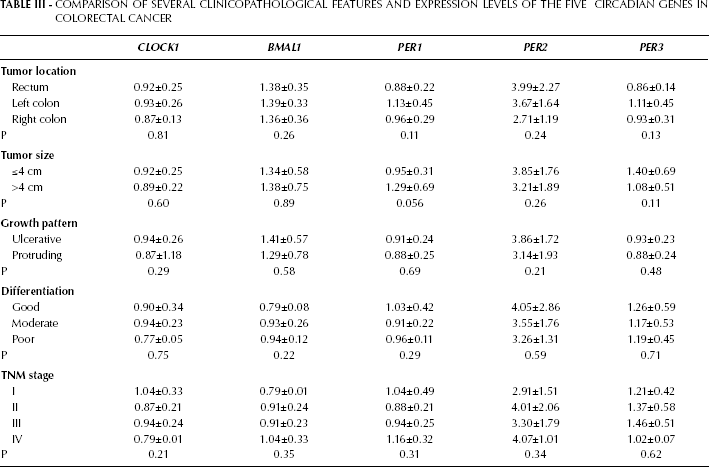
The relations between the expression levels of clock genes and clinicopathological features were also examined. In our cases, the expression levels of the clock genes were unrelated to tumor size, growth pattern, differentiation, TNM stage, and patient survival (Tab. III).
Comparison of Several Clinicopathological Features and Expression Levels of the Five Circadian Genes in Colorectal Cancer
Discussion
Circadian rhythms regulate various human functions and may represent a critical control point for cancer development and tumor progression (16). Recent reports suggest that circadian clock genes regulate the expression and the transcription activity of a variety of cell cycle genes including C-MYC, TP53 and PAK2, especially in rapidly dividing cells like intestinal cells and bone marrow precursors. In particular BMAL1 is believed to suppress the expression of p21, which is a negative regulator of the G1/S phase of the cell cycle, while BMAL1-/- knock-out mice were found to have dramatically increased and no longer rhythmic expression of p21 (7). Lin et al showed that the overexpression of BMAL1 in NIH-3T3 cells led to increased expression of β-catenin and upregulation of the canonical Wnt pathway, which is known to be highly related to CRC (17). Recently, Elshazley et al considered BMAL1 as a potential therapeutic target in malignant mesothelioma, showing that knocking down of this gene in MPM cells downregulated cyclin B, induced apoptosis and reduced cell growth and proliferation, thereby causing mitotic catastrophe (18). These previous reports are in agreement with our results detecting higher BMAL1 expression in CRC tissues in comparison with matched healthy tissues. Interestingly, Taniguchi et al showed that BMAL1 expression is reduced by promoter methylation in hematological malignancies but not in solid tumors and that the overexpression of BMAL1 suppresses the growth of lymphoma cell lines, suggesting a tumor suppressor role of BMAL1 in hematological malignancies (19). Finally, according to a recent report, vascular endothelial growth factor, which is considered to play an important role in CRC pathogenesis, is transcriptionally upregulated by BMAL1 (20).
PER1 is known to be implicated in cell growth and DNA repair mechanisms, which are critical in the development of CRC as well. In particular, Gery et al highlighted the importance of this molecule in sensitizing cells to DNA–damage-induced apoptosis mainly by activating checkpoint 2 kinase (ChK2) through ATM regulation. They also showed that ectopic expression of PER1 suppressed the growth of the colon cancer cell line SW84 and suggested that at least some of these activities were p53 dependent (8). Moreover, Mazzoccoli et al and Oshima et al found decreased expression of PER1 and PER3 in CRC tissues (12, 14).
Our results are in agreement with the above reports as PER1 and PER3 expression was found to be significantly decreased in cancer tissues, suggesting that these genes may play a tumor suppressor role in CRC. Interestingly, Krugluger et al showed that decreased PER1 and PER2 expression in high-grade CRC was correlated with low expression of dihydropyrimidine dehydrogenase, a critical enzyme in terms of 5-fluorouracil metabolism. The authors suggested that the low expression of this enzyme can be predictive of increased 5- fluorouracil toxicity in these patients (21). Finally, according to another study, CRC tissues from mice were found to have decreased expression of PER1 and PER2 while the rhythmicity of BMAL1 expression was completely abolished (22).
We also observed higher expression of the CLOCK1 gene in CRC compared to healthy tissues, which is in agreement with previous data (14). The CLOCK1 transcription factor is known to create heterodimers with BMAL1 in the nucleus, leading to increased transcription of PER and CRY genes. It is believed that the CLOCK1 gene remains steady throughout the 24-hour day and high levels of BMAL1 promote the formation of this heterodimer, which has many transcriptional targets including cyclin D1, a known promoter of cancer cell proliferation (23). On the other hand, PER proteins are believed to play a suppressor role in terms of clock gene expression. This negative feedback can explain our observation that lower PER1 and PER3 expression is accompanied by higher CLOCK1 expression in CRC tissues.
We could not identify a statistically significant difference in PER2 expression between cancer and healthy tissues. Wang et al in a recent report demonstrated lower expression of PER2 in CRC tissues with a heterogeneous pattern of staining. They also showed that downregulation of PER2 may be related to the development, invasion and metastasis of CRC (13). On the other hand, Krugluger et al did not find any significant difference in PER2 expression between healthy and cancerous tissues (21).
Interestingly, the majority of recent reports demonstrated a correlation between clock gene expression and clinicopathological characteristics of CRC patients like TNM stage, grade and clinical outcome. In particular, Mazzoccoli et al suggested that lower expression of PER1 and PER2 is correlated with a poor prognosis and lower survival rates (12), while Oshima et al found that lower expression of PER2 is related to a worse clinical outcome and higher expression of BMAL1 and lower expression of PER1 are related to increased incidence of liver metastasis (14). In our study there was no correlation between the expression of clock genes and TNM stage, differentiation, incidence of liver metastasis and clinical outcome. For this divergence in the association with prognosis, one should seriously take into consideration the relatively small sample of patients included in our study and the low incidence of metastatic CRC amongst those patients.
In conclusion, our results suggest that BMAL1 and CLOCK1 are upregulated in CRC whereas PER1 and PER3 are downregulated. According to our study the expression levels of BMAL1, CLOCK1, PER1, PER2 and PER3 are not correlated with any clinicopathological characteristics of CRC patients nor with their clinical outcome. Further experimental and clinical studies will aid in fully understanding the impact of circadian genes on the pathogenesis of CRCas well as developing interesting biomarkers in terms of prognosis and discovering new targets for therapeutic interventions.