
Abstract
Small colony variant (SCV) bacteria arise spontaneously within apparently homogeneous microbial populations, largely in response to environmental stresses, such as antimicrobial treatment. They display unique phenotypic characteristics conferred in part by heritable genetic changes. Characteristically slow growing, SCVs comprise a minor proportion of the population from which they arise but persist by virtue of their inherent resilience and host adaptability. Consequently, SCVs are problematic in chronic infection, where antimicrobial treatment is administered during the acute phase of infection but fails to eradicate SCVs, which remain within the host causing recurrent or chronic infection. This review discusses some of the phenotypic and genotypic changes that enable SCVs to successfully proliferate within the host environment as potential pathogens and strategies that could ameliorate the resolution of infection where SCVs are present.
Discovery of Small Colony Variants (SCVs)
Pure bacterial cultures are not genetically homogeneous, and their behavior is determined by genomic characteristics, such as a high degree of plasticity. Slow-growing subpopulations of bacteria in pure culture have been described from as early as 1913; reported to emerge in response to diverse environmental pressures, they were termed SCVs because they formed pin-prick-sized colonies when cultured on solid media.1,2 Initially, SCVs were thought of as morphological variants with a secondary role in infectious disease because of their markedly diminished pathogenicity and impaired production of virulence factors.
2
Furthermore, it was believed that the
Early studies clarified the link between environmental stress and the phenotypic changes that became associated with SCVs, including atypical colony morphology, slow growth rate, lack of pigmentation, reduced hemolytic activity, reduced coagulase activity, reduced carbohydrate utilization, low virulence potential, and elevated antibiotic resistance (Fig. 1).7–9 Indeed, the growth rate of SCVs has been estimated to be approximately nine times slower than that of the progenitor organisms. 10 As such, SCVs are now better defined as a microbial subpopulation constituting a naturally occurring, slow-growing but diverse bacterial morphotype.7,11 Clinically, this is problematic; the presence of SCVs during infection is correlated with recurrent or chronic infectious disease. A combination of extended incubation time in addition to altered phenotypic and biochemical traits often means that SCVs in patient samples are overlooked by clinical microbiologists utilizing conventional diagnostic tests. This results in the cessation of antimicrobial treatment before SCVs are effectively cleared from an infection; therefore, they persist causing recurrent and chronic infection.7,12
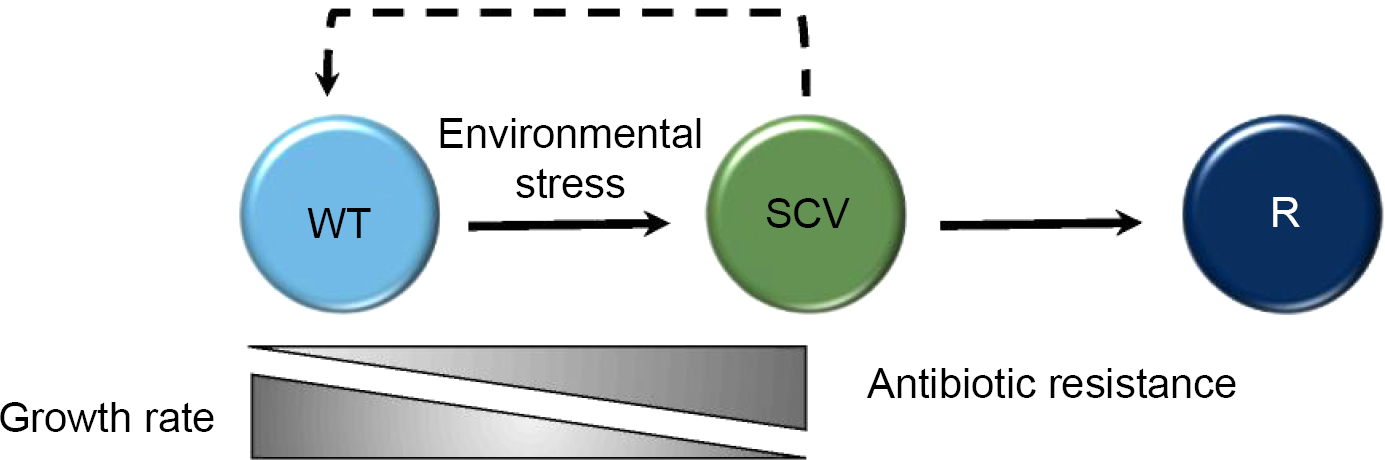
WT organisms undergo a shift to the SCV phenotype under the conditions of stress, where they exhibit a slower growth rate but increased antibiotic resistance. they can revert to a WT-like (indicated by a dashed line to denote that WT-like organisms are not identical to the original WT progenitor) or alternative revertant (R) phenotype when the environmental stress is removed, regaining a faster growth rate but becoming more susceptible to antibiotic treatment.
Various environmental stimuli appear to result in phenotypically distinct varieties of SCV.7,13 Some undergo permanent genetic changes, whereas a subpopulation reverts to a wild-type (WT) phenotype or to a different phenotype that is distinct from both the progenitor and the SCV upon repeated subculture (revertant phenotype) (Fig. 1).
14
Phenotypic reversion, where genetic mutations have not occurred, happens rapidly and circumvents any permanent fitness costs.
14
The tendency to permanent genetic alteration as compared to phenotypic reversion seems to depend largely on the nature of the original environmental pressure.
7
There is not always commonality between phenotypic and genotypic changes within different SCV populations, but there are a number of prevalent auxotrophies, characterized initially in
Unique Phenotypic Traits Associated with SCVs
In addition to the auxotrophies described above, there are a number of other phenotypic characteristics typically associated with SCVs that likely contribute to their ability to persist under adverse growth conditions. One such conventional SCV attribute is diminished electron transport, observed in various species of
Characteristics associated with SCVs.

The unique cell wall structure of SCVs is believed to confer some degree of protection from stress and is allied with aberrant electron transport. Abnormal cell division has been described for SCVs of
Characteristically, in addition to the aforementioned phenotypic changes, the small regulatory RNA molecule, RNAIII, is usually absent from SCVs and has been particularly well defined for SCVs of
Once attached to the host cell, SCVs, like their WT counterparts, induce host-cell changes by actin rearrangement, which mediates internalization, effectively hijacking non-phagocytic cells (including endothelial cells, fibroblasts, osteoblasts, and keratinocytes).41,42 Pathogens that are not categorized as SCVs utilize the same mechanism of internalization, but crucially, SCVs are far more efficient at this process than their progenitors.
43
Fundamentally, this intracellular protection affords additional defense against immune clearance or antimicrobial treatment. Owing to their reduced toxicity, the uptake of SCVs in this manner occurs in vitro without damage to host cells.
44
Once inside the host cell, intracellular survival is critical to retain protection.
40
SCVs characteristically proliferate intracellularly, more successfully than their progenitors, which is a trait that directly contributes to antibiotic treatment failure and poor prognosis in patients.45–47 A marked increase in the expression of member genes of the arginine deaminase pathways in SCVs of
In addition, SCVs evade the immune response and persist intracellularly by escaping from intracellular phagosomes,43,48 thus avoiding the hydrolytic activity of lysosomes. 48 It has been proposed that unlike other intracellular pathogens, once in the cytoplasm, SCVs may no longer disrupt normal actin polymerization of the cells in which they reside, meaning that they do not elicit normal intracellular cytokine and chemokine defense mechanisms. 40 Therefore, the ability of SCVs to dampen the proinflammatory response means that the attenuated virulence associated with the SCV phenotype is in fact favorable for their survival and prolonged persistence within the host. 46 The recovery of SCVs from the cases of asymptomatic infection supports this theory of persistence through diminished host damage.48,49
Unique Genotypic Features Associated with SCVs
Several genetic mutations can result in the electron transport-defective SCV phenotype described above, including the mutations in
During infection, organisms are exposed to high levels of haem that may be toxic because of the accumulation of super-oxides. By virtue of the mutations described above, haem stress for SCVs is significantly alleviated, suggesting that a reduction in haem-associated stress may be an additional factor enabling the survival of SCVs during chronic infection. 54 Genes governing other aspects of general metabolic pathways associated with energy production and respiration in SCVs often also carry mutations (Table 1). They primarily include genes encoding proteins of the Entner–Doudoroff pathway, reconciling the slower growth rate of SCVs that contributes to their persistence.
Increased adhesion and biofilm formation are correlated with the enhanced expression of surface-bound adhesins and their cognate transcriptional regulators.55–58 Adhesins not only function as a means of binding directly to host proteins prior to colonization but also enable interbacterial aggregation, which is critical to the development of biofilm; SCVs are characteristically prolific biofilm-forming organisms.
59
The expression of adhesin genes is often governed by global transcriptional regulators that form part of an intricate transcriptional network that responds to environmental cues, usually involving quorum sensing. Therefore, it is not surprising to find that genes encoding transcriptional regulators, such as
Although differentially expressed or mutated genes tend to be conserved in SCVs, to date no defined core set of SCV genes has been documented. Often SCV-associated phenotypic traits are not the result of permanent genetic mutations but may instead result from genome rearrangements; therefore, identifying SCVs at the genotypic level is potentially as challenging as identifying them based on phenotype alone. Moreover, numerous phenotypic traits can be attributed to epigenetics.
67
Where genetic traits are conserved, they usually confer essential adaptations; transient characteristics that are not an absolute requirement for survival, but which confer a competitive advantage, are likely controlled by uncharacterized global transcriptional regulators or alternative sigma factors (Table 1) that form part of a larger and as yet undefined
SCVs within Microbial Communities
Variants occur at random within microbial populations; most are transitory with only those changes that allow bacteria to remain viable and confer an advantage becoming fixed within a population. Microbial adaptation to a particular environment and competition between the members of a heterogeneous population, comprising a parent (wild type) and progeny (including mutants), are dictated by growth parameters and stresses. 69 Numerous laboratory studies have demonstrated that successful microorganisms, namely, those that succeed within a given environment, do so because they exhibit the highest growth rate under prevailing conditions. 70 Despite this, SCVs persist within microbial communities, albeit as a minor constituent that is never entirely outcompeted by the parental strain. Given the tenacity of SCVs to survive under stress, it might be expected that they should eventually predominate. Certainly with regard to antimicrobial interventions, 71 this is the case, as the more susceptible parent strain is eradicated leaving behind a population of SCVs that can undergo reversion, which results in a recurrence of infection (Fig. 2). 72 SCVs that arise during human infection appear better adapted for survival and persistence within the host 73 despite their impaired rate of growth and increased host dependency. Selection because of loss or redundancy of metabolic activity is not unusual and under certain growth conditions, 74 such as the host environment, might confer a fitness advantage, if not in terms of growth, in terms of survival. 75 Many host-adapted and therefore invariably host-dependent pathogens undergo reductive evolution; 76 driven by the host habitat and their ability to utilize host metabolites, such organisms become slow growing and nutritionally fastidious, often adopting an intracellular lifestyle. 77 This process is ordinarily mediated by the loss of large fragments of genomic DNA. SCVs appear similar to host-adapted pathogens, and the loss of metabolic function renders them reliant on the host to meet their nutritional needs; 78 indeed, many SCVs survive intracellularly. Where reductive evolution occurs in pathogens, it is correlated with increased virulence 79 that is not observed for SCVs, which conversely seem to exhibit attenuated virulence. 80 Significantly, pathogens that undergo reductive evolution do not regain “lost” genetic function 81 unlike SCVs that are able to revert to WT or WT like once selective pressures, such as antimicrobial treatment, are removed (Figs. 1 and 2). Therefore, for SCVs, there is apparently a trade-off between virulence and persistence that exploits the ability to revert to a WT or WT-like variant, which is less host dependent and regains virulence. The specific adaptations that the minority SCV population depends on to ensure survival among their faster growing counterparts include the increased expression of surface adhesins as previously described. If two microorganisms are competing for the same human receptor, then those with a binding advantage (ie, more surface adhesins) are more likely to adhere.82,83 Combined with enhanced biofilm formation, such colonizers are less likely to be removed from the host by detachment. 84 Intracellular survival provides more than simple protection from immunity, with the cell cytoplasm providing a nutritionally rich habitat for auxotrophic SCVs that is not afforded to the parental strain, but at the same time reduces competition for space at the tissue surface. 85 It is proposed that in this way, both SCV and progenitor can coexist.
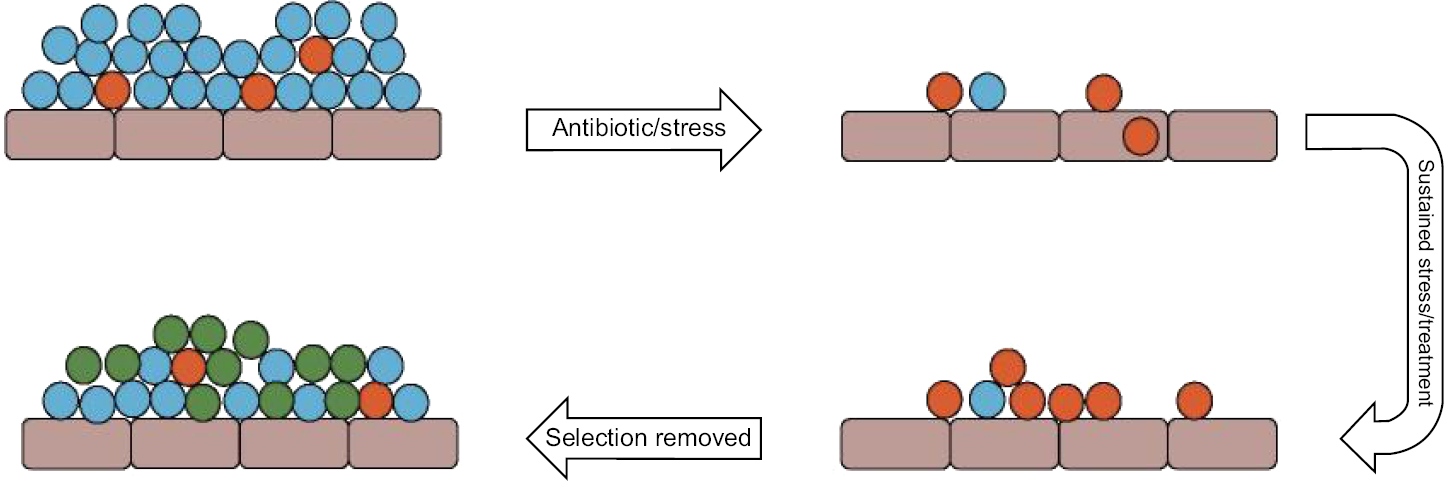
Pure populations of bacteria often comprise WT (major population; blue) and SCV (minor population; orange), which arise spontaneously; under environmental stress, such as antibiotic treatment, the WT population is diminished and SCVs survive; under sustained stress, such as a course of antibiotics to treat an infection, the SCVs become the dominant members of the population. When the selective pressure is removed, WT organisms proliferate and become the dominant members of the population compared to slow-growing SCVs; significantly a proportion of SCVs revert to either the WT phenotype or a WT-like phenotype (green), which regains characteristics that enable faster growth.
SCVs in Chronic Infection
SCVs show enhanced resistance to a range of antibiotics7,25,86–88 and have been directly associated with persistent infections in a number of diseases, including, but not limited to, cystic fibrosis,89,90 chronic obstructive pulmonary disease,
91
diabetic foot ulcers,
92
chronic rhinosinusitis,
93
chronic wound infections,94–96 systemic infections,
97
and infections arising from surgical intervention or medical devices,98–102 which can lead to serious and sometimes fatal clinical consequences, such as endocarditis, bacteremia, or meningitis.97,102 It has been proposed that in the case of the ventriculoperitoneal shunt (a medical device used in the treatment of hydrocephalus), SCV-associated meningitis infection can arise from inadequate disinfection of the shunt and failure to identify and treat these persistent variants.
103
Chronic infections represent a significant burden to both patients and health-care providers. Where chronicity ensues, biofilm is frequently present.104–108 The presence of SCVs within biofilms has been directly linked to chronic antibiotic-resistant infections, including cystic fibrosis in lung, osteomyelitis, catheter and pacemaker infection, among others as previously described.87–93,108–115 The respiratory tract of cystic fibrosis patients provides a unique environment for the selection of a subgroup of autoaggregative and highly adherent SCVs of
The rate of occurrence of SCVs in chronic infection is likely to vary depending on the clinical conditions;
7
nonetheless, SCVs are detected in approximately 1% of isolates in a clinical microbiology laboratory and their incidence is the highest in cystic fibrosis and osteomyelitis.
7
It is pertinent to highlight that in patients with osteomyelitis, surgical placement of slow-release gentamicin beads along with debridement is a common practice for treatment and may be linked to SCV induction.
120
This is of concern as inadvertent iatrogenic-induced SCVs may be formed as a result of the long-term exposure to gentamicin; studies have verified that SCVs can be recovered from patients undergoing treatment with gentamicin beads.
120
It has consequently been suggested that routine screening for SCVs should take place for patients treated with gentamicin beads for osteomyelitis.
28
Furthermore, given the recalcitrance of SCV-associated infection, it might seem reasonable to screen persons who are predisposed to developing chronic infection following the completion of antimicrobial chemotherapy. Therefore, with regard to efficacious antimicrobial treatments, the identification of SCVs is as important as ensuring an appropriate dose of antimicrobial is administered. However, this approach is confounded by the relatively limited information describing successful treatment of SCV infections. Since aminoglycosides are known to promote the emergence of SCVs in some bacterial species, including
Future Perspectives
Although SCVs have been known to exist for over century, little attention was originally given to them as they were believed to be nonvirulent and therefore not clinically important. However, as more is understood about their role in persistent infections, it has become imperative that mechanisms of SCV persistence and resistance, as well as population dynamics, are thoroughly explored. Recent investigations have proposed a low-cost point-of-care test for the diagnosis of
Footnotes
Author Contributions
Conceived the concepts: BEJ, KJP, NPT, SEM. Analyzed the data: BEJ, KJP, NPT, SEM. Wrote the first draft of the manuscript: BEJ, KJP, NPT, SEM. Contributed to the writing of the manuscript: BEJ, KJP, NPT, SEM. Agree with manuscript results and conclusions: BEJ, KJP, NPT, SEM. Jointly developed the structure and arguments for the paper: BEJ, KJP, NPT, SEM. Made critical revisions and approved final version: BEJ, KJP, NPT, SEM. All authors reviewed and approved of the final manuscript.