The model organism Pyrococcus furiosus has recently been reported to interact with Methanopyrus kandleri in coculture, suggesting a H2 symbiosis. In the current study we further investigated this hypothesis by growing P. furiosus with four other hyperthermophilic methanogens providing evidence that the organisms did not only exert positive effects (P. furiosus/Methanocaldococcus villosus and P. furiosus/Methanocaldococcus infernus) on each other, but also neutral (P. furiosus/Methanocaldococcus jannaschii) and even inhibitory interactions (P. furiosus/Methanotorris igneus) were detected suggesting interspecies relationships not only based on H2 symbiosis. Using various microscopic techniques we further analyzed the coculture with the highest positive interactions (P. furiosus/M. villosus) concerning its growth behavior on various surfaces, which turned out to be in stark contrast to the previous reported coculture of P. furiosus/M. kandleri. This communication provides new insights into possible interactions of extremophilic Archaea in cocultures and again raises the question if and how hyperthermophilic Archaea communicate besides metabolic intermediates like H2.
The study of microbes living in communities was not in the focus of classical microbiology for a long period of time, although Winogradsky stated already more than 100 years ago that in pure cultures a very effective growth factor, namely competition with other Bacteria, is missing.1 Today it is evident that microbes prefer living in mixed species communities rather than growing isolated in pure cultures.2 Microorganisms often have strong dependencies on other organisms, relying for example on metabolic waste products making cultivation of particular organisms challenging. This might very likely be one of the reasons why only 1% of the estimated microbial bioburden can be cultivated under standard laboratory conditions.3,4
Only for a few Archaea interactions with other prokaryotes have been studied in detail.5 For instance, in the cold sulfidic water of the Sippenauer Moor (vicinity of Regensburg, Germany) a macroscopically visible microbial community forming a string-of-pearls-like structure has been discovered.6 Minute white pearls are connected to each other by thin white-colored threads. The interior part of the pearls is mainly composed of SM1 Euryarchaea, whereas Bacteria of the filamentous sulfide oxidizing genus Thiothrix are predominantly surrounding the Archaea. Due to the consistency of their appearance, a symbiotic relationship based on a sulfur cycle has been proposed for the microbes in the pearl.7
A highly specialized and intimate association between two hyperthermophilic Archaea is formed between Ignicoccus hospitalis, a member of the Crenarchaeota, and Nanoarchaeum equitans, the first representative of the novel phylum Nanoarchaeota.8,9N. equitans cells are diminutive cocci (diameter about 400 nm) and have a rudimentary genome lacking nearly all important genes for the biosynthesis of lipids, amino acids, nucleotides, and cofactors.10 They strongly depend on a close physiological contact to I. hospitalis, apparently because of the transport of metabolic substances, eg, membrane lipids and amino acids, into their cells.11 The fact that Ignicoccus cells are not harmed in this interaction rules out a classification as a typical host-parasitism system.
In marine and also in freshwater environments microbial communities between anaerobic methane oxidizing Archaea (ANME) and sulfate reducing Bacteria can be found.12,13 In these consortia the anaerobic oxidation of methane (AOM) is the exclusive energy source with sulfate acting as final electron acceptor. This leads to the fact that AOM consortia consume over 90% of the methane generated by the activity of marine microbes which corresponds to about 7%–25% of the total global methane production.14 The process of AOM therefore is of great ecological relevance in controlling global methane emission.
In habitats characterized by a high turnover of molecular hydrogen (H2), eg, in anaerobic high temperature habitats, the driving force for symbiotic relationships is an interspecies H2-transfer.15 This, in short, is the formation of methane by methanogenic Archaea using free H2 produced by one or more heterotrophic (micro)organisms. Stable associations based on H2 symbiosis have also been successfully established under laboratory conditions for example between Pelotomaculum thermopropionicum and Methanothermobacter thermoautotrophicus.16 In one of our previous studies the establishment of a stable coculture between the heterotrophic Archaeon P. furiosus and the methanogen Methanopyrus kandleri has been successfully demonstrated.17 In this intimate association, both species benefited from each other suggesting an interspecies H2-transfer. Furthermore, an extensive biofilm formation on different surfaces was observed, including the production of extracellular polymeric substances. In the current study we asked if P. furiosus might interact with other species. The results reported here prove that P. furiosus can also be grown in coculture with four other hyperthermophilic methanogens. Surprisingly, different types of interactions were present in these cocultures, reaching from beneficial, over neutral to negative. This communication provides new insights into possible interactions of extremophilic Archaea and emphasizes their complexity.
Materials and Methods
Strains and growth conditions
Four different species of the family Methanocaldococcaceae were chosen for the establishment of cocultures with P. furiosus (VC1, DSMZ 3638):18Methanocaldococcus jannaschii (JAL-1T, DMS 2661),19Methanocaldococcus infernus (MET, DSM 11812)20, Methanocaldococcus villosus (KIN24-T80T, DSM 22612)21 and Methanotorris igneus (Kol 5T, DSM 5666).22 MGG medium (standard methanogen medium with a ionic composition similar to sea salts)21 supplemented with 0.1% yeast extract and 0.25% peptone was used for cultivation of cocultures that were grown in 120 mL bottles containing 20 mL anaerobic medium at 85 °C with agitation. Growth was monitored by cell counting using a Thoma cell counting chamber (depth 0.02 mm) and a Nikon Microphot epi-fluorescence microscope. Phase contrast signals (all cells visualized) and auto-fluorescence signals (only methanogens visualized) were compared for differentiation of the species in a coculture. The cocultures were absolutely stable; they were transferred (1:1000 dilution) in weekly intervals over a period of 6 months, which means that they in principle were established 24 times consecutively. Although cell yields obtained by these weekly transfers differed by a maximum of 20% only, the data given in Table 1 and Fig. 1 were taken from a single experiment.
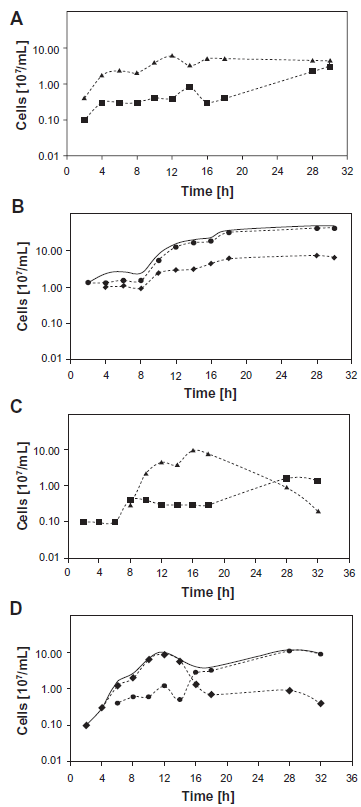
Growth curves of single cultures (A) and the cocultures of P. furiosus/M. villosus (B) and single cultures (C) and cocultures of P. furiosus/M. igneus (D).
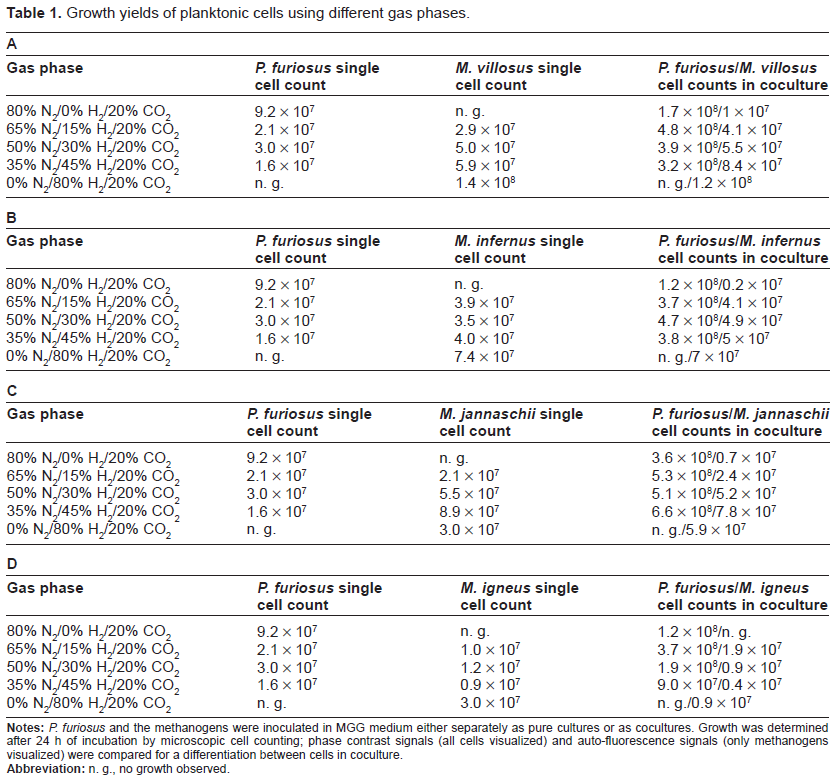
Growth yields of planktonic cells using different gas phases.
Notes:P. furiosus and the methanogens were inoculated in MGG medium either separately as pure cultures or as cocultures. Growth was determined after 24 h of incubation by microscopic cell counting; phase contrast signals (all cells visualized) and auto-fluorescence signals (only methanogens visualized) were compared for a differentiation between cells in coculture.
Abbreviation: n. g., no growth observed.
Generation of P. furiosus specific antibodies linked with fluorescent dye23
P. furiosus was cultured anaerobically in modified SME medium at 90 °C in 120 mL bottles.24 Cells were sheared with an Ultraturrax T25 (IKAWerke, Staufen, Germany) for 1 min at 13,000 rpm and for 10 s at 20,500 rpm in order to remove the cell-appendages. Afterwards cells were diluted to 1 × 107-1 × 108 cells/mL in fresh SME medium (without the addition of organic compounds). Immunization was performed by Davids Biotechnologie GmbH Regensburg. For initial immunization 1 mL of cell suspension was administered to a rabbit with two booster doses following after five and eight weeks of immunization. The antibodies were purified from serum by protein G affinity chromatography and their specificity was proven in western blot experiments by reaction with crude cell lysates. Afterwards the polyclonal antibodies were linked with AlexaFluor 555. Therefore 6–7 mg/mL of purified antibody solution were incubated with 100 µg of fluorescent dye for 1 h at room temperature under addition of 60 µL 1 M NaHCO3. Excess dye was removed by use of a Microcon CM 30 centrifugal filter unit.
Adherence studies using confocal laser scanning microscopy and scanning electron microscopy
Analyses of adherence to various surfaces were conducted as described earlier.17,21,25 Different materials were added to ½ SME medium which was inoculated and incubated at 85 °C for 20 h until stationary growth phase was surely reached.24 Gas phases were N2/CO2 for P. furiosus, H2/CO2 for M. villosus and N2/H2/CO2 (65/15/20, v/v/v) for the coculture. Thereafter materials were removed and washed with sterile medium. For all materials, with exception of gold grids, which were analyzed by transmission electron microscopy, an Olympus BX 50 epi-fluorescence microscope was used to detect the cells on the surfaces. The detection of the autofluorescence of M. villosus and the labeling of P. furiosus with surface specific antibodies linked with AlexaFluor 555 allowed a differentiation of the species on opaque materials. In case of light-transmissive materials phase contrast and auto-fluorescence signals were sufficient for differentiation.
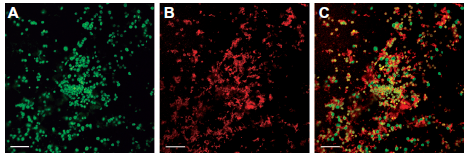
For confocal laser scanning microscopy adherent cells were analyzed after double fluorescent staining. First, P. furiosus cells were stained with an AlexaFluor 555 labeled cell surface antibody. In a second step the cells of both species were stained with AlexaFluor 488. AlexaFluor 555 was excited with a wavelength of 543 nm and AlexaFluor 488 with 488 nm. The different fluorescent signals allowed a differentiation between the species (green for AlexaFluor 488 = all cells, red for AlexaFluor 555 = P. furiosus; in the overlay P. furiosus cells appear yellow colored).
For scanning electron microscopy cells were incubated for 20 h with either mica, cover slips or glassy carbon added to the medium. “Flocks” consisting of cells unattached to a surface were directly picked from the medium using pasteur pipettes. Subsequently, cells were fixed using a solution containing 1 part glutardialdehyde (25% (w/v), EM-grade), 1 part HEPES-buffer (0.5 M) and 8 parts medium without organic. Further processing was as described earlier.25
Results and Discussion
Here we report on relationships between hyperthermophilic heterotrophs and methanogens established on a potential interspecies H2-transfer. Recently, research on the biological production and consumption of methane has undergone tremendous changes due to the increasing public interest in global warming. Beyond doubt, methane gas increases the greenhouse effect on Earth and consequently, methanogens are discussed to be significant contributors to climate change and global warming.26 It has been estimated that about 500–600 Tg methane is emitted into the atmosphere per year with about 74% deriving from microbial activities.12 Major sources of methane emission are anoxic environments, such as the guts of termites, the intestinal tract of ruminants and rice fields,12 all of them being mesophilic habitats of methanogens. Their interactions with other microorganisms require scientific attention and further analyses in order to understand their ecological role in depth. Here, we focused on possible interactions of methanogenic and heterotrophic Archaea under non-mesophilic but hyperthermophilic conditions in order to shed light onto the advantages and disadvantages methanogens may gain from interspecies relationships. For this reason we have chosen P. furiosus, a model organism for hyperthermophilic heterotrophic Archaea, and analyzed if it can grow in association with methanogenic species, in this case four species of the family Methanocaldococcaceae.
Establishing cocultures
The establishment of stable cocultures between these species–-probably based on an intermediate H2-transfer–-seemed plausible, because all of them exist in similar marine biotopes, characterized by high-temperatures and anaerobic conditions. Furthermore, a H2 symbiosis is likely to occur between P. furiosus and the methanogenic organisms, since P. furiosus is inhibited by H2,18 whilst the methanogens metabolize it for energy production. The data presented in Table 1 clearly show that P. furiosus as well as the methanogens could be grown separately to high cell densities in MGG medium if the ideal gas phases for the pure cultures were used: N2/CO2 (80:20 v/v) and H2/CO2 (80:20 v/v) for P. furiosus and the Methanocaldococci, respectively. After combining P. furiosus with one of the methanogens, cocultures could be successfully established in MGG medium in all cases. It has to be noted that the cell numbers of both organisms in coculture considerably fluctuated (up to 35% relative standard error). In spite of this fact the cocultures could be successfully transferred over a period of six months at weekly intervals, confirming their stability. The data provided in Table 1A–D show that a gas mixture containing N2 as well as H2 was necessary to allow growth of both species. Simultaneous growth of both organisms was impossible under a H2/CO2 gas phase and only possible to some extent under N2/CO2. Hence we concluded that P. furiosus does not produce sufficient H2 during its fermentation to support reasonable growth of its methanogenic partner under a N2/CO2 gas phase. On the other hand, the growth of P. furiosus is inhibited by the high amounts of H2 when providing only H2/CO2.
Analyses of the cocultures
Particularly in the coculture consisting of P. furiosus and M. villosus the influence of gas phase variation on growth yields was pronounced (Table 1A). An increase of the H2 fraction in the gas mixture lead to a decrease of the cell numbers of P. furiosus, while it stimulated the growth of the methanogenic partner. Similar observations also hold true for the coculture P. furiosus/M. infernus (Table 1B). Nevertheless, all of the different gas mixtures (except H2/CO2) lead to cocultures which had a stimulatory effect on both partners. Especially P. furious cells grew much higher in coculture compared to the corresponding pure culture, whilst for the methanogens a slightly positive effect was detected. These findings were supported by growth curve determinations, which clearly show the differences in growth yields after cultivation of the organisms either in coculture or in pure culture (see Fig. 1A and B for data of the P. furiosus/M. villosus coculture). By a comparison of phase contrast pictures (detecting both species) and auto-fluorescence signals (detecting the methanogenic species only), we proved that P. furiosus was the dominant species in the cocultures. This was not surprising, since we had already shown earlier that P. furiosus grows to higher cell numbers in coculture with M. kandleri.17 We conclude that in the case of the P. furiosus/M. villosus and the P. furiosus/M. infernus cocultures both species benefit from the presence of the partner and propose a mutualistic interspecies relationship relying on the transfer of intermediate H2. This type of interaction has been also described in the cases of a Bacterium and a methanogenic Archaeon (eg, P. thermopropionicum and M. thermoautotrophicus or Thermotoga maritima and M. jannaschii)16,27 and of two Archaea (P. furiosus and M. kandleri).17
Considering the other cocultures of P. furiosus and the methanogens, an unanticipated finding was that the partners did not always benefit from each other. In contrast to the aforementioned cocultures with positive interactions, cultivation of P. furiosus together with M. jannaschii revealed an advantage for the heterotrophic Archaeon only. In other words, we observed an increase in cell number for P. furiosus in the coculture, whereas the methanogenic organism just tolerated the presence of its “partner” (Table 1C).
Moreover, a negative effect was observed if P. furiosus was grown together with M. igneus. As shown in Table 1D, the growth of M. igneus was significantly lower in the coculture than in pure culture. Analysis of growth curves (Fig. 1C and D) revealed that M. igneus actually grew in coculture, but “died off” quickly (about 12 hrs after inoculation) before P. furiosus entered the exponential growth phase. In pure cultures M. igneus cell numbers declined only about 20 hrs after inoculation. P. furiosus and M. igneus never were observed to reach high cell numbers at the same time point in coculture and rather seem to “avoid each other”. Though these results indicated an inhibitory effect of both organisms on each other, this “coculture” could be transferred without any problems over prolonged time (6 months).
The importance of flagella for initial surface attachment has been proven for Sulfolobus solfataricus.28 For P. furiosus it has been reported that flagella are essential for the establishment of a permanent adhesion25 and for the formation of intra- and interspecies cell-cell contacts.17M. igneus was described to possess only a few flagella, a feature differentiating it from the three species of the genus Methanocaldococcus used in this study. Therefore a prerequisite for successful coculture establishment might be missing in this case. Consequently, the question rises on how these cells communicate. So far, the potential to produce quorum sensing signals like acyl-homo-serine lactones in Archaea has been described for Natronococcus occultus only.29 Because of the heat instability of these molecules they are not expected to be present in hyperthermophilic cultures and up to now no such signal molecules have been discovered for these organisms.29,17 In a study in which we have tested more than 800 archaeal isolates for the production of quorum sensing signals, not a single one was detected; neither in the culture supernatants–-using two different detection systems–-nor on the DNA level was there any indication for the existence of such signal molecules (own, unpublished data). It has to be mentioned for completeness that we did not examine the gasphase because up to now all known quorum sensing signals (like N-acyl-homoserine lactones or peptide autoinducers) are not gaseous. At the moment it remains unclear, how P. furiosus and M. igneus sense each other and avoid growing together in a coculture.
Adhesion of the P. furiosus/M. villosus coculture to surfaces
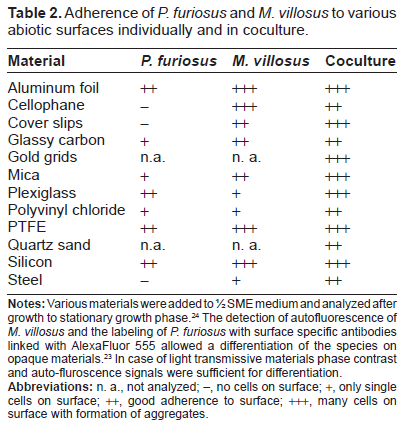
Recently we have reported that P. furiosus does use its flagella not only for swimming but also for adherence to different abiotic surfaces.25 Experiments revealed that P. furiosus could adhere to surfaces related to the natural habitat of the Archaeon (eg, sand grains), which seems reasonable for the microorganism to stay under favorable growth conditions. Additionally the P. furious cells adhered onto various surfaces, which in the case of metals and plastics have been chosen because of a potential biotechnological application. We had also demonstrated that in a hyperthermophilic coculture, P. furiosus can interact with cells of its partner M. kandleri, resulting in the first structured archaeal bi-species biofilm between cultured Archaea.17 This is in contrast to biofilm formation in three closely related hyperthermophilic Crenarchaeota: Sulfolobus acidocaldarius, S. solfataricus and S. tokodaii.30 Here it has clearly been shown that different biofilm architectures were built on solid surfaces ranging from carpet-like structures in S. solfataricus to high density tower-like structures in S. acidocaldarius. Biofilm formation in extremophilic Archaea seems to be a result of adaptation to extreme conditions, to high temperatures in the case of P. furiosus. This awoke our interest in a potential biofilm-forming style in the P. furiosus/M. villosus coculture, which additionally might be due to the optimal transfer of molecular hydrogen. Therefore, the growth on a variety of materials was analyzed in comparison to surface growth in pure culture. Table 2 provides an overview of the results of these adhesion studies. The fact that P. furiosus is not able to adhere to glass surfaces had already been shown previously and was confirmed here.17,25M. villosus was described to attach to glass cover slips21 and somehow seems to prime the surface for adhesion of P. furiosus, since in coculture both species were observed to attach to this material. Synthetic polymers like PTFE and silicon allowed the adherence of both species in pure culture as well as in coculture. Also, gold grids, plexiglass and aluminum foil were suitable for adhesion of the coculture. On mica, the majority of the cells were spread evenly over the surface; in addition, several aggregates could be detected by using various microscopic techniques.
Adherence of P. furiosus and M. villosus to various abiotic surfaces individually and in coculture.
Material
P. furiosus
M. villosus
Coculture
Aluminum foil
++
+++
+++
Cellophane
–
+++
++
Cover slips
–
++
+++
Glassy carbon
+
++
++
Gold grids
n.a.
n. a.
+++
Mica
+
++
+++
Plexiglass
++
+
+++
Polyvinyl chloride
+
+
++
PTFE
++
+++
+++
Quartz sand
n.a.
n. a.
++
Silicon
++
+++
+++
Steel
–
+
++
Notes: Various materials were added to ½ SME medium and analyzed after growth to stationary growth phase.24 The detection of autofluorescence of M. villosus and the labeling of P. furiosus with surface specific antibodies linked with AlexaFluor 555 allowed a differentiation of the species on opaque materials.23 In case of light transmissive materials phase contrast and auto-fluroscence signals were sufficient for differentiation.
Abbreviations: n. a., not analyzed; –, no cells on surface; +, only single cells on surface; ++, good adherence to surface; +++, many cells on surface with formation of aggregates.
To study the three dimensional aggregates on mica, samples with adherent cells were examined via confocal laser scanning microscopy. A microcolony scanned in the x, y and z directions revealed a three-dimensional structure with a height of 10.5 µm. Different fluorescent signals allowed a differentiation of the two coccoid Archaea (see Fig. 2). Both species were present in this microcolony and were associated very closely to each other in a random distribution–-this is in stark contrast to the P. furiosus/M. kandleri coculture growing in a defined structure.17 In order to further examine the type of interaction between the cells, scanning electron microscopy (SEM) was performed. A differentiation of both species was possible, since M. villosus cells are somewhat bigger than P. furiosus cells and characterized by an “outstanding pattern of the cell surface” (compare Fig. 3C and D).21 On most of the examined materials, the cells were spread evenly over the entire surface rather than forming large aggregates. It became obvious that the cells interacted extensively via their flagella. On certain materials like mica and cover slips, flagella formed a dense network (see Fig. 3A and B). Interestingly, during early stages of establishing the coculture of both Archaea, macroscopically visible, whitish “flocks” appeared in the liquid medium. SEM analyses of those aggregates revealed that they consist of P. furiosus and M. villosus cells connected via their flagella even without the presence of a solid surface (see Fig. 3D). The data presented in Figure 3 clearly show that in the coculture between P. furiosus and M. villosus cells are in close contact via their flagella, which might be a prerequisite for optimal H2 exchange. Whether both species contribute to these interactions, or only the flagella of one species are involved needs to be analyzed further. In the case of a coculture consisting of the Bacterium P. thermopropionicum and the Archaeon M. thermoautotrophicus a cell-cell communication via flagella has been reported.31 In this coculture aggregates were formed and stabilized by the flagella of the Bacterium. It turned out that the tip protein of the flagella provoked a change in gene expression in the Archaeon, resulting in higher expression of genes responsible for methanogenesis. It has to remain open, if such a situation–-signaling and interaction between both partners via their flagella–-also occurs for the P. furiosus/M. villosus system studied here. We assume this to be very likely, because the flagella of both P. furiosus and M. villosus have been reported to mediate adhesion of cells to abiotic surfaces (including sand grains from the original biotope) and between each other.25,21 Studies asking if P. furiosus and M. villosus will grow in close association and interact in hyperthermophilic, anaerobic marine biotopes via their flagella are therefore warranted.
Laser scanning micrograph of the coculture P. furiosus/M. villosus on mica.
Scanning electron micrograph (SEM) of the coculture P. furiosus/ M. villosus on different surfaces and in liquid phase.
Conclusions
Together with the earlier established coculture P. furiosus/M. kandleri a total of five cocultures of the heterotrophic Archaeon P. furiosus with methanogenic Archaea have now been analyzed with respect to the interactions of the partners involved. Of these, one turned out to be of a negative kind (P. furiosus/M. igneus), and one to be neutral (P. furiosus/M. jannaschii). In three cocultures positive effects on both partners have been observed, ie, they are of a symbiotic kind for P. furiosus/M. kandleri, P. furiosus/M. villosus and P. furiosus/M. infernus. A notable observation is that the P. furiosus/M. kandleri and P. furiosus/M. villosus systems differ by the development of structured biofilms (former coculture) versus unstructured biofilms (latter coculture).
These findings demonstrate very nicely the enormous breadth of possible interactions between two different microorganisms. We take this to indicate that predictions of potential interactions are difficult at the very last, and only experimentation will reliably show which of the various kinds of possible interactions nature prefers.
Disclosures
This manuscript has been read and approved by all authors. This paper is unique and not under consideration by any other publication and has not been published elsewhere. The authors and peer reviewers report no conflicts of interest. The authors confirm that they have permission to reproduce any copyrighted material.
Footnotes
Acknowledgements
We gratefully acknowledge the expert technical help by Y. Bilek and S. Dobner and thank M. Cooper for critically reading the manuscript. This work was supported by DFG grant WI 731/10-1 to RW.
References
1.
ZavarzinG.Winogradsky and modern microbiology. Microbiol.2006; 75(5): 501–11.
2.
FlemmingH-C, WingenderJ.Biofilme—die bevorzugte Lebensform der Bakterien: Flocken, Filme und Schlämme. Biologie in unserer Zeit.2001; 31(3): 169–80.
3.
AmannR., LudwigW., SchleiferK.Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev.1995; 59(1): 143–69.
Moissl-EichingerC., HuberH.Archaeal symbionts and parasites. Curr Opin Microbiol.2011; 14(3): 364–70.
6.
RudolphC., WannerG., HuberR.Natural communities of novel Archaea and Bacteria growing in cold sulfurous springs with a string-of-pearls-like morphology. Appl Environ Microbiol.2001; 67(5): 2336–44.
7.
MoisslC., RudolphC., HuberR.Natural communities of novel Archaea and Bacteria with a string-of-pearls-like morphology: molecular analysis of the bacterial partners. Appl Environ Microbiol.2002; 68(2): 933–7.
8.
HuberH., HohnM.J., RachelR., FuchsT., WimmerV.C., StetterK.O.A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont. Nature.2002; 417(6884): 63–7.
9.
PaperW., JahnU., HohnM.J.Ignicoccus hospitalis sp. nov., the host of ’ Nanoarchaeum equitans’. Int J Syst Evol Microbiol.2007; 57(4): 803–8.
10.
WatersE., HohnM.J., AhelI.The genome of Nanoarchaeum equitans: Insights into early archaeal evolution and derived parasitism. PNAS.2003; 100(22): 12984–8.
11.
JahnU., SummonsR., SturtH., GrosjeanE., HuberH.Composition of the lipids of Nanoarchaeum equitans and their origin from its host Ignicoccus sp. strain KIN4/I. Arch Microbiol.2004; 182(5): 404–13.
12.
LiuY., WhitmanW.B.Metabolic, phylogenetic, and ecological diversity of the methanogenic Archaea. Ann NY Acad Sci.2008; 1125(1): 171–89.
13.
KnittelK., BoetiusA.Anaerobic oxidation of methane: progress with an unknown process. Annu Rev Microbiol.2009; 63(1): 311–34.
14.
HinrichsK.U., BoetiusA.The anaerobic oxidation of methane: new insights in microbial ecology and biogeochemistry. In: WeferG., BillettD., HebbelnD., JørgensenB., SchlüterM., van WeeringT., editors. Ocean Margin Systems.Berlin: Springer-Verlag; 2002: 457–77.
15.
AdamsM.W.W.The metabolism of hydrogen by extremely thermophilic, sulfur-dependent bacteria. FEMS Microbiol Lett.1990; 75(2-3): 219–37.
16.
IshiiS., KosakaT., HoriK., HottaY., WatanabeK.Coaggregation facilitates interspecies hydrogen transfer between Pelotomaculum thermopropionicum and Methanothermobacter thermautotrophicus. Appl Environ Microbiol.2005; 71(12): 7838–45.
17.
SchopfS., WannerG., RachelR., WirthR.An archaeal bi-species biofilm formed by Pyrococcus furiosus and Methanopyrus kandleri. Arch Microbiol.2008; 190(3): 371–7.
18.
FialaG., StetterK.O.Pyrococcus furiosus sp. nov. represents a novel genus of marine heterotrophic Archaebacteria growing optimally at 100 °C. Arch Microbiol.1986; 145(1): 56–61.
19.
JonesW.J., LeighJ.A., MayerF., WoeseC.R., WolfeR.S.Methanococcus jannaschii sp. nov., an extremely thermophilic methanogen from a submarine hydrothermal vent. Arch Microbiol.1983; 136(4): 254–61.
20.
JeanthonC., L'HaridonS., ReysenbachA.L.Methanococcus infernus sp. nov., a novel hyperthermophilic lithotrophic methanogen isolated from a deep-sea hydrothermal vent. Int J Syst Evol Microbiol.1998; 48(3): 913–9.
21.
BellackA., HuberH., RachelR., WannerG., WirthR.Methanocaldococcus villosus sp. nov., a heavily flagellated archaeon that adheres to surfaces and forms cell–cell contacts. Int J Syst Evol Microbiol.2011; 61(6): 1239–45.
22.
BurggrafS., FrickeH., NeunerA.Methanococcus igneus sp. nov., a novel hyperthermophilic methanogen from a shallow submarine hydrothermal system. Syst Appl Microbiol.1990; 13: 263–9.
23.
SchopfS.Untersuchungen der Flagellen von Pyrococcus furiosus: Flagellenvermittelte Interaktionen, Ultrastruktur und Molekularbiologie. PhD thesis.2011: 58.
24.
StetterK., KoenigH., StackebrandtE.Pyrodictium gen.nov., a new genus of submarine disc-shaped sulphur reducing Archaebacteria growing optimally at 105 °C. Syst Appl Microbiol.1983; 4(4): 535–51.
25.
NaetherD.J., RachelR., WannerG., WirthR.Flagella of Pyrococcus furiosus: multifunctional organelles, made for swimming, adhesion to various surfaces, and cell-cell contacts. J Bacteriol.2006; 188(19): 6915–23.
26.
SinghB.K., BardgettR.D., SmithP., ReayD.S.Microorganisms and climate change: terrestrial feedbacks and mitigation options. Nat Rev Micro.2010; 8(11): 779–90.
27.
MuralidharanV., RinkerK.D., HirshI.S., BouwerE.J., KellyR.M.Hydrogen transfer between methanogens and fermentative heterotrophs in hyperthermophilic cocultures. Biotechnol Bioeng.1997; 56(3): 268–78.
PaggiR.A., MartoneC.B., FuquaC., De CastroR.E.Detection of quorum sensing signals in the haloalkaliphilic archaeon Natronococcus occultus. FEMS Microbiol Lett.2003; 221(1): 49–52.