
Abstract
B-vitamins are important for producing energy from amino acids, fatty acids, and glucose. The aim of this study was to elucidate the effects of excess vitamin intake before starvation on body mass, organ mass, blood, and biological variables as well as on urinary excretion of riboflavin in rats. Adult rats were fed two types of diets, one with a low vitamin content (minimum vitamin diet for optimum growth) and one with a sufficient amount of vitamins (excess vitamin diet). Body mass, organ mass, and blood variables were not affected by excess vitamin intake before starvation. Interestingly, urinary riboflavin excretion showed a different pattern. Urine riboflavin in the excess vitamin intake group declined gradually during starvation, whereas it increased in the low vitamin intake group. Excess vitamin intake before starvation does not affect body mass, organ mass, or blood variables but does affect the urinary excretion of riboflavin in starving rats.
Keywords
Introduction
It is generally considered that water-soluble vitamins undergo more rapid turnover than any other nutrients. Our group reported that urinary excretion levels of water-soluble vitamins reflect recent 1-week intakes of these vitamins.1–4 In addition, we reported that fasting and starvation alters urinary excretion levels of water-soluble vitamins in humans and rats.5,6 The most striking phenomena were the sharp decrease in urinary excretion of vitamin B1 in humans and rats,5,6 increased urinary excretion of vitamin B2 in humans, 5 and decreased urinary excretion of vitamin B2 in rats.5,6 These findings showed that urinary excretion levels of water-soluble vitamins do not always reflect intake or nutritional status during fasting and starvation, particularly in terms of vitamins B1 and B2. These two vitamins are the most important vitamins for producing the energy compound ATP.
Because the frequency of earthquakes is very high in Japan, people may suddenly receive notice to evacuate their locations. In such cases, Japanese people are faced with the risk of insufficient food supply. Therefore, carrying macronutrients such as protein, lipids, and carbohydrate should be recommended given that energy shortage is the source of many diseases. However, this is difficult since the weight of macronutrients required per day is large (approximately 600 g/day), while carrying micronutrients, such as vitamin and mineral tablets, is more likely since the weight required per day is low (approximately 1 g/day). Thus, we were interested in how body conditions are affected when body stores of vitamins differ before starvation. Currently, there is no information regarding how differences in body vitamin stores before starvation affect body mass, organ mass, and blood biological variables. If the body's vitamin stores before starvation were at minimal amounts for preventing deficiencies, vitamin deficiency may be accelerated by food deprivation. The aim of this study was to determine whether physiological variables are affected by body vitamin stores before starvation. Additionally, the urinary excretion of riboflavin during starvation was investigated because the urinary excretion of riboflavin has been reported to increase in humans during starvation.5,7
Methods
Diets
The two chemically defined diets were designated as shown in Table 1. Vitamin-free milk casein, L-methionine, and sucrose were purchased from Wako Pure Chemical Industries (Osaka, Japan). Corn oil was purchased from Nisshin OilliO Group, Ltd. (Tokyo, Japan). Gelatinized cornstarch, the mineral mixture (AIN-93G), 8 and the vitamin mixture (AIN-93) 8 were obtained from Oriental Yeast Co., Ltd. (Tokyo, Japan).
Diet composition.

Reeves, RG, Components of the AIN-93 diets as improvements in the AIN-76A diet.
Animals
The care and treatment of experimental animals conformed to the University of Shiga Prefecture guidelines for the ethical treatment of laboratory animals. The room temperature was maintained at approximately 22 °C and 60% humidity with a 12-h/12-h light/dark cycle (06:00-18:00/18:00-06:00).
Male Wistar rats (8 weeks old, weighing 188-202 g) were obtained from CLEA Japan, Inc. (Tokyo, Japan). Rats were individually kept in a metabolic cage CT-10 (CLEA Japan) and were randomly divided into two groups. One group was fed a diet containing a sufficient amount of vitamins (10 g of the AIN-93 vitamin mixture per kg of diet; referred to as the V1.0% group) (Table 1), while the other group was fed a diet containing a minimum requirement of vitamins for optimum growth of weanling Wistar rats (3 g of AIN-93 vitamin mixture per kg of diet, referred to as the V0.3% group) 9 for 10 days. This 10-day period was used to acclimatize the rats to their new conditions and to evaluate whether food intake and body weight gain were the same between the two groups. 9 Additionally, the 10-day period was sufficiently long to result in different body vitamin stores in the rats. Food intake and body weight gain were identical and urinary excretion levels of eight water-soluble vitamins in rats fed the V0.3% were nearly zero at the beginning of starvation. Following this 10-day period, each dietary treatment group was divided into two groups, a feeding group (V1.0%-Control; V0.3%-Control) and a starvation group (V1.0%-Starved; V0.3%-Starved) for 6 days. The 24-h urine samples (09:00-09:00) were collected each day and stored at −20 °C until analysis.
All rats except starving rats had access to food ad libitum and all rats, including starving rats, had access to tap water ad libitum. Body mass and food consumption were recorded daily. Rats were sacrificed after the last urine samples had been collected. Blood samples were collected in EDTA-2 K tubes and centrifuged at 1,700 ×
Analytical methods
Plasma glucose, triglyceride (TG), blood urea nitrogen (BUN), aspartate aminotransferase (AST), alanine aminotransferase (ALT), and creatinine were measured with a FUJI DRI-CHEM 4000 Chemistry Analyzer (Fujifilm Ltd., Tokyo, Japan). The cerebrum, heart, lungs, kidneys, liver, spleen and testes were dissected and weighed. Thiamin hydrochloride was purchased from Wako Pure Chemical Industries (Osaka, Japan). Thiamin in urine was measured using the high-performance liquid chromatography (HPLC) post-label method using fluorescence detection. 10 Riboflavin was purchased from Wako Pure Chemical Industries. Riboflavin in urine was measured by HPLC. 11
Statistical analysis
Values in tables and figures are expressed as the mean ± SEM. The main effects of “fed or starved” and “vitamin intake” were analyzed using a two-way analysis of variance (ANOVA), followed by Bonferroni's post hoc test. When the two-way ANOVA indicated the presence of a “fed or starved”-“the vitamin intake” interaction, one-way ANOVA was conducted followed by Tukey's multiple-comparison test among the groups (Tables 2 and 3). Figures 2 and 3 show the results of Student's
Effect of vitamin intake on blood variables in fed and starved rats.

Effect of vitamin intake on organ mass in fed and starved rats.

Results
Food intake and body mass
Food intake during acclimatization was the same between the two groups (V1.0% vs. V0.3% group). Average daily food intake was 18.5 ± 0.8 and 18.9 ± 1.5 g/day/rat (V1.0% and V0.3% groups, respectively). Thus, vitamin intake was approximately 3:1 in the V1.0% group vs. the V0.3% group. The specific daily rate of body mass loss (dM/Mdt) during starvation is shown in Figure 1. There was no significant difference between the V1.0% and V0.3% groups.

Effect of vitamin intake on the rate of body mass loss (dM/Mdt) during starvation. Values are means ± SEM, n = 4.
Blood variables
Table 2 shows the blood variables in the fed and starved rats. Starvation-induced hypoglycemia was observed in both vitamin groups. Plasma TGs showed a similar pattern to glucose. BUN was increased by starvation. Plasma AST was not affected by starvation. Plasma ALT was increased in the V1.0%-starved group. Plasma creatinine also increased in the V1.0%-starved group.
Organ mass
Table 3 shows organ masses. In the V1.0% groups, cerebrum, lungs and testes mass were not changed by starvation. Heart, kidneys, liver, and spleen mass decreased in the starved group. In the V0.3% groups, the cerebrum, heart, and testes mass were not changed by starvation, while the lungs, kidneys, liver, and spleen mass decreased in the starved group. Both groups showed remarkable changes in liver mass after starvation.
Urinary excretion of thiamin (Vitamin B1)
Urinary excretion of thiamin during starvation is shown in Figure 2. In the V1.0% group, the urinary excretion of thiamin was significantly decreased on day 1 during starvation, and these decreased values were maintained during starvation. In the V0.3% group, the urinary excretion of thiamin was very low even before starvation and remained low during starvation.
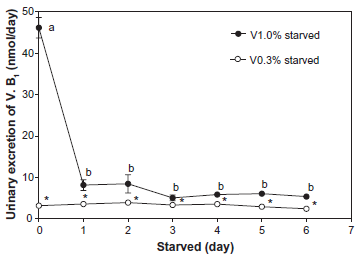
Effect of vitamin intake on the urinary excretion of vitamin B1 (thiamin) during starvation. Values are means ± SEM, n = 4. Student's
Urinary excretion of riboflavin (Vitamin B2)
Urinary riboflavin excretion during starvation is shown in Figure 3. In the V1.0% group, urine riboflavin gradually decreased during starvation, whereas that of the V0.3% group increased by approximately 3-fold.

Effect of vitamin intake on urinary excretion of vitamin B2 (riboflavin) during starvation. Values are means ± SEM, n = 4. Student's
Discussion
Vitamins, particularly vitamins B1, vitamin B2, and niacin, are important in the production of energy from amino acids, fatty acids, and glucose. Thus, body vitamin stores may be critical for energy production during starvation. The requirement of these three vitamins increases with energy consumption. 12 The aim of this study was to elucidate the effects of dietary vitamin intake before starvation on body mass, organ mass, blood, and biological variables as well as on urinary excretion of riboflavin in starved rats.
We previously reported the effect of food restriction on the conversion of L-tryptophan to niacin in rats. 13 In the study, urinary excretion of the sum of niacin and its catabolites did not decrease during starvation for 9 days; instead, these levels significantly increased on days 1 and 9 during starvation. 13 When humans or rats were starved, urinary elimination of vitamin B1 ceased and the urine value on day 1 (first 24 h of starvation) was nearly zero. 5 Therefore, we investigated the effects of vitamin intake before starvation on body mass, organ mass, and blood biological variables, as well as on the urinary excretion of thiamin and ribo-flavin, since these vitamins are involved in energy production. 12
Vitamin contents in the diet (the V1.0% diet was used as an excess vitamin diet and the V0.3% diet was used as a minimal vitamin diet for optimum growth) did not affect body mass change during acclimatization and starvation periods. Some variables, such as BUN and ALT, were higher in the starved groups than in the fed groups (used as control), regardless of vitamin intake, while differences were observed in the weights of lungs, heart, and testes between the V1.0% and V0.3% groups. However, these differences may be within the range of measurement error. Other variables were not affected by vitamin intake except urinary riboflavin excretion. Urinary excretion of riboflavin increased 2.7-fold with starvation. In a previous human study, 7 a 1-day fasting period increased the urinary excretion of riboflavin by 2.6-fold. A similar phenomenon was observed in human and rat experiments when a supplementary vitamin was not administered. This suggests that rats can be used as a model for human fasting studies.
There are limitations to the present study. We did not measure levels of some important blood variables such as hemoglobin, hematocrit, and creatinine kinase. These variables would be better indicators of body functional status. In conclusion, excess vitamin body stores before starvation did not affect body mass, organ mass, or blood variables. These findings indicate that excess vitamins in the body are not important nutrients for short periods of starvation in rats.
Funding
This investigation was part of the project “Studies on the Dietary Reference Intakes for Japanese” (principal investigator, Katsumi Shibata), which was supported by a Research Grant for Comprehensive Research on Cardiovascular and Life-Style Related Diseases from the Ministry of Health, Labour and Welfare of Japan.
Author Contributions
Designed the study: AM, TF, KS. Drafted the manuscript: AM, KS. Reviewed the manuscript: TF. Performed the experiment: AM. All authors reviewed and approved the final manuscript.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication, the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests.