
Abstract
Reactive oxidants damage the retinal pigment epithelium (RPE), which is required for viability of overlying photoreceptors. Smoking which leads to chronic accumulation of reactive oxidants in the circulation is linked to age-related macular degeneration (AMD) where RPE death is seen along with photoreceptor loss in the central macular region of the retina. It is unclear why this damage is concentrated in the central retina. We asked whether circulating oxidant might specifically target the central retina. Mice were administered the classic reactive oxidant iodate through tail vein injection, and visual acuity was followed by optokinetic response. Histology and apoptosis was examined by H&E and immunostaining. Iodate indeed selectively damaged the central retina, and this damage was highlighted by early apoptosis of RPE in the central retina followed by apoptosis of photoreceptors adjacent to the region of RPE loss–-cones were lost preferentially. The pattern and extent of this damage was independent of exposure to light. We then conclude that circulating oxidant is sufficient to selectively damage the central retina highlighted by sequential apoptosis of RPE and photoreceptors, with cones being the most sensitivity to this RPE loss.
Introduction
Age related macular degeneration (AMD) is the leading cause of blindness in patients over the age of 65 in the US.1,2 There are two forms: wet and dry. Dry AMD is more common, and approximately 85% of intermediate and advanced AMD consist of the dry. One of the earliest indicators of dry AMD is loss of retinal pigment epithelium (RPE), which is essential to maintain viability of the adjacent photoreceptors. AMD has been genetically linked to components of the immune response including the complement cascade.3,4 But, it is unclear if such immune system involvement is a primary initiating event in AMD or a secondary response, and to date a genetic animal model which recapitulates key features of AMD is not available. 5 Because of the late onset of AMD, it is thought that chronic exposure to an environmental factor(s) may be an initiating event. 6 Indeed, AMD is strongly linked to smoking, which classically leads to chronic accumulation of circulating oxidants.7,8 The RPE is particularly sensitive to oxidative damage, and classic oxidants such as iodate selectively damage the RPE when introduced in the circulation. 9 11 Taken together, the results raise the possibility that RPE is a target of oxidant damage, and that photoreceptor loss is secondary to the initial RPE damage.
A central pattern of retinal damage is a hallmark of AMD, but little is known of why such damage is confined to the central retina. It has been proposed that UV irradiation, which also generates free radicals leading to oxidative damage, focuses light entering the eye onto the central retina and contributes to the central retinal specificity seen in AMD. 12 However, there is an absence of definitive evidence linking UV exposure to AMD, although there may be a correlation between general exposure to blue light and the age of AMD onset. 13 Alternatively, it is possible that the central retina is simply more sensitive to an environmental factor such as circulating oxidant than the peripheral retina. An animal model which recapitulates the features of AMD is not available, so addressing AMD progression and this central retina-specific pattern experimentally in an AMD model is not yet possible. As noted above, smoking which leads to chronic accumulation of circulating oxidants is genetically linked to AMD. But, it is unclear whether exposure to a circulating oxidant would be sufficient to selectively damage the central retina.
Previous studies have demonstrated that injection of the reactive oxidant sodium iodate leads to death of the retinal pigment epithelium (RPE) and a loss of vision. 14 Here, we show that injection of iodate into the circulation leads to rapid apoptosis of the RPE in the central retina of the mouse. This in turn is followed by apoptosis of underlying photoreceptors, which depend upon the RPE, and loss of vision. Cones are lost preferentially compared to rods, implying that they are more sensitive to RPE loss than rods. This central pattern and time course of sequential RPE and photoreceptor loss following iodate injection was independent of exposure to light. These results then provide evidence that circulating oxidant is sufficient to specifically damage the central retina.
Materials and Methods
Animals
C57BL (B6) mice were purchased from Jackson Laboratories (Bar Harbor, ME). The animals were handled in accordance with policies and procedures recommended by the Institutional Animal Care and Use Committee at the University of Louisville, and all the procedures adhered to the ARVO Statement for the Use and Care of Animals in Ophthalmic and Vision Research. The animals were housed in the standard animal room under a 12-hour-light/12-hour-dark cycle. All animals used in this study were males and 6–8 wks old. Sodium iodate (Sigma-Aldrich, St. Louis, MO) was injected intravenously at a concentration of 0.4 mg/ml. Experiment groups received 20 or 25 mg/kg of sodium iodate. The control group was injected with phosphate buffered saline. Twenty mice (40 eyes) were used in each experiment group and in control groups.
Histology and nuclei counting
Following euthanasia, the eyes were marked at the 12 o'clock meridian to maintain orientation, and they were then enucleated and fixed in 10% formalin. Fixed eyes were then paraffin embedded and sectioned at 5-μm. Sections were stained with H&E, and the and the number of cells in the ONL and INL were counted. The cell numbers of the central retina (or posterior pole) and peripheral retina were counted at regions approximately 250 um distance from the optic nerve or the ora serrata, respectively. A total of 20 eyes were used for each group, randomly, 3 pictures at each position were taken for cell counts.
Immunostaining
Paraffin slides were deparaffinized, rehydrated to distilled water, and then subjected to antigen retrieval by microwaving in 10 mM citrate buffer. The slides were blocked before applying the primary antibody. The slides were incubated overnight at 4 °C or 1 h at room temperature with primary antibodies in blocking solution. After the slides were washed, secondary antibodies were applied for 1 h at room temperature. The primary antibodies used were: mouse anti rhodopsin (Millipore, 1:300), rabbit anti red/green opsin (Millipore 1:500). The secondary antibodies were CY3 conjugated goat anti mouse antibody or goat anti rabbit antibody. The nuclei were counterstained with Dapi. The images were captured with a Zeiss inverted fluorescence microscope (Axiovert 2000). As negative controls, no immunostaining was evident in the absence of primary antibodies.
Optokinetic Reflex (OKR)
OKR was performed using the OptoMotry system (CerebralMechanics, Lethbridge, AB, Canada). 15 For measurement of visual acuity, animals were assessed for this tracking behavior until unambiguous tracking was observed. Spatial frequency of the grating was increased until the animal no longer responded. The highest spatial frequency the mouse could track was identified as the threshold.
Apoptosis assay
Apoptotic cells were detected by incorporation of fluorescein-12-dUTP at the 3′-OH DNA ends using the terminal deoxynucleotidyl transferase recombinant enzyme (rTdT) and the cell nuclei were stained with Dapi. 16 Five hundred cells were counted for each specimen, and the apoptotic index (percentage of the apoptosis nuclei) was calculated as the percentage of fluorescent nuclei.
Effect of light on iodate damage
Mice were either maintained in a dark environment 24 hour a day for 2 wks following sodium iodate injection, or they were maintained on a standard 12 hours light/12 hours dark cycle. In the control group, the mice were housed in a conventional animal room (temperature 18 °C–24 °C) with a 12/12 h light/dark cycle (lights on: 07:00 h, light intensity at shelf level approximately 100 lux; lights off: 19:00 h, light intensity approximately 2 lux). In the dark group, mice keep totally in the dark. Animals were sacrificed after two weeks for histology and cell counting.
Statistical analysis
The SEM was calculated and compared by one way ANOVA. Differences were considered significantly at P < 0.05.
Results
Introduction of the reactive oxidant iodate into the circulation leads to a concentration-dependent loss of visual acuity
To assess the effect of sodium iodate on vision, mice were analyzed using OKR. Multiple concentrations of sodium iodate were tested. We saw little effect on the retina with less than 20 mg/kg, and results with 25 mg/kg and greater concentrations were similar, but increased toxicity was evident with dosages greater than 25 mg/kg. Therefore, we compared 20 and 25 mg/kg doses of sodium iodate. With 20 mg/kg, there was only a partial effect on visual acuity. Increasing the iodate dosage to 25 mg/kg led to a more severe loss of vision. Introduction of an oxidant into the circulation then leads to a persistent loss of vision, and these two concentrations of iodate cause damage ranging from partial to more extensive loss of vision (Fig. 1).
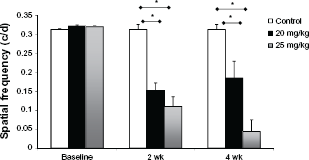
Iodate causes a decrease in visual function in mice. OKR measurements are shown at two and four weeks after iodate injection.
Subsequent to loss of RPE nuclei in the central retina, there is loss of the adjacent ONL
While there was no detectable effect on the ONL one day after iodate injection in the central retina, there was a significant decrease in the ONL and the rows appeared disorganized three days after injection in this region (Figs. 2A and 4 below). The number of ONL rows was further reduced after one and two weeks, and no recovery in the ONL was evident at four weeks (Fig. 2A–C). By contrast, there was relatively little effect of either iodate concentration on the ONL in the peripheral retina at any of the time points, and the boundary of morphologic ONL loss between the central and peripheral retina appeared to be sharp (Fig. 2B). As a control, the INL was unaffected by iodate in either the central or peripheral retina. We conclude that iodate specifically affects the ONL in the central retina.

Iodate injection leads to a sharply defined boundary of ONL loss in the central vs. peripheral retina. (
Rapid central retina-specific loss of RPE following iodate injection
One day after iodate injection, Dapi-stained RPE nuclei were no longer evident in the central retina (Fig. 3A and B). Despite this loss of Dapi+ nuclei, continuous RPE pigment was still evident, although these pigmented ghosts of the RPE were condensed compared to the untreated retina. These pigmented ghosts of RPE were still evident at day 3, but by day seven they were significantly disrupted. By contrast to the central retina, RPE nuclei were still evident in the peripheral retina two weeks after iodate injection. These results demonstrate rapid RPE nuclear disruption in the central retina following iodate injection, whereas RPE nuclei in the peripheral retina were unaffected.

The RPE is lost in the central vs. peripheral retina following iodate injection. (
RPE cells in the central retina undergo apoptosis at day one following iodate injection
Tunnel immunostaining was used to assess apoptosis in the central and peripheral retina following iodate injection. Consistent with the loss of Dapi-staining RPE nuclei at day one, most nuclei in the central retina were apoptotic at this time (Fig. 4). Apoptosis of the RPE was no longer evident at day three and subsequent times consistent with loss of the RPE nuclei at day one (Fig. 3A and B). By contrast, few apoptotic cells were evident in the RPE at any of the time points in the peripheral retina (Fig. 4). As with the boundary demarcating loss of the ONL in central vs. peripheral retina (Fig. 2B), there was also a sharp boundary between apoptosis of the RPE in these regions (Fig. 4, top left panel). These results demonstrate very rapid apoptosis of the RPE in the central retina following iodate injection.

RPE in the central retina undergo rapid apoptosis following iodate injection. TUNEL immunostaining is shown at the indicated times following injection of 20 mg/kg of iodate. Nuclei are stained with Dapi. Arrows indicate RPE nuclei. On the top row, the left-hand boxed region in the left panel is shown at higher power in the following two panels. The middle and lower rows show TUNEL immunostaining in the central and peripheral retina at 3 and 7 days following iodate injection as indicated.
Photoreceptors in the central retina undergo apoptosis subsequent to RPE loss after iodate injection
Few apoptotic cells were evident in the ONL one day after iodate injection (Fig. 5A–C). However, apoptotic cells were evident in the ONL in the central retinal three days after iodate injection consistent with a diminished number of nuclei in the ONL at this time (Fig. 2A–C). Although apoptotic cells were still evident in the ONL four weeks after injection, the highest percentage of apoptotic cells occurred at day three–-the percentage of apoptotic cells decreased at day seven and it decreased further at 4 weeks (Fig. 5C). After four weeks, the ONL was reduced to 1–2 rows, but it was not eliminated. Consistent with the lack of morphologic effects of iodate on the ONL in the peripheral retina (Fig. 2A–C), little apoptosis was evident in the ONL in the peripheral retina (Fig. 5). There is a significant difference in apoptosis between the central and peripheral retina (Fig. 5B). As a control, apoptosis did not occur in the INL following iodate injection. These results demonstrate that apoptosis of the RPE in the central retina precedes apoptosis of adjacent photoreceptors in this region.

iodate injection leads to apoptosis of the ONL in the central retina, but little apoptosis is evident in the ONL in peripheral retina and apoptosis is absent in the INL. (
Iodate leads to selective loss of cones in the central retina
Cone photoreceptors are highly concentrated in the human macula (ie, central retina) and are selectively lost during AMD. Mice contain a rod-dominant retina and cones are not concentrated centrally but instead are distributed throughout the retina. We wondered if cones in the central retina were selectively lost compared to rods in this region after iodate injection. Therefore, we immunostained retinal sections for the cone marker red/green opsin and the rod marker rhodopsin. Iodate had no detectable effect on rods vs. cones in the peripheral retina (Fig. 6). By contrast, no immunostaining for red/green opsin was detected in the central retina–-all of the remaining ONL cells in the central retina were rhodopsin-positive indicating that they are rods. 17 However, rhodopsin expression in these remaining ONL cells was redirected to the cell body with no outer segment apparent, suggesting that these remaining rods in the central retina may not be functional (Fig. 6).

Loss of opsin and re-arrangement of rhodopsin at 7 days and beyond in central retina. Immunostaining for cone-specific red/green opsin and rod-specific rhodopsin is shown at the indicated times following 20 mg/kg iodate injection.
Iodate-induced damage to the central retina is independent of light
We reasoned that exposure of mice to light may synergize with iodate to specifically damage RPE and in turn photoreceptors in the central retina. Therefore, following iodate injection mice were either maintained in a control group on a 12 hr light/dark cycle as above, or they were maintained in a dark environment. After two weeks, retinal histology was compared in the two groups (Fig. 7). We found that maintaining the mice in a dark environment had no detectable effect on the central vs. peripheral damage to the RPE and ONL, implying that exposure to light is not contributing to the central retinal-specific effects of iodate.

Light does not affect retinal damage following iodate injection. (
Discussion
It has been demonstrated that the classic reactive oxidant iodate leads to RPE damage and photoreceptor loss.18,19 Here, we show that this damage is confined to the central retina and it is highlighted by rapid apoptosis of RPE in the central retina followed by apoptosis of adjacent photoreceptors. And, we demonstrate that cone photoreceptors are more sensitive than rods to this loss of RPE. Such results raise the possibility that damage from environmental oxidants may be targeted at RPE in the central retina, and that chronic exposure to such oxidants may contribute to RPE loss and ultimately cell death in the overlying cones in the macula as observed in AMD.
As in the mouse, acute exposure of humans to reactive oxidants such as iodate also leads to retinal damage and loss of vision. 20 Plurodesis is done to prevent reoccurrence of penumothorax or pleural effusion. And, povidone iodine can be instilled between the parietal and the visceral layers of the pleura to trigger a fibrotic response which in turn obliterates the space between the layers, thereby preventing collapse of the lung or fluid accumulation. 21 Recently, loss of vision has been reported following pleurodesis with povidone iodine. 22 This iodate-induced retinopathy is highlighted by a sharply demarcated RPE loss confined to the region of the central fundus. Taken together, these results suggest that, as in the mouse, a circulating reactive oxidant can lead to specific death of the RPE in the central retina of humans and ultimately to vision loss. Smoking has been strongly linked to AMD, and cigarette smoke classically leads to elevated levels of reactive oxidants in the circulation.8,23 Based on the results described above, it is possible that chronic exposure to circulating oxidants from cigarette smoke may contribute to the central retina-specific death of the RPE and loss of overlying photoreceptors in AMD.
Because light is focused on the macula in man, it is possible that the combination of chronic light exposure in conjunction with circulating oxidants may synergize to damage RPE in the central retina. We did not detect any effect of light on the central retina-specific pattern of RPE damage in acute exposure to circulating iodate in the mouse, and the acute damage to the RPE in the central retina following instillation of povidone iodine in the lung was rapid did not require chronic exposure to light. Thus, light may not contribute to the centralized pattern of RPE damage.
Interestingly, the line demarking RPE and photoreceptor damage in the central vs. peripheral retina was very sharp following iodate injection, and likewise the centralized RPE damage seen following povidone iodine treatment in pleurodesis was clearly demarcated. 22 Conceivably this RPE damage boundary simply reflects differences in RPE cells in the central vs. peripheral retina with regard to their sensitivity to oxidant. However, it is also possible that such central vs. peripheral differences in RPE susceptibility to an oxidant may be due to differences in circulation in the central vs. peripheral retina leading to regional differences in toxin delivery. Recently, laser-targeted angiography has been used to examine choroidal circulation (which services the outer retina and RPE) in the rat and monkey.24,25 These studies provide evidence that the circulation is highly segmented with feeders acting as end-arteries thereby preventing cross-circulation between segments and leading to sharp boundaries between the segments. Interestingly, the arrangement of choroidal vessels is distinct in the central vs. peripheral segments–-a lobular arrangement of the choriocappillaris is seen in the central area with capillaries arranged in a spindle-shape, but at the periphery, the choroidal arterioles and venules take a parallel course, with the choriocapillaris joining the adjacent arterioles and venules at right angles in a ladder-like pattern. 26
This arrangement of vessels correlates with increased blood flow in the central vs. peripheral region. Thus, circulation to the central and peripheral RPE, and outer retina, appears highly segmented with little evidence of cross circulation between the two segments. Moreover, differences in organization of the choroidal vasculature appear to lead to altered circulation rates in these two regions of the retina, which may account for the enhanced sensitivity of RPE in the central retina to a circulating toxin. Previously, we have injected pigs with iodoacetic acid, which leads directly to death of photoreceptors due to their dependence upon glycolysis (which is blocked by iodoacetic acid at the level of glyceraldehyde 3-phosphate dehydrogenase) (Zhou et al 2011). While the aim of these studies was to provide a model of photoreceptor loss to test transplantation of induced pluripotent stem cells, during the course of the experiments we found that iodoacetic acid specifically led to loss of photoreceptors in the central retina of the pig (the peripheral retina was unaffected). Taken together these results in the mouse and pig using two different retinal toxins (one targeting RPE and the other photoreceptors directly) argue for differential delivery of circulating toxins to the central vs. peripheral retina.
In AMD the central loss of RPE is accompanied by macular degeneration leading to loss of cones, which are concentrated in this region in man. In the mouse, there is no macula or visual streak and cones are distributed throughout the retina allowing us to assess the relative effect of iodate on rods vs. cones. While rods were diminished in the central region of the retina, cones were eliminated. Therefore, we conclude that cones appear to be more sensitive to loss of RPE than rods.
Disclosures
Author(s) have provided signed confirmations to the publisher of their compliance with all applicable legal and ethical obligations in respect to declaration of conflicts of interest, funding, authorship and contributorship, and compliance with ethical requirements in respect to treatment of human and animal test subjects. If this article contains identifiable human subject(s) author(s) were required to supply signed patient consent prior to publication. Author(s) have confirmed that the published article is unique and not under consideration nor published by any other publication and that they have consent to reproduce any copyrighted material. The peer reviewers declared no conflicts of interest.
Footnotes
Acknowledgments
We thank Maureen McCall for helpful discussions and Guirong Liu for histological sections. These studies were supported in part by American Health Assistance Foundation, NIH Grants (P20 RR018733 and EY015636), Research to Prevent Blindness, and The Commonwealth of Kentucky Research Challenge.