
Abstract
There is disagreement among microbiologists about whether Legionella requires a protozoan host in order to replicate. This research sought to determine where in biofilm Legionellae are found and whether all biofilm associated Legionella would be located within protozoan hosts. While it is accepted that Legionella colonizes biofilm, its life cycle and nutritional fastidiousness suggest that Legionella employs multiple survival strategies to persist within microbial systems. Fluorescent in situ hybridization (FISH) and confocal laser scanning microscopy (CLSM) demonstrated an undulating biofilm surface architecture and a roughly homogenous distribution of heterotrophic bacteria with clusters of protozoa. Legionella displayed 3 distinct spatial arrangements either contained within or directly associated with protozoa, or dispersed in loosely associated clusters or in tightly packed aggregations of cells forming dense colonial clusters. The formation of discreet clusters of tightly packed Legionella suggests that colony formation is influenced by specific environmental conditions allowing for limited extracellular replication. This work represents the first time that an environmentally representative, multispecies biofilm containing Legionella has been fluorescently tagged and Legionella colony morphology noted within a complex microbial system.
Introduction
The ecological niche of Legionella is often disputed. An ongoing debate exists over the multiplication requirements of Legionella within environmental biofilms. Some reports suggest growth of Legionella occurs solely via parasitization of amebic hosts, and others that multiple survival strategies exist that exploit other avenues of nutrition and proliferation. 1
As our understanding of microbial ecology expands, research becomes increasingly directed towards whole system biology rather than individual species interactions. Biofilms represent a pinnacle of microbial ecology, being complex aggregations of bacteria, fungi, viruses and protozoa, and higher eukaryotes that physiologically coordinate to form colonies that efficiently sequester nutrients.2–4 Biofilms persist under conditions unfavorable to microbial growth, proliferate, and disseminate their component organisms.3,5–8 Legionella bacteria are the cause of Legionnaires’ disease, a serious bacterial pneumonia. They are of particular interest due to their survival in and dissemination from environmental biofilms.9–12
Biofilms harboring human pathogens, such as Legionella, present problems, as their removal from industrial water systems and cooling towers is problematic and the emphasis has been placed upon control rather than removal as the more realistic option.13–17
It is inadequate to study mixtures of planktonic bacteria as a measure of microbial community function and ecology. In order to elucidate the range of processes that occur in biofilms, various systems have been designed that provide adequate conditions for biofilm formation.18–21 Bioreactors and chemostats are common in industrial microbiology, providing optimal growth conditions for biofilm formation. Conventional bioreactors do not resemble industrial water systems, heat exchangers, or cooling towers, and, thus, the resulting films may not be representative of environmental systems. This paper outlines the use of a modified Robbins device, which has been constructed to resemble an industrial heat exchanger/cooling tower in its operation to produce lab-scale analogues of environmental biofilms containing Legionella. 22
Optical methods to observe biofilms have been used to give insight into the architecture of communities and their formation and persistence.22–24 Visual reconstruction of the components of these films provides a nonmolecular avenue of study into the phenomena occurring within complex microbial systems. Developing tools to study interactions between biofilm associated organisms may eventually lead to an understanding of critical events or stages in biofilm formation and better target our attempts to remove them from water systems.
Methods
Biofilm growth in a modified robbins device/chemostat
To culture Legionella and biofilm associated microflora, modified chemostats were constructed that resembled heat exchangers in their function and design (Fig. 1). Each system consisted of a reservoir of 10 L volume containing a sparger to provide adequate aeration and a heater to maintain a temperature of 35 °C. Sparged air was vented into a moisture trap to ensure no aerosols were generated, and collected condensate was returned to reservoirs. Any fluid deficiency was made up using distilled water in order to maintain the homeostasis of the system. Water was circulated at a speed of 1 L per minute through a system at of polypropylene pipes, with an approximate volume of 3 liters, via an adjustable peristaltic pump (Masterflex L/S variable-speed modular drive, Cole-Parmer®, Illinois, USA) before being returned to the reservoir. The flow rate of 1 L per minute was chosen in order to achieve a laminar flow within the pipe systems (Reynold's number of <2000). Twelve sampling points were spaced at equal intervals along the length of the piping at which coupons of various materials may be added and periodically sampled for biofilm deposition and analysis. During the course of the study, 6 stainless steel and 6 polypropylene strips taken from cooling tower fill were used as biofilm growth substrates. Coupons were cut to provide a final size of 10 cm long and 1 cm wide. Steel coupons were 1 mm thick and polypropylene 0.25 mm thick.

Modified Robbins device consisting of an aerated, heated reservoir connected to a series of opaque pipes.
Each system was filled with water sourced from the cooling tower at the Flinders Medical Centre, South Australia, and was provided with 0.05% (final concentration in system) yeast extract to stimulate initial colonization of the system by native microflora. To further ensure that a representative microbial community was present in biofilm, systems were seeded with water derived from a cooling tower associated with a Legionnaires’ disease outbreak. This process involved the addition of outbreak associated cooling water to 20% of the final volume of each system (3 L added, final volume ~15 L). Systems were continuously operated without further addition of nutrients or microbial inoculum for a period of 1 month before biofilm was sampled.
Biofilm sampling
Coupons were removed from the system and placed into tubes containing 50 mL Milli-Q water (A10 Academic water purifying system, Millipore, MA, USA). For microscopic visualization, tubes were gently inverted to remove attached biofilm without disrupting film structure. This method was chosen as coupons fit perfectly within 50 mL disposable tubes, with little room for movement by the coupon during inversion, reducing damage to biofilms. As the edges of each biofilm became detached, larger areas of film sloughed from the substrate allowing for the removal of large, mostly intact sections. This method allowed for the harvesting of high quality pieces of biofilm, with minimal damage to microbial structures and overall film architecture.
For culture, tubes were vortexed for 60 seconds, followed by sonication for 60 seconds to break apart biofilm structure.
Microbiological characterization of water systems
The bacterial load of system water was determined using methods described by Chen et al. 25 One cm2 sections of coupon surfaces were scraped with cotton swabs to detach biofilm, placed in sterile water, and vortexed for 60 seconds, followed by sonication for 60 seconds to break apart biofilm. Samples were serially diluted to 1 × 10- 7 and 10 µL aliquots of 1 × 10- 2 to 1 × 10- 7 dilutions were carefully transferred onto R2a agar plates to form a 6 × 6 grid, with adequate spacing to ensure no drops touched. Plates were left to dry before incubation at 22 °C overnight. The number of colonies in each dilution series were counted and used to calculate the number of colony forming units (CFU) per cm2 of coupon surface.
Coupon polysaccharide measurement and comparison
To determine the polysaccharide concentration on coupon surfaces (which can be used as an estimate of the total biofilm present), 1 cm2 sections of coupon surfaces were scraped with cotton swabs to detach biofilm and vortexed in 1 mL sterile water to dislodge biomass from the swabs. Coupons were swabbed at the base, center, and tip to determine if sample location had any influence on the polysaccharide concentration. Polysaccharide content of each sample was analyzed using adaptation of methods by Masuko et al. 26
Isolation and culture of Legionella from biofilm
The presence of Legionella within system derived biofilm was confirmed by isolation and culture using a modification of methods by Bopp et al. 27 Coupons were placed in 50 mL of sterile water and vortexed for 60 seconds, followed by sonication for 60 seconds to break apart biofilm structure. Settled biofilm was collected and serially diluted into HCl:KCl buffer, adjusted to pH 2.2, and incubated at room temperature for 12 minutes. Then, 100 µL aliquots were plated on selective media consisting of buffered charcoal yeast extract agar (BCYE) supplemented with L-cysteine, α-ketoglutarate and ferric-pyrophosphate, and a selective supplement of glycine, vancomycin, polymixin, and cycloheximide (GVPC) (Oxoid, Adelaide, Australia).
Plates were incubated in candle jars at 37 °C and observed for Legionella-like colonies at 3, 5, and 7 days of incubation using a dissecting microscope. Colonies displaying representative Legionella-like morphology–-circular, off white, with a grainy ground glass appearance and opalescence–-were subcultured onto clean BCYE agar and blood agar. Colonies displaying growth on BCYE and not on blood agar were tested using latex agglutination (Legionella latex test kit, Oxoid, Adelaide, Australia) for positive identification as Legionella pneumophila.
Fluorescent in situ hybridization and mounting
Oligonucleotide probes were selected using Probe-base 28 to select sequences specific to organisms of interest. Three probes were selected to target Legionella LEG705 (5‘-CTG GTG TTC CTT CCG ATC-3’), 29 prokaryotes EUB338 (5‘-GCT GCC TCC CGT AGG AGT-3’), 30 and eukaryotes UNIV1389b (5‘-ACG GGC GGT GTG TAC AAA-3’). 31 Probes were tagged with distinct fluorophores with little to no overlap in fluorescence emission, LEG705 with Alexa Fluor 647, EUB338 with Alexa Fluor 546, and UNIV1389b with Alexa Fluor 488 (Invitrogen, Melbourne, Australia).
Sampled biofilm was transferred to tubes containing 4% w/v paraformaldehyde and incubated at 4 °C overnight. Samples were transferred to microscope slides and air dried at 35 °C. Fixed samples were washed once with phosphate buffered saline (PBS) and dehydrated in a series of ethanol baths, of 50%, 80%, and 96% respectively.
Samples were hybridized using an adaptation of methods by Grimm et al. 32 Samples were covered with 200 µL hybridization buffer (0.9 M NaCl, 0.01% sodium dodecyl sulphate, 20 mM tris-HCl [pH 7.6]) containing 100 ng of each probe, covered with hybridization chambers to maintain optimal humidity, and incubated for 90 minutes at 46 °C.
Unbound probes were removed by incubating samples with warmed washing buffer (0.9 M NaCl, 0.01% sodium dodecyl sulfate, 20 mM Tris-HCl [pH 7.6]) for 10 minutes at 48 °C. Samples were given a final wash with Milli-Q water and air dried in the dark before mounting with Citifluor (Citifluor ltd, Leicester, UK).
Negative and positive controls were generated to ensure probes did not bind to nontarget organisms. Each probe was tested against a culture of Legionella, E. coli (ATCC 25922) and biofilm derived Bacillus cereus, and visualized to ensure the absence or presence of fluorescence.
Confocal laser scanning microscopy
Images were acquired using a Leica SP5 confocal microscope. Bandpath filters were set to encompass the maximal emission of each fluorophore to remove interference from autofluorescence from film components. High resolution image stacks were compiled for regions of interest, encompassing scans through the full thickness of each film sample.
All imaging work was carried out at Flinders Microscopy using Leica LAS AF software and imageJ. 33
Results
Microbiological content of biofilms
The presence of Legionella in biofilm was confirmed by culture and latex agglutination (Oxoid, Adelaide, Australia).
Plate count biofilm organisms
Plate count results generally showed wide variation between replicates. Plate counts showed an average CFU/cm2 of 9.6 × 10 5 with a standard deviation of 1.5 × 10 6 . Plates tended to become overgrown rapidly at lower dilutions, making counting impossible, while higher dilutions showed large variability in the number of countable colonies. This is a result of the strongly self-adhesive nature of biofilm, which does not neatly homogenize by agitation or pipetting. This leads to difficulties in sampling as any dilution or attempts to break up samples generally results in an assortment of fragments, which may deposit unevenly leading to high variability in results.
Biofilm deposition on coupons
Biofilm deposition was relatively homogeneous between coupons (Fig. 2) but varied along the length of the coupon, with greater deposition on the proximal end. Sample location showed minimal influence over carbohydrate content of sample except where large sections of biofilm remain attached to coupons, resulting in large spikes in variation. Comparisons of coupon material and related biofilm showed no significant differences (P > 0.05) in quantified polysaccharide, with an average polysaccharide concentration of 317 ± 35 mg/cm2 on stainless steel coupons and 299 ± 48 mg/cm2 on polypropylene coupons.

Black polypropylene substrate with associated biofilm. Biofilm appears as rusty orange/brown depositions on the surface of the plastic.
Biofilm tended to be a rusty brown/orange color on coupons, which were not exposed to light. Biofilm forming in the semitransparent tubing and reservoir tended to be a dark green color due to the presence of algae. Associated with biofilms were a wide range of grazing organisms, including ciliates and amebae, rotifers, and nematodes. There appeared to be no discernible difference in the presence of Legionella or the structure and composition of biofilms derived from either polypropylene or steel coupons.
Probes showed little to no nonspecific binding in controls and fluorescently tagged biofilms. Organisms of interest appeared distinctly against background fluorescence and with identifiable microbial structures appearing within the appropriate fluorescent channels.
Figure 3 is a typical representation of biofilm harvested from this system, with inset image of a transverse section of the film showing undulating surface architecture. Fluorescence has been artificially colored to increase contrast. EUB338 (all bacteria) was colored red, LEG705 (Legionella), blue and UNIV1389b (eukaryotes), green. Some localized areas of increased bacterial density were present, which punctuated an otherwise homogeneous distribution. Protozoa generally appeared clustered, that is, appearing as groups of green spheroids visible in the top right and middle left of Figure 3. Samples were previsualized using light microscopy to observe the presence of chlorophyll and to further discriminate between protozoa and algae. Legionella appeared as distinct groups of cells (indicated by arrows), often surrounding or associated with protozoa, and occasionally as high density clusters or dispersed groups of cells in localized areas of biofilm. Diffuse, low intensity fluorescence was occasionally seen where the eukaryote probe bound to targets originating from eukaryotic cell remnants or nonspecifically to EPS.

Typical representations of FISH tagged biofilms from this system. (A) is annotated to highlight Legionella colonies (indicated by arrows), with inset image showing transverse view displaying undulating surface. Fluorescent tags have been artificially colored to increase image contrast. Eukaryotes appear green, Legionella, blue, and all other bacteria, red.
Figure 4 presents a series of common representations of Legionella colonies in fluorescently tagged biofilms derived from this system. Legionella appeared in 3 conformations within biofilms derived from this system. Figure 3A shows a cluster of protozoa (green) that appear as elliptical/spheroid forms surrounded with Legionella (blue). Where Legionella was observed interacting with protozoa, typically trophozoites (not shown) and cysts were surrounded or pervaded with substantially higher numbers of Legionella bacteria than in other areas of biofilm. Figure 4B shows a high concentration of clustered Legionella. Cells in this conformation were very closely packed with little to no edgewise dispersion indicating a lack of motility. Figure 4C shows Legionella in a loosely associated group. Cells are separated by distances ranging from 1 to 10 microns, clustered in a roughly spherical distribution. This suggests cells are motile and are displaced over the remains of a lysed protozoa indicated by weak fluorescence.
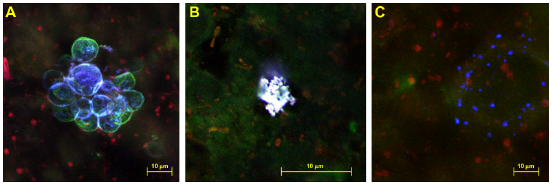
Legionella appeared in 3 distinct colony morphologies, either present associated with or encapsulated within protozoa (A), as closely grouped collections of cells (B), or as individual cells apparently dispersing from a roughly central origin (C).
Discussion
This study represents the first time that the structure of a complex, multispecies biofilm containing Legionella has been visualized and reconstructed using FISH and CLSM. Previously, Legionella has been visualized parasitizing amebae using fluorescent in situ hybridization (FISH), confocal laser scanning microscopy (CLSM), and epifluorescence microscopy. 32 To date, studies have not been conducted to visualize Legionella in intact environmental biofilms. Although studies of Legionella/protozoa interactions in cocultures are valuable for understanding Legionella growth, extrapolation of the observed interactions into the context of complex multispecies environmental biofilm processes are inadequate, as they avoid the complexity of these systems. Processes such as nutrient cycling,34,35 quorum sensing, predation, and cometabolism 36 may significantly modify the growth and interactions that these organisms have with one another.
The modified chemostat developed for use in this study provides a suite of abiotic conditions which may be altered to suit the requirements of the experiment undertaken. Temperature, aeration, and flow rate may be adjusted to mirror the conditions common to many industrial water systems and heat exchangers. The length of the polypropylene piping housing containing the test surfaces allows for a natural temperature gradient to form, reflecting the conditions within a cooling tower connected to a heat exchanger. Test surfaces are interchangeable, which allows a wide range of materials to be studied for their biofilm forming potential. Small sections of transparent tubing and a transparent reservoir allowed for algal growth in this study; however, all piping can be replaced with opaque or transparent alternatives to provide the required light levels if other photobiological effects are to be studied.
This approach is limited, however, by the inability to fully characterize the microbiological content of the water used to inoculate the system and any biofilm derived from this inoculum. As a concession, light microscopy can be used to determine the presence of larger grazing organisms, and, as presented in this study, a range of relatively simple microbiological techniques can be employed to give an estimate of the bacterial activity/load of the system.
Figures 3 and 4 demonstrate that Legionella colonies tend to exist as distinct clusters of cells rather than being homogenously distributed throughout the film, often forming larger colonies around protozoa or protozoan cysts. This dispersion pattern is common is biofilm inhabiting organisms, 24 but Legionella presents a unique case, as its ecological niche, multiplication, and survival mechanisms and still in debate.1,37–39 If Legionella survives as an obligate intracellular pathogen with little extracellular survival capability outside of protozoan hosts, then the vast majority of Legionella would be visible associated with these hosts. If other survival mechanisms are employed, then groups of cells should exist in a similar fashion to other common biofilm inhabiting organisms, as noted in Figure 4B.
Numerous studies have demonstrated Legionella's capability to multiply intracellularly within suitable amebic and macrophagic hosts.40–47 Figure 4 shows the localization of Legionella to specific regions of the film, which may reflect its lifecycle and ability to act as an intracellular pathogen of amebae.
Legionella multiply within hosts until amino acid levels become too low to facilitate further growth, at which point they become flagellated assisting in host lysis and egress. 48 This process would leave pockets of Legionella located around the lysed remains of their host in relatively uniform groupings of dispersed cells originating from the remains of their host (Fig. 4C) or located within amebic hosts during intracellular replication (Fig. 4A). This presents a distinctly different appearance than Legionella in Figure 4B, which are clearly not associated with a protozoan host and are nonmotile as denoted by their tight clustering.
Legionella has been demonstrated to multiply on algal exudates49,50 and saprotrophically on the remains of other dead organisms if the nutrient content is sufficiently high. 51 The closely grouped cells notable in Figure 4B may reflect the formation of nutrient rich areas within the biofilm that provide the conditions required to form the observed colonies.
Recent biochemical work has shown that Legionella produces at least one siderophore, legiobactin, 52 a compound that chelates Fe3+ increasing its solubility and bioavailability. Many aquatic bacteria possess siderophores, and their production is a necessity for multiplication in low iron environments such as soil, aquatic and marine sediments, and their associated biofilms. 53 The role of iron in Legionella multiplication has been well documented, and it is known to be a crucial nutrient required for the growth of Legionella. 54 Cianciotto 52 demonstrated that Legionella possess siderophore synthetase and secretion genes, lbtAB, with structures homologous to E. coli, Sinorhizobium, and Bordetella. lbtAB deficient Legionella mutants were still capable of normal infection and multiplication inside human macrophages, hartmanella, and mouse lungs. 52 This strongly suggests that the presence and expression of this siderophore is required for growth and multiplication within aquatic environments rather than during intracellular growth.
The ecological niche of Legionella may not be as straightforward as previously assumed. Parasitism appears to be the primary mechanism by which Legionellae multiply in environmental sources but may represent only one survival strategy in a relatively complex lifecycle. Work by Garduno et al55,56 has demonstrated the existence of multiple morphological and life cycle stages incorporating periods of dormancy in preparation for intracellular growth. Although this work is not designed to quantitatively compare the likelihood of specific spatial arrangements of Legionella occurring in biofilms, we suggest that Legionella persists in a dormant state within a biofilm following periods of intracellular replication; however, periods of limited extracellular growth may occur when localized biofilm conditions are nutritionally favorable to multiplication. The ability to culture Legionella on nutritionally supplemented media without the presence of a host attests to this replication mechanism. It is also possible that Legionella is able to multiply on the nutrient rich remains of lysed protozoa displaying some chemotaxic or, more accurately, a chemosensing ability. The nonmotile L. longbeachae genome displays the presence of a chemotaxic region. 57 This suggests that some Legionella species may be able to alter gene expression and cellular activity after existing in a viable, nonproliferative stage in response to the presence of protozoa or nutritionally favorable conditions.
This work provides new information for the ongoing discussion of the ecological niche of Legionella within biofilms. Presently, no clear consensus exists as to whether Legionella is capable of nonparasitic multiplication in environmental water sources. This work demonstrates the presence of microcolonies of Legionella not associated with protozoa. If samples from multiple disparate sources could be analyzed using similar microscopic methods, a model for Legionella colonization of biofilms, independent of the presence of protozoa, could be generated based upon the conditions present in each film, such as temperature, nutrient content, and pH.
As our understanding of microbial ecology, biofilms, and quorum sensing increases, so does our capacity to control microbial colonization of surfaces and reduce their persistence. Visualizing the interactions between microbes in an environmentally relevant setting, when coupled with molecular and culture methods of study, provides a greater depth of understanding of the processes occurring within these complex microbial systems. By altering the conditions that mediate colony formation and overall biofilm architecture, we may gain insight into their control and removal from constructed water systems.
Author Contributions
Conceived and designed the experiments: MT. Analyzed the data: MT. Wrote the first draft of the manuscript: MT. Contributed to the writing of the manuscript: MT. Agree with manuscript results and conclusions: MT, RB, KR. Jointly developed the structure and arguments for the paper: MT, RB, KR. Made critical revisions and approved final version: MT, RB, KR. All authors reviewed and approved of the final manuscript.
Funding
This work was supported by a grant from the Victorian Department of Human Services.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests.
Footnotes
Acknowledgments
The authors acknowledge Flinders Microscopy and Dr. Jennifer Clarke for expert technical advice and for assistance and with fluorescence and confocal microscopes, and the Victorian Department of Human Services for their support of this work.