
Abstract
Representatives of the Staphylococcus genus are the most common pathogens found in hospital environments, and they are etiological agents for a large variety of infections. Various virulence factors are responsible for the symptoms and severity of infections caused by Staphylococcus aureus. Among them are staphylococcal enterotoxins (SEs), which cause staphylococcal food poisoning, and toxic shock syndrome toxin-1 (TSST-1). Some reports indicate that TSST-1 and staphylococcal enterotoxins are also produced by coagulase-negative staphylococci (CNS). The present review aimed to discuss general aspects of staphylococcal toxins as well as the epidemiology, genetics and detection of toxins in Staphylococcus aureus and coagulase-negative staphylococci, since these microorganisms are becoming more and more frequent in nosocomial infections.
Staphylococcus
Bacteria of the Staphylococcus genus comprise various species and subspecies that are widely distributed in nature and found mostly in the skin and mucous membranes of birds and mammals. They are formed by Gram-positive cocci which can be isolated, paired and most often aggregated, forming unmoving grapelike structures. They do not produce spores and most of the species are facultative anaerobic organisms [1].
They can be divided into two groups according to production of coagulase enzyme, which is capable of coagulating blood plasma. The synthesis of this enzyme is restricted to some species in the genus, among which S. aureus, S. schleiferi subsps. coagulans, S. intermedius, S. hyicus and S. delphini can be distinguished. The other staphylococci that do not synthesize coagulase are referred to as coagulase-negative staphylococci (CNS). Among them are S. epidermidis, S. haemolyticus, S. saprophyticus, S. cohnii, S. xylosus, S. capitis, S. warneri, S. hominis, S. simulans, S. saccharolyticus, S. auricularis, S. caprae, S. lugdunensis and S. schleiferi subsps. schleiferi [1,2].
Staphylococci usually maintain a benign or symbiotic relationship with their hosts; however, if the cutaneous barrier is broken by trauma or the presence of foreign bodies, these organisms may reach other tissues and proliferate, thus developing a pathogenic behavior [3].
Representatives of the Staphylococcus genus are the most common pathogens in hospital environments, and they are the etiological agents of a wide variety of infections. Hospital-acquired infections have been the major cause of morbidity and mortality as they are frequently acute and pyrogenic, and if untreated, they may cause bacteremia involving various organs. In the Staphylococcus genus, S. aureus has always been the most significant species in relation to hospital-acquired infections and continues to be the primary cause of community-acquired staphylococcal infections in most countries [4]. The most common infections include furuncles, cellulitis, impetigo and surgical wound sites. Some of the most severe infections are bacteremia, pneumonia, osteomyelitis, acute endocarditis, meningitis, scalded skin syndrome and muscle abscess as well as infection in the genitourinary tract, central nervous system and various intra-abdominal organs [5].
Prior to the 1970s, there were few reports of infection caused by CNS acknowledged by clinical professionals and microbiologists as contaminants in biological samples [6]. At present, they are considered essentially opportunistic microorganisms that prevail in numerous organic situations in order to produce serious infection [1,6]. The emergence of CNS as etiologic agents may also be due to the recognition of this group of organisms as opportunistic pathogens and to the increasing use of invasive procedures, such as intravascular catheters and prostheses in immunodepressed patients and those undergoing intensive treatment, preterm children, patients with neoplasia or those who have undergone transplantation [6]. Data obtained by SCOPE (Surveillance and Control of Pathogens of Epidemiologic Importance) from 1995 to 1998 have shown CNS as the most frequent etiologic agents in hospital-acquired bacteremias in the USA [7].
Staphylococcal Toxins
Various virulence factors are responsible for the symptoms and severity of infections caused by S. aureus. Such factors include hemolysins alpha(α), beta(β), gamma(γ), and delta(δ); leukocidin; exfoliative toxins A and B; toxic shock syndrome toxin-1 (TSST-1) and the staphylococcal enterotoxins (SE) [2].
Staphylococcal toxins can be divided into two groups according to their capacity for lysing cells: hemolysins or cytotoxins, which are capable of producing lesion directly to the outer membrane of target cells [8], and the so-called superantigenic toxins, which do not present direct lytic action, but can produce lesions through the overproduction of cytotoxins from activated T-cells and from monocytes/macrophages [9].
Hemolysins and leukocidin
S. aureus produces five different membrane-damaging toxins, four hemolysins (alpha-, beta-, gamma- and delta-hemolysin) and leukocidin. Alpha-toxin is a pore-forming hemolytic toxin that causes membrane damage to many types of mammalian cells [10]. In particular, rabbit erythrocytes are extraordinarily susceptible to hemolysis by alpha-toxin, at least 100 times more so than other mammals and 1,000 times more than human erythrocytes [10]. The alpha-hemolysin is dermonecrotic and neurotoxic and can be lethal in a variety of animal systems. These factors contribute to the consensus among researchers that this toxin is important in the formation of abscesses characteristic of infections caused by S. aureus. The gene that encodes alpha-hemolysin, hla, was cloned from the S. aureus chromosome and sequenced in 1984 by Gray and Kehoe [11].
S. aureus beta-hemolysin was also shown by these authors to be highly hemolytic for sheep but not rabbit erythrocytes. Beta-toxin is Mg2+-dependent sphingomyelinase C, which degrades sphingomylin in the outer phospholipid layer of the erythrocyte membrane. The hemolytic activity of this toxin was enhanced by incubation below 10 °C after treatment at 37 °C, and is thus known as “hot-cold” hemolysin. The gene, named hlb, is chromosomally located and encodes a 330-amino-acid polypeptide with a predicted molecular weight of 39,000 daltons [12].
Delta-hemolysin, a 26-amino-acid peptide, is capable of causing membrane damage in a variety of mammalian cells. This toxin presents a detergent action on cell membranes resulting in cell lysis. Among the cytotoxins produced by Staphylococcus, delta toxin is characterized by its thermostability, neutralization by lectin, and synergism with beta toxin [13]. The delta toxin may play an important role in the pathogenesis of intestinal diseases and can vary from an acute diarrhea to a severe enteritis [14]. In S. aureus, the hld gene responsible for delta toxin production is situated within the RNAIII locus of the accessory gene regulator (agr gene), which controls the expression of most exoproteins of S. aureus. The hld gene of S. aureus encodes a 44-amino-acid peptide [15]. The same activity is mediated in S. epidermidis by a peptide highly homologous to S. aureus delta toxin, and whose gene is also located within the RNAIII locus of the agr gene [16], but the hld gene of S. epidermidis encodes a 26-amino-acid peptide that differs in only three amino acids from the hld gene of S. aureus [15,16].
Gamma-hemolysin is additionally able to lyse many varieties of mammalian erythrocytes. Gamma-hemolysin is not identifiable on blood agar plates, however, due to the inhibitory effect of agar on toxin activity [17]. These toxins are characterized by two nonassociated secreted proteins, referred to as S and F components (for slow- and fast-eluting proteins in an ion-exchange column). The gamma-toxin locus expresses two class S components (HlgA and HlgC) and one class F component (HlgB). Thus, the Hlg locus can express two functional pairs of proteins, HlgA+HlgB and HlgC+HlgB, both of which display proinflammatory effects when injected into the rabbit eye vitreous humor [18]. Gamma-toxin has also been proposed as playing a role in the pathogenesis of toxic shock syndrome (TSS) together with toxic shock syndrome toxin-1 (TSST-1), since this hemolysin is very frequently found in TSS isolates [19]. Many attempts have been made to understand which components of S. aureus are important to the development and persistence of infection.
Panton-Valentine leukocidin [PVL] is a pore-forming cytotoxin that targets human and rabbit mononuclear and polymorphonuclear cells [20]. Studies have shown that its toxic effect results from the synergistic action of two separate exoproteins, namely, LukS-PV and LukF-PV. These proteins are encoded by two contiguous and cotranscribed genes (lukS-PV and lukF-PV) [21], which are carried on temperate bacteriophages [22]. Recently, there has been much interest in PVL, due to its involvement in severe disease among children and young adults with no known exposure to healthcare establishments. In the United States of America, outbreaks of severe skin infections have occurred in homosexual men, prison inmates, and schoolchildren [23]. More worrisome, however, is the increasing number of reports of PVL-positive strains associated with severe necrotizing community-acquired pneumonia. An association between S. aureus strains carrying the genes for PVL and community-acquired pneumonia was first noted by Lina et al. [24]. They developed a PCR assay to detect the PVL genes and found a significant association between the presence of the locus and severe necrotic community-acquired pneumonia (8% of cases compared to no cases associated with hospital-acquired pneumonia). They also showed that the PVL genes were highly associated with primary cutaneous infections, especially furunculosis, confirming earlier findings by other researchers who used double immunodiffusion to detect the toxin [25]. More recently, cases of community-acquired pneumonia due to PVL-positive S. aureus have been reported in France [26], Sweden [27], the Netherlands [28], and the United Kingdom [29]. In addition, the PVL genes have been identified as a stable marker of community-acquired methicillin-resistant S. aureus (CA-MRSA) strains worldwide [30].
Superantigenic toxins
General aspects
Staphylococcal toxins denominated as superantigens began to be described in 1959 by Bergdoll et al. [31]. There are presently 18 serologically distinct enterotoxins designated by the letters SEA, SEB, SEC, SED, SEE, SEG, SEH, SEI, SEJ, SEK, SEL, SEM, SEN, SEO, SEP, SEQ, SER, SEU [31–35] and toxic shock syndrome toxin-1 denominated TSST-1 [36].
Staphylococcal enterotoxins (SEs) are monomeric, globular water-soluble proteins, with molecular weights from 26,000 to 29,000 daltons, that are rich in lysine, aspartic acid and glutamic acid and present cysteines forming the disulfide bridge [36]. They are relatively resistant to heat and to the proteolytic enzymes trypsin, pepsin and renin, which enables their passage through the gastrointestinal tract without losing activity [37–38].
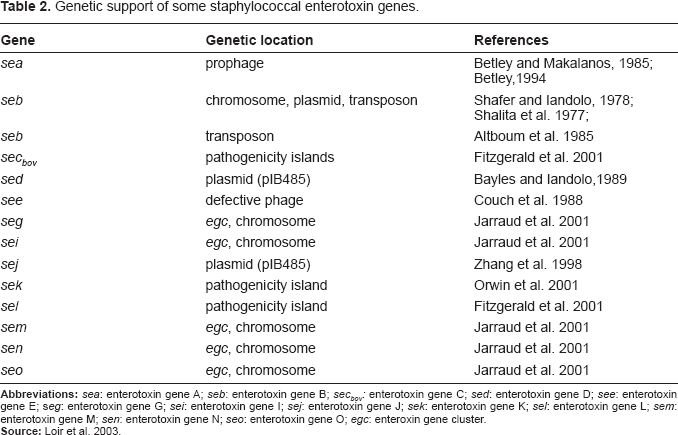
Although staphylococcal enterotoxins are similar in structure and biological properties, they differ in relation to genetic location, production and gene regulation mechanism [39]. Many of the genes responsible for virulence factors are located in strain-specific genetic elements. Genetic elements such as plasmids, transposons, bacteriophages and, more recently, pathogenicity islands have been described for S. aureus [40].
Genetic aspects
Homology percentage between amino acids from different classes of staphylococcal enterotoxins.
SECs are a group of highly preserved proteins of chromosomal origin that present three distinct subtypes: SEC1, SEC2 and SEC3, based on differences among antigenic determinants [39]. Other SEC molecular variations have been described according to the animal host to which they are related, SECbovine, SECcanine, SECovine [44].
Genetic support of some staphylococcal enterotoxin genes.
The amount of enterotoxin expressed by each enterotoxin class also differs. Bergdoll [50] describes that SEB and SEC are expressed in greater amounts than are other enterotoxins, frequently in the amount of 100 μg/ml of culture supernatant whereas the maximum production obtained for SEA, SED and SEE is usually less than 10 μg/ml.
Various regulation systems are involved in the determination of S. aureus virulence, such as the accessory gene regulator (agr), staphylococcal accessory regulator (sar) and various homologous sars, as for instance the repressor of toxins (rot) [51–53]. Of all these existing regulation systems, those which have been most extensively characterized are agr and sar [54].
On the agr locus, two promoters with opposite directions, P2 and P3, produce two transcripts, RNAII and RNAIII, respectively. RNAIII is formed by 510 nucleotides responsible for the transcription of genes for a large number of virulence factors, such as extracellular toxin, enzymes and cellular-surface proteins in Staphylococcus aureus [38]. RNAIII synthesis is induced when the concentration of an autoinducing polypeptide (AIP), specific in the medium, reaches certain levels which are generally detected in the exponential phase of growth for the stationary phase.
RNAII, in turn, is formed by four agr (accessory gene regulator) genes: agrB, agrD, agrC and agrA, arranged in an operon (agr), and they work together in order to induce RNAIII synthesis. In addition to its regulatory role, RNAIII is also an mRNA that encodes delta hemolysin in staphylococci. This hemolysin forms pores in membranes and lyses erythrocytes [55]. In S. aureus, this hemolysin is a polypeptide formed by 44 amino acids while, in S. epidermidis, it consists of 26 amino acids.
An agr operon has also been shown in some coagulase-negative staphylococcus species. In a study [56], RNAIII was detected in S. lugdunensis species; however, the detected mRNA did not encode the delta toxin, and its role in gene regulation is unknown. Other authors [57] described an agr locus in S. epidermidis with a homology of 68% as compared with the agr locus of S. aureus. S. epidermidis RNAII, similarly to S. aureus, also encodes proteins agr A, B, C, and D involved in the regulatory system of genes responsible for toxin production. The same authors also investigated delta toxin, produced during the post-exponential phase, in 23 samples of S. epidermidis and showed its presence in 21 of them [57].
Influence of physical and chemical factors
In addition to the various systems responsible for regulation of Staphylococcus spp. virulence factors, some medium-related factors, such as inoculum size, oxygen concentration, temperature, pH, NaCl concentration, mineral ions and medium composition act as external stimuli in the system's regulation, interfering in the production of protein A (surface protein) as well as in that of many other secreted proteins, acting independently of or interacting with agr [58].
With regard to pH, it is observed that the production of enterotoxins does not occur in pHs higher than 9.0 or lower than 5.0, and, at pH 8.0, a 50% decrease in their production takes place. Optimal pH for SEB and SEC production is 6.8 while SEA production can occur from 5.3 to 6.8 [59].
Thermostable enterotoxins and their inactivation depend on temperature as well as on the medium's purity, composition and pH. Studies have shown that SEC is the most thermostable of enterotoxins, followed by SEB and SEA [60].
Some studies have analyzed the production of SEA and SEB by CNS at different temperatures, pH values and NaCl concentrations, and most production occurred at a temperature of 39.4 °C with pH 7.0. When the temperature was increased to 41 °C at pH 4.5 and NaCl concentration of 12%, inhibition of enterotoxin production was observed [61].
Staphylococcal intoxication
Staphylococcal intoxication occurs after the ingestion of food contaminated with enterotoxin produced by bacteria of the Staphylococcus genus, whose main agent is S. aureus. However, some authors have reported the isolation of other enterotoxigenic coagulase-positive species, such as S. intermedius and S. hyicus [62–63].
Studies have also reported the enterotoxigenicity of coagulase-negative staphylococcus, including S. cohnii, S. epidermidis, S. saprophyticus, S. sciuri, S. warneri, S. chromogenes and S. lentus [40,64–68].
The major symptoms, which occur from 1 to 6 hours after toxin ingestion, are nausea, vomiting, diarrhea, abdominal pain, headache and muscle cramps. Clinical conditions are relatively mild, with durations from a few hours to one day. The symptoms may be severe at times, thus requiring hospitalization. This occurs as a result of the amount ingested and the individual's susceptibility, and may present dehydration, cephalea, sudoresis and body temperature alteration. Deaths are rare; however, they have been observed among children and elderly individuals [3].
Various studies have reported food contamination by asymptomatic carriers of S. aureus, and according to Carmo [69], in most cases of staphylococcal intoxication, food is contaminated by its handlers. Studies describe human beings as the major reservoir of S. aureus and report that 20 to 50% of normal individuals present this organism in the nasal vestibule and oropharynx [38].
Various studies have described the presence of enterotoxigenic S. aureus isolated from food handlers, some of which were associated with food intoxication outbreaks where the presence of type-SEA and type-SEE enterotoxins was detected [69–71]. Among the staphylococcal enterotoxins, SEA is the one most commonly involved in food intoxication outbreaks in the U.S.A, corresponding to 77.7% [72], and to 81%–90% in Taiwan [73].
When investigating workers in restaurants in Kuwait, some authors obtained 14.1% of samples positive for toxigenic CNS from a total of 142 samples isolated from the hands and 13 samples isolated from the nasal cavity. The CNS samples belonged to the species S. epidermidis, S. hominis, S. warneri, S. saprophyticus, S. xylosus, S. haemolyticus and S schleiferi, which produced enterotoxins SEA, SEB, SEC and TSST-1, with SEB being prevalent [68].
The precise amount of enterotoxin necessary to cause food intoxication is not known. In a study on human volunteers to whom enterotoxin was administered, it was estimated that only 1–5 μg/70 Kg would be capable of triggering symptoms [5]. Evenson et al. [74] described an outbreak involving children who had ingested contaminated chocolate milk. The enterotoxin concentration found was only 0.5 ng of SEA/ml.
Superantigenicity
In addition to their important role in staphylococcal food intoxication, enterotoxins can also activate the nonspecific proliferation of T-cells; therefore, they are denominated superantigens.
Bacterial superantigens are capable of nonspecifically activating T-cells by externally bonding to the Vβ domain of the T-cell receptor and to the a chain of the class-II MHC molecule with previously unprocessed antigens. This bond produces a signal that induces the proliferation and polyclonal activation of approximately 10 to 30% of the T-cell repertoire, and CD4 T-cells comprise the prevailing responsive population. Hence, a large production of post-inflammatory cytokines from T-cells, such as IFN-γ and TNF-α as well as IL-2 from monocytes, including IL-1 and TNF-α, is observed. The massive production of post-inflammatory cytokines triggers an intense inflammatory response, causing damage to the host's tissues [75].
TSST-1 [36] is responsible for toxic shock syndrome (TSS), an acute systemic disease characterized by fever, arterial hypotension, cutaneous rash and skin scaling. But on some occasions, isolated lineages from TSS patients do not produce TSST-1. As indicated, enterotoxins A, B and C may be related to clinical manifestations of the disease [5]. TSS, which was previously associated with the use of vaginal tampons, may occur after any staphylococcal infection caused by a TSST-1-producing lineage, and S. aureus is the primary species involved [76]. Nevertheless, some reports indicate TSST-1 is also produced by CNS [68,77–80].
Tarkowski and Wagner [81] described septic arthritis as a cause for the action of enterotoxins produced by enterotoxigenic species of S. aureus which, when activating an exaggerated inflammatory response mediated the cytokine action, in turn activating a large number of T-cells and macrophages responsible for tissue damage.
Similar results are described by Bremell and Tarkowski [82] in an experimental model inoculating staphylococcal enterotoxins in rats, where they demonstrated that 100% of the rats receiving enterotoxins SEA, SEB, SEC and SED developed arthritis. Mortality was also preferentially induced in rats inoculated with SEB and SED from S. aureus.
When analyzing 107 Staphylococcus samples isolated from the feces of 5-year-old children with sporadic diarrhea, Efuntoye and Adetosoye [83] obtained 72 strains of S. aureus, 33 of S. epidermidis and 2 of S. saprophyticcus. Of the 72 S. aureus samples isolated, 61 (84.7%) produced enterotoxins. SEA was prevalent as it was produced by 37 (60.7%) of the enterotoxigenic strains of S. aureus; 12 (19.7%) produced only SEB; 2 strains produced SEA and SEC; 6 strains produced SEA, SEC and SED, and only one 1 strain produced SEA, SEB and SED.
In a recent study performed by Berstein et al. [84], a possible role of staphylococcal enterotoxins in sinusitis with nasal polyps is suggested. In that study, 55% of the investigated patients showed enterotoxigenic S. aureus in the nasal mucosa adjacent to the polyps, where three different enterotoxins were isolated: SEA, SEB and TSST-1.
Staphylococcal enterotoxins also play a role in other serious pathological processes related to sepses, osteomyelitis and respiratory distress syndrome, the latter of which is characterized by a pulmonary dysfunction almost always associated with a septic process [85].
Detection
Various methods have been developed in order to detect the presence of enterotoxins, and immunoassays have been used most frequently due to their simplicity and sensitivity. They are based on the use of enterotoxin-specific polyclonal and monoclonal antibodies [86].
RPHG (Reverse Passive Hemagglutination), which uses the adsorption of specific antibodies in red blood cells, was the first method employed to be regarded as truly sensitive as it is capable of detecting 1 ng/g of enterotoxin present in food. The disadvantages of method are nonspecific hemagglutination and the difficulty in adsorbing certain antibodies on the surface of red blood cells [87].
The system was enhanced by replacing erythrocytes with latex particles—Reverse Passive Latex Agglutination (RPLA), which increased sensitivity to 0.75 ng/g of enterotoxins. Nevertheless, nonspecific reactions still occur with certain food types [88] and difficulty persists in distinguishing such nonspecific reactions [89].
Radioimmunoassay (RIA) is based on the competition between the toxin marked with I125 and the unmarked toxin, which compete for the binding site of the antibodies adsorbed in the solid phase. This method proved an apt option due to its sensitivity level of 1.0 to 5.0 ng/ml [90]. However, the need for highly purified enterotoxins and the handling of radioactive material limited its use.
Due to the need to eliminate the risk of contamination by RIA, the ELISA (Enzyme-linked immunosorbent assay) technique was developed, in which the radioactive marker was replaced by the enzyme, and presently many of such tests show a sensitivity level of 0.1 to 1.0 ng/g of food [91].
In addition to RPLA, there are other commercially available kits. They use a mixture of specific polyclonal antibodies that detect all enterotoxins without differentiation, such as the SET-EIA kit, which detects SEA, SEB, SEC and SED at a sensitivity level of 0.1–1.0 ng/ml. Other kits, namely, TECRA (Bioenterprises Pty. Ltd., Roseville, New South Wales, Australia), TRANSIA (Transia-Diffchamb S. A. Lion, France) and VIDAS (BioMérieux Vitek, Inc., Hazelwood, Missouri), capable of detecting SEA-SEE, are also available [92].
The RIDASCREEN kit (R-Biopharm GmbH, Darmstadt, Germany) individually detects enterotoxins SEA, SEB, SEC, SED and SEE, since it is standardized with monoclonal antibodies and presents high sensitivity (0.2–0.4 ng/ml) and specificity, requiring three hours for analysis [93].
However, these methods do not always detect toxin-producing lineages since their production depends on growth conditions that do not represent in-vivo conditions [94]. Moreover, immunological methods may produce false-positive results due to cross-reactions between antigens and the occurrence of nonspecific reactions [88].
With the development of molecular biology, some techniques have been proposed for the detection of the gene responsible for toxin production. One of the first techniques to be used involved probes that correspond to sequences of nucleotides marked by radioactive substances, biotin, sulphonyl groups or alkaline phosphatase. The hybridized probes are read by means of auto-radiography or colorimetry [95].
At present, Polymerase Chain Reaction (PCR) is one of the most frequently used methods as it enables the identification of the genes responsible for enterotoxin production with high sensitivity and specificity. Protocols for gene detection have been developed by using the sequence of oligonucleotides obtained by the computerized analysis of gene sequences sea, seb, sec, sed and see of the respective staphylococcal enterotoxins SEA, SEB, SEC, SED and SEE [96]. A later study conducted by Jarraud [97] included the sequences referring to the genes of enterotoxins G and I.
In a study conducted by Padmaprya [98], the identification of enterotoxic strains of Staphylococcus species by identifying a staphylococcal accessory regulator (sar) was suggested. By using sar-specific primers, that author detected 49 sar-positives from a total of 67 samples. The results were compared to a PCR performed on the genes of enterotoxins A, B, C and D, which showed the presence of the enterotoxin genes in all sar-positive samples.
When investigating samples of CNS and S. aureus isolated from different foods in French restaurants, Rosec and Gigaud [99] observed that none of the 74 isolated CNS samples contained toxigenic genes whereas, of the 258 S. aureus samples tested, 76 showed toxin genes. In Brazil, when investigating the presence of toxigenic genes in CNS samples isolated from food by the PCR technique, Cunha et al. [80] found 10% of samples positive, of which 25% were positive for gene sec-1 and 75% for gene sea. However, other authors did not achieve positive results in CNS samples. When investigating the presence of toxin genes by the multiplex PCR technique in CNS samples and other non- S. aureus coagulase-positive species isolated from clinical materials, Becker et al. [100] did not detect toxin genes in the tested samples.
Cunha et al. [101] used the PCR technique to detect the genes responsible for enterotoxins in staphylococcus lineages isolated from newborns. The PCR results obtained were compared to those by the RPLA method. From the total number of 120 S. aureus samples isolated, 38.3% were enterotoxin producers, according to RPLA, whereas PCR found 46.6% of these to be positive. Coagulase-negative staphylococcus presented 40.0% positive lineages by PCR as compared to 26.7% by the RPLA method.
Cunha et al. [101] observed the production of TSST-1 and SEB + SEC in S. epidermidis and encountered SEA + SEB and SEA + SEC associations in S. aureus, which were not confirmed as positive when these genes were investigated by the PCR method. Tsen and Chen [102] also found positive S. aureus lineages by RPLA and negative lineages by PCR. This is one of the disadvantages presented by the immunological methods in which false-positive reactions may occur.
These advantages are not found in the direct detection of the genes responsible for toxin production. According to Schmitz et al. [103] the staphylococcus strains that are positive for gene detection could be regarded as strains with potential for toxin production. The PCR technique also enables the genotypic identification of various staphylococcal species with high sensitivity and specificity. By using the Internal Transcribed Spacer-PCR (ITS-PCR), Couto et al. [104] identified different CNS species, thus amplifying an extremely conserved region in the genus. Prokaryotes have genes that are responsible for the expression of rRNA 16S, 23S and 5S. These genes are separated by regions showing a high level of variability in their sequences according to genus and species. This amplified region, when viewed in polyacrilamide gel, shows a specific band pattern for each CNS species. By using their respective ATCCs, it is possible to compare the band pattern of the reference samples with those of the investigated samples so that no doubts remain as to the accurate identification of different staphylococcus species.
Although reports of toxigenic genes in CNS species can be found in the literature, many authors still question their toxigenic potential; therefore, genotypic identification is important for the confirmation of CNS species. Lotter and Genigeorgis [105] reported not believing that coagulase-negative staphylococci could produce enterotoxins and suggested the possibility of error in species identification. Later, Fox et al. [106] reported the isolation of S. aureus lineages that produced little coagulase or did not express that enzyme, and identified them as coagulase-negative staphylococci. Hence, genotypic identification confirms phenotypic identification by excluding the possibility of error.
With the purpose of rejecting this possibility, Cunha et al. [107] performed genotypic identification by the ITS-PCR technique for species confirmation in CNS samples presenting toxigenic genes. Those authors observed that all the samples identified as CNS by the phenotypic method were confirmed to be CNS by the genotypic technique.
The PCR technique, however, enables the detection of genes contained in the lineages independently of their expression, since, although the gene is present in the microorganism, it cannot be active. The development of genotypic methods also provides the possibility of detecting the messenger mRNA (mRNA) sequence responsible for the target-enterotoxin expression, by using the reverse transcription polymerase chain reaction (RT-PCR). The RNA sample is first reverse transcribed to cDNA with reverse transcriptase. Gene expression is usually directly proportional to the number of copies of mRNA of a particular gene. In order to detect gene expression from small amounts of RNA, amplification of the gene transcript is necessary by the PCR technique, utilizing specific primers to confirm the expression of genes by the cDNA of the mRNA. The expression of the mRNA sequence which encodes toxin synthesis leaves no doubts as to the microorganism's toxic potential.
Toxigenicity in Coagulase-Negative Staphylococci
The existing disagreement concerning the enterotoxigenicity of CNS and their capacity to cause food intoxication and/or other associated diseases points out the need for further studies using reliable techniques that can confirm the capacity of such staphylococci to produce toxins. In a study on CNS samples isolated from newborns in Brazil which used the RT-PCR technique, Cunha et al. [107] observed that out of 14 CNS samples presenting toxigenic genes for SEA, three confirmed the mRNA expression of this toxin whereas of the 33 samples positive for SEC by PCR, only three confirmed its expression. S. epidermidis was the most toxigenic species among CNS, and five samples were positive for mRNA expression that encodes SEA and SEC. Among other CNS species, only S. lugdunensis displayed positive results for SEC by the RT-PCR technique. None of the thirteen samples presenting genes for TSST-1 showed toxin production when investigated by the RT-PCR technique.
Some authors have also reported the production of enterotoxins and TSST-1 by S. epidermidis, S. haemolyticus and S. warneri by using the RPLA method [79]. When investigating food handlers, Udo et al. [68] also found, by using the RPLA method, samples of CNS and S. aureus that produced enterotoxins and TSST-1. Of the total CNS investigated, 14.1% were enterotoxins or TSST-1 producers, and samples of the species S. hominis, S. warneri, S. saprophyticus, S. epidermidis, S. xylosus, S. haemolyticus and S. schleiferi were positive for toxins SEA, SEB, SEC and/or TSST-1. Marin et al. [66] describe a study performed on enterotoxin-producing staphylococci in ham samples by the RPLA method. Of the 135 staphylococci isolated, two belonged to the S. epidermidis species and one was an SEC producer.
When studying the virulence factors of CNS isolated from newborns, Cunha et al. [108] found, by the RPLA method, that 37.6% of CNS had produced SEA, SEB or SEC. The species S. epidermidis, and S. lugdunensis produced SEA, SEB and SEC whereas S. haemolyticus, S. hominis and S. simulans produced only SEC.
Although various authors have questioned the toxigenic potential of CNS [105–106], operon agr, which plays an important role in regulating the expression of staphylococcal toxins, has also been found in other staphylococcus species, such as S. intermedius [109], S. lugdunensis [56] and S. epidermidis [57]. In addition to these regulatory systems present in the microorganism, some factors from the external environment may act directly on enterotoxin production. It is known that enterotoxin production is optimal in neutral pH and decreases in acid pH. Bergdoll [50] found that glucose in the culture medium has an important inhibitory effect on enterotoxin production, especially on enterotoxins SEB and SEC. Such inhibition is due to a pH decrease as a result of glucose metabolism. According to Regassa et al. [110] and Novick and Jiang [111], glucose and pH decrease may be associated with agr expression. Pereira et al. [61] analyzed SEA and SEB production by CNS at different temperatures, pH values and NaCl concentrations. Most production occurred at a temperature of 39.4 °C with pH 7.0. Therefore, enterotoxin synthesis may differ depending on laboratory and natural conditions.
The exact mechanism by which these systems are activated or inhibited has not yet been fully clarified. The presence of an agr locus in S. epidermidis species and its capacity to produce delta toxin associated with this agr system [57] demonstrate that the system is acting and does not exclude the possibility of enterotoxin production at any moment. Therefore, the prevalence of S. epidermidis among the toxigenic CNS species may be associated with staphylococcal regulatory systems that are also present in this species.
The RT-PCR method is a rapid and efficient technique, and it clearly shows that CNS are capable of expressing mRNA which encodes staphylococcal enterotoxins; however, as it depends on gene activation, further investigation on environmental factors and clarification of the regulation mechanisms that may interfere with its expression must be evaluated.
Various studies distinguish CNS as important pathogens and point out their toxigenic potential by emphasizing that greater attention must be given to such microorganisms, which are still often considered to be merely contaminants.