The genus Listeria consists of a closely related group of Gram-positive bacteria that commonly occur in the environment and demonstrate varied pathogenic potential. Of the 10 species identified to date, L. monocytogenes is a facultative intracellular pathogen of both humans and animals, L. ivanovii mainly infects ungulates (eg., sheep and cattle), while other species (L. innocua, L. seeligeri, L. welshimeri, L. grayi, L. marthii, L. rocourtiae, L. fleischmannii and L. weihenstephanensis) are essentially saprophytes. Within the species of L. monocytogenes, several serovars (e.g., 4b, 1/2a, 1/2b and 1/2c) are highly pathogenic and account for a majority of clinical isolations. Due to their close morphological, biological, biochemical and genetic similarities, laboratory identification of pathogenic and nonpathogenic Listeria organisms is technically challenging. With the development and application of various molecular approaches, accurate and rapid discrimination of pathogenic and nonpathogenic Listeria organisms, as well as pathogenic and nonpathogenic L. monocytogenes strains, has become possible.
Listeria was first described by E.G.D. Murray in 1926 in Cambridge, England, who referred to the causative agent for monocytosis in laboratory rodents as Bacterium monocytogenes. In 1927, a bacterium causing mortality in gerbils was identified in Johannesburg, South Africa, and named Listerella hepatolytica in honor of Joseph Lister, a surgeon who pioneered antiseptic surgery. With the realization that Bacterium monocytogenes and Listerella hepatolytica were in fact the identical bacterium and that the name Listerella had been already taken for a slime mold and a protozoan, the organism was renamed Listeria monocytogenes in 1940.1 In addition to L. monocytogenes,9 other species (L. ivanovii, L. innocua, L. seeligeri, L. welshimeri, L. grayi, L. marthii, L. rocourtiae, L. fleischmannii and L. weihenstephanensis) have since been identified within the genus.
Although L. monocytogenes was implicated in human disease from the late 1920s, it was not until 1979 that the link of this bacterium to serious food-borne listeriosis in humans was established.1 In immunocompetent individuals, L. monocytogenes tends to cause gastrointestinal symptoms that are transient in nature and often disappear within a short period. In the immunocompromised individuals such as pregnant women, neonates, and the elderly, L. monocytogenes infection may lead to severe clinical diseases, with abortion and death being usual outcomes.2,3
Considering their close morphological and biological similarities and their varied pathogenicity, it is important that pathogenic and nonpathogenic Listeria species/L. monocytogenes serovars/strains are correctly identified. Over the years, a number of phenotypic procedures have been developed and used for identification and differentiation of Listeria organisms. However, given their variable performance and slow turnover, phenotypic tests for Listeria diagnosis have been largely superseded by molecular approaches. The purpose of this article is to provide an update on the utility of molecular techniques for the improved determination of pathogenic and nonpathogenic listeriae.
Listeria Classification
The genus Listeria covers a group of Gram-positive, non-spore-forming, rod-shaped bacteria of 0.4–0.5 ±m × 1–1.5 µm in size and between 36–39% in G + C content. Taxonomically, the genus Listeria is classified in the family Listeriaceae, order Bacillales, class Bacilli, phylum Furmicutes, domain Bacteria, kingdom Prokaryotae. Apart from Listeria, the only other genus in the Listeriaceae family is Brochothrix. To date, 10 species are recognized within the genus: L. monocytogenes, L. ivanovii (previously known as L. monocytogenes serotype 5), L. seeligeri, L. innocua, L. welshimeri, L. grayi, L. marthii, L. rocourtiae, L. fleischmannii and L. weihenstephanensis.4–9 Of these, L. monocytogenes is a facultative intracellular pathogen of both humans and animals, L. ivanovii primarily infects ungulates (eg, sheep and cattle), and the other 8 species are free-living saprophytes.10–17 Nonetheless, non-monocytogenes Listeria species including L. ivanovii,18–21L. seeligeri,22L. innocua,23L. welshimeri,24 and L. grayi25–27 have been occasionally implicated in human clinical cases, mainly in individuals with suppressed immune functions and/or underlying illnesses.
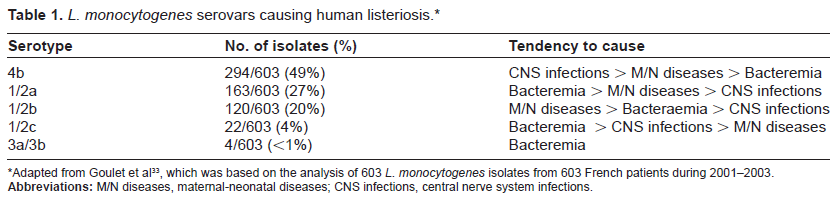
Since L. monocytogenes strains display notable variations in virulence, attempts have been made to develop and use laboratory procedures to differentiate pathogenic from nonpathogenic strains, and to monitor the strains involved in the listeriosis outbreaks. Serotyping on the basis of immunological reactions between listerial somatic (O)/flagellar (H) antigens and specific antibodies represents an early approach to identifying and tracking Listeria bacteria. Using this approach, Listeria is separated into at least 16 serovars, including 13 for L. monocytogenes (serovars 1/2a, 1/2b, 1/2c, 3a, 3b, 3c, 4a, 4ab, 4b, 4c, 4d, 4e and 7), 1 for L. ivanovii (serovar 5), 3 for L. innocua (serovars 1/2b, 6a and 6b), 3 for L. welshimeri (serovars 1/2b, 6a and 6b), 6 for L. seeligeri (serovars 1/2a, 1/2b, 3b, 4a, 4b, 4c and 6b), and 1 for L. grayi (serovar Grayi).28–32 The determination of L. monocytogenes serovars has clinical implications, as serovar 4b strains have been shown to cause endemic human listeriosis, and serovars 1/2a, 1/2b and 1/2c are responsible for sporadic listeriosis in humans. Indeed, according to a French study conducted in 2006, L. monocytogenes serovars 4b, 1/2a, 1/2b and 1/2c account for over 98% isolations from clinical cases of human listeriosis, with serovar 4b alone causing 49% of Listeria-related endemic foodborne diseases (Table 1).33 Similarly, in experimental mouse models, L. monocytogenes serovars 4b, 1/2a, 1/2b and 1/2c show a heightened infectivity through intragastric inoculation.34,35 However, all L. monocytogenes serovars except 4a are capable of inducing mouse mortality via intraperitoneal route.36–39
L. monocytogenes serovars causing human listeriosis.*
Serotype
No. of isolates (%)
Tendency to cause
4b
294/603 (49%)
CNS infections > M/N diseases > Bacteremia
1/2a
163/603 (27%)
Bacteremia > M/N diseases > CNS infections
1/2b
120/603 (20%)
M/N diseases > Bacteraemia > CNS infections
1/2c
22/603 (4%)
Bacteremia > CNS infections > M/N diseases
3a/3b
4/603 (<1%)
Bacteremia
Adapted from Goulet et al33, which was based on the analysis of 603 L. monocytogenes isolates from 603 French patients during 2001–2003.
Abbreviations: M/N diseases, maternal-neonatal diseases; CNS infections, central nerve system infections.
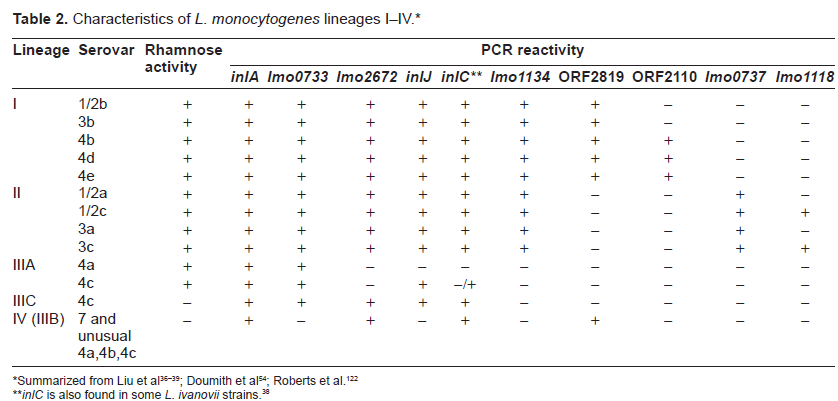
In light of the extensive antigenic sharing among Listeria serovars (e.g., serovars 1/2a and 3a both contain H antigens A and B; serovars 1/2c and 3c both possess H antigens B and D; serovars 1/2b, 3b, 4a, 4b, 4c, 4d, 5, 6a, 6b and 7 all have H antigens A, B, and C; serovars 1/2a.1/2b, 1/2c, 3a, 3b and3c all share O antigen II; serovars 4a, 4ab, 4b, 4c, 4d, 4e, 5, 6a and 6b all have O antigen V), serotyping lacks desired specificity.40,41 As a consequence, genotyping techniques have been developed to improve the identification and epidemiological tracking of Listeria bacteria.42 This has facilitated the separation of L. monocytogenes bacteria into 4 genetic lineages (I–IV) (Table 2).43–50 While lineage I encompasses serovars 1/2b, 3b, 4b, 4d and 4e; lineage II covers serovars 1/2a, 1/2c, 3a and 3c; lineage III includes serovars 4a and 4c. In addition, lineage III has been further distinguished into subgroup IIIA (containing typical rhamnose-positive avirulent serovar 4a and virulent serovar 4c strains), subgroup IIIC (consisting of atypical rhamnose-negative virulent serovar 4c strains), and subgroup IIIB (which is now known as lineage IV) (covering atypical rhamnose-negative, virulent non-serovar 4a and non-serovar 4c, as well as serovar 7 strains).51
Characteristics of L. monocytogenes lineages I–IV.*
Being small, Gram-positive rods, listeriae resemble other Gram-positive bacteria such as streptococci and corynebacteria morphologically. To differentiate the genus Listeria from other bacterial genera, a batch of biochemical tests has been traditionally employed.32,52,53 Recent application of molecular techniques has simplified the genus-specific identification of listeriae, with the following gene targets being commonly exploited:
the house-keeping genes prs and ldh flanking the prs-prfA-plcA-hly-mpl-actA-plcB-orfX-orfZ-orf B-orfA-ldh cluster, which consists of the well known 9.6 kb PrfA-regulated virulence gene cluster (or Listeria pathogenicity island 1, LIPI-1). While the ldh gene codes for lactate dehydrogenase (~310 amino acids), the prs gene encodes phosphoribosyl pyrophosphate synthetase (318 amino acids).54 Additionally, the underlying gene encoding VclB (Lmo0209/Lin0289), a conserved protein of unknown function, is also found in all Listeria species and can be used for Listeria determination.
the 23S rRNA-16S rRNA locus (consisting of about 1500 and 2500 bp, respectively), which is a highly conserved gene region encoding ribosomal RNA molecules (rRNA). The key functions of rRNA are to decode messenger RNA (mRNA) into amino acids and to interact with the transfer RNA (tRNA) during translation by providing petidyltransferase activity. Because of its conserved nature, the 23S rRNA-16S rRNA locus offers a valuable target for phylogenetic analysis.30,55–58 Paillard et al58 employed primers S2F and S2R to generate an 890 bp fragment from the 5’ end of Listeria rRNA 23S gene. Subsequent digestion of this fragment with restriction enzyme XmnI enabled distinction of L. monocytogenes, L. ivanovii and L. seeligeri (forming 770 and 120 bp bands) from L. innocua, L. welshimeri and L. grayi (forming 650, 120 and 120 bp bands). Moreover, digestion of the 890 bp fragment with restriction enzyme CfoI facilitated differentiation of L. ivanovii, L. seeligeri and L. grayi (forming 600, 170 and 120 bp bands) from L. monocytogenes, L. innocua and L. welshimeri (forming 470, 170, 130 and 120 bp bands).
the iap gene. This gene encodes the “invasionassociated protein” (IAP, also known as P60 reflecting its molecular size of 60 kDa), which is involved in host cell invasion by pathogenic listeriae and acts in all Listeria species as a murein hydrolase necessary for proper cell division. The iap gene has been successfully incorporated in PCR for Listeria genus specific detection.59,60
An added benefit of incorporating a genus-specific primer set in a PCR assay for listerial identification lies in the fact that it also functions as an internal control for the assay.
Species-Specific Identification
Correct identification of Listeria species is critical for effective control and prevention of listeriosis. Previously, phenotype-based methods (such as biochemical and serological tests) have been employed for the speciation of Listeria bacteria.61–63 In view of their superior sensitivity and specificity over the phenotypic methods, molecular techniques have been widely adopted in clinical and research laboratories for discrimination between pathogenic and nonpathogenic Listeria organisms.64,65 Evolving from non-amplified procedures (eg., DNA hybridization), molecular detection of Listeria bacteria has increasingly moved towards nucleic acid amplification and real time detection.66–69
The identification of a range of gene targets has further enhanced the appeal and versatility of molecular procedures for Listeria species-specific determination. While several shared genes such as 16S and 23S rRNA genes, their intergenic spacer regions, ssrA gene (which encodes a transfer-messenger RNA or tmRNA), and iap (which encodes invasion associated protein) have proven valuable for identification of all Listeria species,56,58,59,70–77 many Listeria species-specific genes have been described. For instance, the following genes targets may be used for specific determination of L. monocytogenes: hly,78,79plcA,80plcB,80actA,81inlA,82,83inlB,83,84lmaA/lmaB,85flaA,86pepC,87clpE,81fbp,88lmo0733,89 and lmo223490 Similarly, liv22–22891 and smcL92 have been specifically targeted for L. ivanovii, lse24–31593 for L. seeligeri, lin046494 and lin248395 for L. innocua, fbp,88lwe7–57196 and lwe180197 for L. welshimeri, and lgr20–24698 for L. grayi.
Lineage Delineation
As L. monocytogenes lineages I (particularly serovars 1/2b and 4b) and II (especially serovars 1/2a and 1/2c) strains are commonly associated with human clinical cases, it is important that they are accurately identified and subtyped.99–101
Similar to species-specific identification, 2 major approaches are used for L. monocytogenes lineage delineation and subtyping: phenotypic and genetic.102 The phenotypic subtyping approach utilizes serotyping, phage typing, multilocus enzyme electrophoresis (MLEE) and esterase typing techniques.103 The genetic subtyping approach ranges from pulsed-field gel electrophoresis (PFGE), ribotyping, PCR-based subtyping [e.g., random amplification of polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), PCR-restriction fragment length polymorphism (PCR-RFLP), repetitive element PCR (REP-PCR)], to DNA sequencing-based subtyping techniques [such as multilocus sequence typing (MLST)].30,43–45,55,57,104–118 With its high sensitivity, discriminatory power and reproducibility, the genetic subtyping approach offers a method of choice for the laboratory determination of L. monocytogenes lineages and subtypes. In particular, a combination of 2 or more subtyping techniques helps clarify the ambiguity that can be encountered when a single typing method is used.119–121
Due to their sequence divergences among Listeria serovars, the actA and plcB genes have been often targeted for the determination of L. monocytogenes lineages and genotypes.44,113,120,122 Analyses of 2 house-keeping genes (ribC and purM) together with 2 virulence genes (actA and inlA) uncovered evidence of a more prevalent recombination in lineage II than in lineage I.123 Moreover, comparisons of the actA gene sequences of L. seeligeri isolates from different habitats permitted discrimination of 2 different actA subtypes forming 2 phylogenetic lineages.124
Another important group of gene targets for Listeria lineage determination is internalin genes. Through sequencing analysis of the ascB-dapE internalin cluster, Chen et al125 showed that L. monocytogenes lineage II can be distinguished into 3 distinct sublineages, IIA, IIB, and IIC, with inlGHE, inlGC2DE, and inlC2DE for IIA, IIB, and IIC, respectively. While IIA and IIC displayed a higher frequency of recombination, IIB was more notably affected, leading to high nucleotide diversity. Furthermore, internalin profiling of 13 L. monocytogenes lineage III strains identified 10 internalin types that are clustered in 4 subpopulations IIIA–1, IIIA–2, HIB, and IIIC. Whereas lineage IIIA–2 strains had reduced pathogenicity, the other lineage III strains had comparable virulence to lineages I and II. Because of its phylogenetical distinction from other sub-populations, HIB may represent a novel lineage.126 Similarly, examination of internalin genes of L. innocua resulted in the identification of 4 subgroups within the species.127
Group-Specific Identification
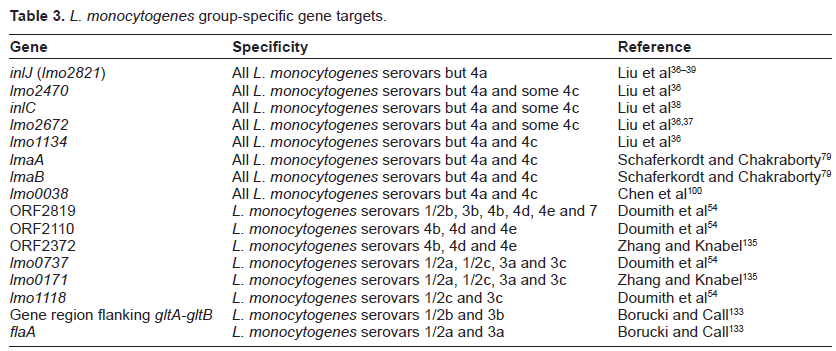
Given the predominance of L. monocytogenes serovars 4b, 1/2a, 1/2c and 1/2b in human clinical isolations, the availability of methods to determine the serotype of a particular strain is vital for its epidemiological tracking and therapeutic monitoring (Table 3). Although conventional serotyping methods have played a valuable role in the tracking of L. monocytogenes isolates involved in listeriosis, they are sometimes unable to correlate serovars directly with species identities, and are expensive to set up and maintain.40,128,129 Without these obvious shortcomings, molecular techniques provide a precise and low-cost alternative for determination of L. monocytogenes serovars/groups.130,131
Jinneman and Hill132 reported a mismatch amplification mutation assay (MAMA) targeting a 446-bp region within the hly gene for rapid screening and characterization of L. monocytogenes lineage types I–III. Borucki and Call133 utilized primers from an iron transport protein gene, GLT primers (from a 1/2b serotype-specific region flanking the gltA-gltB cassette), the MAMA-C PCR primers,132 and primers from the flaA gene (encoding the L. monocytogenes flagellin protein) to identify L. monocytogenes serotypes. Doumith et al54,134 developed a multiplex PCR that incorporates L. monocytogenes lmo0737 gene primers for recognition of serovars 1/2a, 1/2c, 3a, and 3c; lmo1118 gene primers for detection of serovars 1/2c and 3c; ORF2819 primers for serovars 1/2b, 3b, 4b, 4d, and 4e; ORF2110 primers for serovars 4b, 4d, and 4e; and prs primers as an internal amplification control covering all L. monocytogenes serovars. Zhang and Knabel135 described a multiplex PCR assay for rapid identification and easily interpretable differentiation of serovars 1/2a and 4b from other serovars of L. monocytogenes by simultaneously targeting 2 virulence genes (inlB and inlC) and 2 serovar-specific genes (ORF2372 and lmo0171). Nightingale et al136 combined a multiplex PCR with sigB allelic typing to classify the 4 major serovars (i.e., 1/2a, 1/2b, 1/2c, and 4b) into unique genetic subgroups, and to differentiate lineage I serovar 4b isolates from the genetically distinct lineage III serovar 4b isolates. More recently, Kérouanton et al137 designed 2 multiplex PCR assays to cluster L. monocytogenes strains into 5 molecular serogroups: IIa, IIb, IIc, IVa, and IVb. The first multiplex PCR recognizes L. monocytogenes serotypes 1/2a, 1/2c, 1/2b and 4b, together with the prfA gene primers for L. monocytogenes species confirmation. The second multiplex PCR incorporating the flaA gene primers (specific for 1/2a and 3a strains) and prs gene primers (specific for Listeria genus) resolves a small number of IIa and IIc molecular serogroup strains (consisting of serotypes atypical 1/2a, 3a and 1/2c strains) that give equivocal results in the first multiplex PCR, leading to a total agreement between molecular and conventional serotyping methods.
In addition, by using primers from inlA for species-specific recognition, and those from inlJ (or lmo2821) and inlC for virulence determination in a multiplex PCR, L. monocytogenes naturally avirulent serovar 4a strains were rapidly differentiated from other serovars that have the potential to cause mouse mortality via the intraperitoneal route.37
Identification of Epidemic Clones
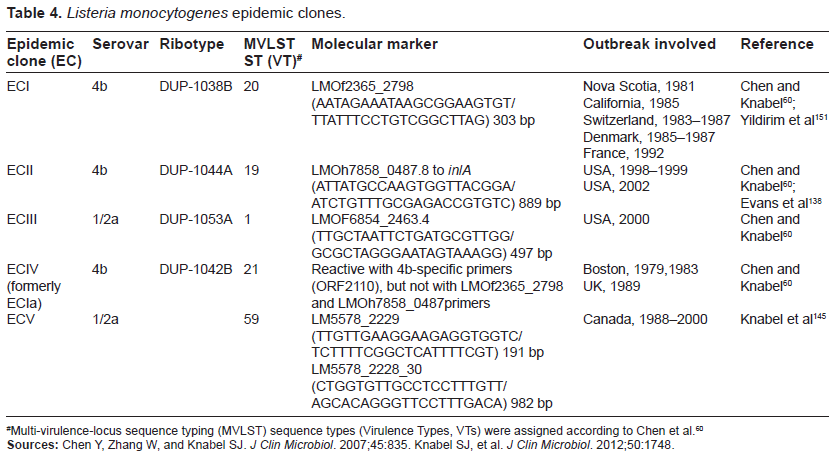
Although a variety of L. monocytogenes strains have been isolated from environments and foodstuff, only a limited number of virulent strains are known to cause listeriosis epidemics, particularly of those belonging to serovars 4b, 1/2a, and 1/2b.138–141 The term “epidemic clone” refers to a group of genetically related isolates of a common ancestor that are implicated in geographically and temporally unrelated outbreaks.142 To date, 5 epidemic clones (ECs) of L. monocytogenes (ECI, ECII, ECIII, ECIV, and ECV) have been defined (Table 4).60,142–146
Multi-virulence-locus sequence typing (MVLST) sequence types (Virulence Types, VTs) were assigned according to Chen et al.60
Sources: Chen Y, Zhang W, and Knabel SJ. J Clin Microbiol. 2007;45:835. Knabel SJ, et al. J Clin Microbiol. 2012;50:1748.
Identification and tracking of L. monocytogenes epidemic clones are critical to understanding the long-term transmission of L. monocytogenes and to establishing efficient surveillance systems for this pathogen.147–149 The methods for the identification of L. monocytogenes epidemic clones have evolved over the years from the phenotypic (e.g., serotyping and phage typing) to genotypic methods.150–153 The latter include the fragment-based typing methods, which range from (i) restriction digestion-based methods such as ribotyping (RT) and pulsed-field gel electrophoresis (PFGE) and (ii) PCR-based methods such as randomly amplified polymorphic DNA and repetitive sequence-based PCR to (iii) combined amplification-restriction methods such as amplified fragment length polymorphism (at endonuclease restriction or primer annealing sites) and PCR-restriction fragment length polymorphism. This was followed by DNA sequence-based methods such as multilocus sequence typing (MLST) that combines PCR and automated DNA sequencing to analyze slowly diversified house-keeping gene sequences.154 More recently, multi-virulence-locus sequence typing (MVLST, targeting virulence genes prfA, inlB, inlC, dal, clpP, and lisR) was developed to overcome the limited discriminatory power associated with MLST, allowing categorization of L. monocytogenes isolates into higher-level groups, such as evolutionary lineages, clonal complexes, and epidemic clones.90,155 Indeed, Knabel et al145 employed multilocus sequence typing (MLST) and multi-virulence-locus sequence typing (MVLST) to identify a predominant clone (clonal complex 8; virulence type 59; proposed epidemic clone 5 [ECV]) belonging to serotype 1/2a that has caused human illness across Canada for more than 2 decades.
To further streamline the identification of L. monocytogenes epidemic clones, Chen and Knabel60 developed a multiplex PCR assay that facilitated simultaneous detection of Listeria genus, L. monocytogenes serovar 1/2a and 4b, and L. monocytogenes epidemic clones I, II, and III. This multiplex PCR assay offers a powerful tool to screen and subgroup L. monocytogenes cultures and significantly reduces the number of isolates that need to be subtyped by more expensive and discriminatory molecular methods, such as PFGE and sequence-based typing.
Conclusion
The genus Listeria contains 10 closely related Gram-positive bacterial species with ubiquitous distribution. Although a majority of Listeria species are nonpathogenic, L. monocytogenesis is a well known pathogen of both humans and animals, and L. ivanovii causes severe diseases in ungulates. For the epidemiological tracking and control of listeriosis outbreaks, it is important to distinguish beween pathogenic and nonpathogenic Listeria species, as well as between pathogenic and nonpathogenic L. monocytogenes strains. While traditional phenotypic methods have contributed to the identification and detection of Listeria organisms in the past, they are largely overtaken by new generation molecular techniques that demonstrate superior sensitivity, specificity and speed. It is envisaged that continuing innovations such as microarrays, biosensors, and next generation sequencing will offer promise to further improve the sensitivity, rapidity and specificity of laboratory characterization of Listeria genus, species, lineages, serovars and epidemic clones.
Author Contributions
Conceived and designed the experiments: DL. Analyzed the data: DL. Wrote the first draft of the manuscript: DL. Contributed to the writing of the manuscript: DL. Agree with manuscript results and conclusions: DL. Jointly developed the structure and arguments for the paper: DL. Made critical revisions and approved final version: DL. All authors reviewed and approved of the final manuscript.
Funding
Author(s) disclose no funding sources for this work.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the author has provided signed confirmation of compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.
References
1.
McCarthyS.A.Listeria in the Environment. In: Foodborne Listeriosis.MillerA.J., SmithA.K., SomkutiG.A. (Eds). 1990; Elsevier: New York; 25–29.
DoganayM.Listeriosis: clinical presentation. FEMS Immunol Med Microbiol.2003; 35(3): 173–175.
4.
LiuD.Identification, subtyping and virulence determination of Listeria monocytogenes, an important foodborne pathogen. J Med Microbiol.2006; 55(6): 645–659.
5.
GravesL.M., HelselL.O., SteigerwaltA.G.Listeria marthii sp. nov., isolated from the natural environment, Finger Lakes National Forest. Int J Syst Evol Microbiol.2010; 60(6): 1280–1288.
BertschD., RauJ., EugsterM.R.Listeria fleischmannii sp. nov., isolated from cheese. Int J Syst Evol Microbiol.2013; 63(2): 526–532.
8.
den BakkerH.C., ManuelC.S., FortesE.D., WiedmannM., NightingaleK.K.Genome sequencing identifies Listeria fleischmannii subsp. coloradensis subsp. nov., a novel Listeria fleischmannii subspecies isolated from a ranch in Colorado. Int J Syst Evol Microbiol.2013, Mar 22. [Epub ahead of print].
9.
Lang HalterE., NeuhausK., SchererS.Listeria weihenstephanensis sp. nov., isolated from the water plant Lemna trisulca taken from a freshwater pond. Int J Syst Evol Microbiol.2013; 63(2): 641–647.
10.
GlaserP., FrangeulL., BuchrieserC.Comparative genomics of Listeria species. Science.2001; 294(5543): 849–852.
11.
BuchrieserC., RusniokC., KunstF., CossartP., GlaserP; Listeria Consortium. Comparison of the genome sequences of Listeria monocytogenes and Listeria innocua: clues for evolution and pathogenicity. FEMS Immunol Med Microbiol.2003; 35(3): 207–213.
12.
BuchrieserC., RusniokC., GarridoP.Complete genome sequence of the animal pathogen Listeria ivanovii, which provides insights into host specificities and evolution of the genus Listeria. J Bacteriol.2011; 193(23): 6787–6788.
13.
NelsonK.E., FoutsD.E., MongodinE.F.Whole genome comparisons of serotype 4b and 1/2a strains of the food-borne pathogen Listeria monocytogenes reveal new insights into the core genome components of this species. Nucleic Acids Res.2004; 32: 2386–2395.
14.
SchmidM.W., NgE.Y.W., LampidisR.Evolutionary history of the genus Listeria and its virulence genes. Syst Appl Microbiol.2005; 28: 1–18.
15.
HainT., SteinwegC., ChakrabortyT.Comparative and functional genomics of Listeria spp. J Biotechnol.2006; 126(1): 37–51.
16.
HainT., GhaiR., BillionA.Comparative genomics and transcriptomics of lineages I, II, and III strains of Listeria monocytogenes. BMC Genomics.2012; 13: 144.
17.
SteinwegC., KuenneC.T., BillionA.Complete genome sequence of Listeria seeligeri, a nonpathogenic member of the genus Listeria. J Bacteriol.2010; 192: 1473–1474.
18.
CumminsA.J., FieldingA.K., McLauchlinJ.Listeria ivanovii infection in a patient with AIDS. J Infect.1994; 28(1): 89–91.
GuilletC., Join-LambertO., Le MonnierA.Human listeriosis caused by Listeria ivanovii. Emerg Infect Dis.2010; 16(1): 136–138.
22.
RocourtJ., HofH., SchrettenbrunnerA., MalinverniR., BilleJ.[Acute purulent Listeria seelingeri meningitis in an immunocompetent adult]. Schweiz Med Wochenschr.1986; 116(8): 248–251. French.
23.
PerrinM., BemerM., DelamareC.Fatal case of Listeria innocua bacteremia. J Clin Microbiol.2003; 41(11): 5308–5309.
24.
AndreP., GenicotA.First isolation of Listeria welshimeri from human beings. Zentbl. Bakteriol. Parasitenkd. Infektkrankh. Hyg Abt I Orig Reihe A.1987; 263: 605–606.
25.
TodeschiniG., FrisoS., LombardiS., CasarilM., FontanaR., CorrocherR.A case of Listeria murray/grayi bacteremia in a patient with advanced Hodgkin's disease. Eur J Clin Microbiol Infect Dis.1998; 17(11): 808–810.
26.
RaposeA., LickS.D., IsmailN.Listeria grayi bacteremia in a heart transplant recipient. Transpl Infect Dis.2008; 10(6): 434–436.
27.
SalimniaH., PatelD., LephartP.R., FairfaxM.R., ChandrasekarP.H.Listeria grayi: vancomycin-resistant, gram-positive rod causing bacteremia in a stem cell transplant recipient. Transpl Infect Dis.2010; 12(6): 526–528.
28.
SeeligerH.P.R., HöhneK.Serotyping of Listeria monocytogenes and related species. Methods Microbiol.1979; 13: 31–49.
29.
SeeligerH.P.R., JonesD.Listeria. In: Bergey's Manual of Systematic Bacteriology, Vol. 2.SneathP.H.A. (Eds). 1986; Baltimore: Williams and Wilkins; 1235–1245.
30.
VaneechoutteM., BoerlinP., TichyH.V., BannermanE., JagerB., BilleJ.Comparison of PCR-based DNA fingerprinting techniques for the identification of Listeria species and their use for atypical Listeria isolates. Int J Syst Bacteriol.1998; 48: 127–139.
31.
PalumboJ.D., BoruckiM.K., MandrellR.E., GorskiL.Serotyping of Listeria monocytogenes by enzyme-linked immunosorbent assay and identification of mixed-serotype cultures by colony immunoblotting. J Clin Microbiol.2003; 41: 564–571.
32.
GorskiL.Phenotypic identification. In: LiuD. (Ed). Handbook of Listeria monocytogenes.2008; Boca Raton: Taylor and Francis CRC Press; 139–168.
33.
GouletV., JacquetC., MartinP., VaillantV., LaurentE., de ValkH.Surveillance of human listeriosis in France, 2001–2003. Euro Surveill.2006; 11: 79–81.
34.
BarbourA.H., RamplingA., HormaecheC.E.Variation in the infectivity of Listeria monocytogenes isolates following intragastric inoculation of mice. Infect Immun.2001; 69: 4657–4660.
35.
KimS.H., BakkoM.K., KnowlesD., BoruckiM.K.Oral inoculation of A/J mice for detection of invasiveness differences between Listeria monocytogenes epidemic and environmental strains. Infect Immun.2004; 72: 4318–4321.
36.
LiuD., AinsworthA.J., AustinF.W., LawrenceM.L.Characterization of virulent and avirulent Listeria monocytogenes strains by PCR amplification of putative transcriptional regulator and internalin genes. J Med Microbiol.2003; 52(12): 1065–1070.
SchönbergA., BannermanE., CourtieuA.L.Serotyping of 80 strains from the WHO multicentre international typing study of Listeria monocytogenes. Int J Food Microbiol.1996; 32(3): 279–287.
41.
LiuD., LawrenceM., GorskiL.Listeria monocytogenes serotype 4b strains belonging to lineages I and III possess distinct molecular features. J Clin Microbiol.2006; 44: 214–217.
42.
OrsiR.H., den BakkerH.C., WiedmannM.Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int J Med Microbiol.2011; 301(2): 79–96.
43.
RasmussenO.F., SkouboeP., DonsL., RossenL., OlsenJ.E.Listeria monocytogenes exists in at least three evolutionary lines: Evidence from flagellin, invasive associated protein and listeriolysin O genes. Microbiology.1995; 141: 2053–2061.
44.
WiedmannM., BruceL.J., KeatingC., JohnsonA.E., McDonoughP.L., BattC.A.Ribotypes and virulence gene polymorphisms suggest three distinct Listeria monocytogenes lineages with differences in pathogenic potential. Infect Immun.1997; 65: 2707–2716.
45.
KerouantonA., BrisaboisA., DenoyerE.Comparison of five typing methods for the epidemiological study of Listeria monocytogenes. Int J Food Microbiol.1998; 43: 61–71.
46.
DuceyT.F., PageB., UsgaardT.A single-nucleotide-polymorphism-based multilocus genotyping assay for subtyping lineage I isolates of Listeria monocytogenes. Appl Environ Microbiol.2007; 73: 133–147.
47.
MeinersmannR.J., PhillipsR.W., WiedmannM., BerrangM.E.Multilocus sequence typing of Listeria monocytogenes by use of hypervariable genes reveals clonal and recombination histories of three lineages. Appl Environ Microbiol.2004; 70: 2193–2203.
48.
SaudersB.D., SchukkenY., KornsteinL.Molecular epidemiology and cluster analysis of human listeriosis cases in three U.S. states. J Food Prot.2006; 69(7): 1680–1689.
49.
SaudersB.D., OverdevestJ., FortesE.Diversity of Listeria species in urban and natural environments. Appl Environ Microbiol.2012; 78(12): 4420–4433.
50.
TsaiY.H., MaronS.B., McGannP., NightingaleK.K., WiedmannM., OrsiR.H.Recombination and positive selection contributed to the evolution of Listeria monocytogenes lineages III and IV, two distinct and well supported uncommon L. monocytogenes lineages. Infect Genet Evol.2011; 11(8): 1881–1890.
51.
WardT.J., DuceyT.F., UsgaardT., DunnK.A., BielawskiJ.P.Multilocus genotyping assays for SNP-based subtyping of Listeria monocytogenes. Appl Environ Microbiol.2008; 74(24): 7629–7642.
52.
BilleJ., CatimelB., BannermanE.API Listeria, a new and promising one-day system to identify Listeria isolates. Appl Environ Microbiol.1992; 58: 1857–1860.
53.
HarveyJ., GilmourA.Characterization of Listeria monocytogenes isolates by esterase electrophoresis. Appl Environ Microbiol.1996; 62: 1461–1466.
54.
DoumithM., BuchrieserC., GlaserP., JacquetC., MartinP.Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J Clin Microbiol.2004; 42: 3819–3822.
55.
BruceJ.L., HubnerR.J., ColeE.M., McDowellC.I., WebsterJ.A.Sets of EcoRI fragments containing ribosomal RNA sequences are conserved among different strains of Listeria monocytogenes. Proc Natl Acad Sci USA.1995; 92: 5229–5233.
56.
SallenB., RajoharisonA., DesvarenneS., QuinnF., MabilatC.Comparative analysis of 16S and 23S rRNA sequences of Listeria species. Int J Syst Bacteriol.1996; 46(3): 669–674.
57.
De CesareA., BruceJ.L., DambaughT.R., GuerzoniM.E., WiedmannM.Automated ribotyping using different enzymes to improve discrimination of Listeria monocytogenes isolates, with a particular focus on serotype 4b strains. J Clin Microbiol.2001; 39: 3002–3005.
58.
PaillardD., DuboisV., DuranR.Rapid identification of Listeria species by using restriction fragment length polymorphism of PCR-amplified 23S rRNA gene fragments. Appl Environ Microbiol.2003; 69: 6386–6392.
59.
BubertA., KöhlerS., GoebelW.The homologous and heterologous regions within the iap gene allow genus- and species-specific identification of Listeria spp. by polymerase chain reaction. Appl Environ Microbiol.1992; 58: 2625–2632.
60.
ChenY., KnabelS.J.Multiplex PCR for simultaneous detection of bacteria of the genus Listeria, Listeria monocytogenes, and major serotypes and epidemic clones of L. monocytogenes. Appl Environ Microbiol.2007; 73(19): 6299–6304.
61.
DonnellyC.W.Historical perspectives on methodology to detect Listeria monocytogenes. J Assoc Off Anal Chem.1988; 71(3): 644–646.
62.
RocourtJ., CatimelB.Biochemical characterization of species in the genus Listeria. Zentralbl Bakteriol Mikrobiol Hyg A.1985; 260: 221–231.
63.
LoessnerM.J.Improved procedure for bacteriophage typing of Listeria strains and evaluation of new phages. Appl Environ Microbiol.1991; 57: 882–884.
64.
AznarR., AlarconB.On the specificity of PCR detection of Listeria monocytogenes in food: a comparison of published primers. Syst Appl Microbiol.2002; 25: 109–119.
65.
HuangB., EglezosS., HeronB.A.Comparison of multiplex PCR with conventional biochemical methods for the identification of Listeria spp. isolates from food and clinical samples in Queensland, Australia. J Food Prot.2007; 70: 1874–1880.
66.
KlingerJ.D., JohnsonA., CroanD.Comparative studies of nucleic acid hybridization assay for Listeria in foods. J Assoc Off Anal Chem.1988; 71: 669–673.
67.
KöhlerS., Leimeister-WächterM., ChakrabortyT., LottspeichF., GoebelW.The gene coding for protein p60 of Listeria monocytogenes and its use as a specific probe for Listeria monocytogenes. Infect Immun.1990; 58(6): 1943–1950.
68.
RattanachaikunsoponP., PhumkhachornP.Identification of viable Listeria species based on reverse transcription-multiplex PCR (RT-MPCR) and restriction digestion. Biosci Biotechnol Biochem.2012; 76(6): 1189–1194.
69.
LaksanalamaiP., JacksonS.A., MammelM.K., DattaA.R.High density microarray analysis reveals new insights into genetic footprints of Listeria monocytogenes strains involved in listeriosis outbreaks. PLoS ONE.2012; 7(3): e32896.
70.
BubertA., HeinI., RauchM.Detection and differentiation of Listeria spp. by a single reaction based multiplex PCR. Appl Environ Microbiol.1999; 65: 4688–4692.
71.
WangR.F., CaoW.W., JohnsonM.G.16S rRNA-based probes and polymerase chain reaction method to detect Listeria monocytogenes cells added to foods. Appl Environ Microbiol.1992; 58, 2827–2831.
72.
GrahamT., Golsteyn-ThomasE.J., GannonV.P., ThomasJ.E.Genus- and species-specific detection of Listeria monocytogenes using polymerase chain reaction assays targeting the 16S/23S intergenic spacer region of the rRNA operon. Can J Microbiol.1996; 42: 1155–1162.
73.
GrahamT.A., Golsteyn-ThomasE.J., ThomasJ.E., GannonV.P.Inter- and intraspecies comparison of the 16S–23S rRNA operon intergenic spacer regions of six Listeria spp. Int J Syst Bacteriol.1997; 47: 863–869.
74.
ManzanoM., CocolinL., CantoniC., ComiG.Temperature gradient gel electrophoresis of the amplified product of a small 16S rRNA gene fragment for the identification of Listeria species isolated from food. J Food Prot.2000; 63: 659–661.
75.
SomerL., KashiY.A PCR method based on 16S rRNA sequence for simultaneous detection of the genus Listeria and the species Listeria monocytogenes in food products. J Food Prot.2003; 66: 1658–1665.
76.
WangJ., YamadaS., OhashiE.Rapid identification of Listeria species and screening for variants by melting curve and high-resolution melting curve analyses of the intergenic spacer region of the rRNA gene. Can J Microbiol.2010; 56(8): 676–682.
77.
JinD., LuoY., ZhangZ.Rapid molecular identification of Listeria species by use of real-time PCR and high-resolution melting analysis. FEMS Microbiol Lett.2012; 330(1): 72–80.
78.
FurrerB., CandrianU., HoefeleinC., LuethyJ.Detection and identification of Listeria monocytogenes in cooked sausage products and in milk by in vitro amplification of haemolysin gene fragments. J Appl Bacteriol.1991; 70: 372–379.
79.
BlaisB.W., TurnerG., SooknananR., MalekL.T.A nucleic acid sequence-based amplification system for detection of Listeria monocytogenes hlyA sequences. Appl Environ Microbiol.1997; 63: 310–313.
80.
VolokhovD., GeorgeJ., AndersonC., DuvallR.E., HitchinsA.D.Discovery of natural atypical nonhemolytic Listeria seeligeri isolates. Appl Environ Microbiol.2006; 72(4): 2439–2448.
81.
LonghiC., MaffeoA., PentaM., PetroneG., SegantiL., ConteM.P.Detection of Listeria monocytogenes in Italian-style soft cheeses. J Appl Microbiol.2003; 94: 879–885.
82.
PoyartC., Trieu-CuotP., BercheP.The inlA gene required for cell invasion is conserved and specific to Listeria monocytogenes. Microbiology.1996; 142: 173–180.
83.
JungY.S., FrankJ.F., BrackettR.E., ChenJ.Polymerase chain reaction detection of Listeria monocytogenes on frankfurters using oligonucleotide primers targeting the genes encoding internalin AB. J Food Prot.2003; 66: 237–241.
84.
PangalloD., KaclíkováE., KuchtaT., DrahovskáH.Detection of Listeria monocytogenes by polymerase chain reaction oriented to inlB gene. New Microbiol.2001; 24: 333–339.
85.
SchäferkordtS., ChakrabortyT.Identification, cloning, and characterization of the Ima operon, whose gene products are unique to Listeria monocytogenes. J Bacteriol.1997; 179(8): 2707–2716.
86.
GrayD.I., KrollR.G.Polymerase chain reaction amplification of the flaA gene for the rapid identification of Listeria spp. Lett Appl Microbiol.1995; 20: 65–68.
87.
WintersD.K., MaloneyT.P., JohnsonM.G.Rapid detection of Listeria monocytogenes by a PCR assay specific for an aminopeptidase. Mol Cell Probes.1999; 13: 127–131.
88.
GilotP., ContentJ.Specific identification of Listeria welshimeri and Listeria monocytogenes by PCR assays targeting a gene encoding a fibronectinbinding protein. J Clin Microbiol.2002; 40: 698–703.
89.
LiuD., AinsworthA.J., AustinF.W., LawrenceM.L.Use of PCR primers derived from a putative transcriptional regulator gene for species-specific identification of Listeria monocytogenes. Int J Food Microbiol.2004; 91: 297–304.
90.
ZhangW., JayaraoB.M., KnabelS.J.Multi-virulence-locus sequence typing of Listeria monocytogenes. Appl Environ Microbiol.2004; 70(2): 913–920.
91.
LiuD., AinsworthA.J., AustinF.W., LawrenceM.L.PCR detection of a putative N-acetylmuramidase gene from Listeria ivanovii facilitates its rapid identification. Vet Microbiol.2004; 101: 83–89.
92.
Rodríguez-LázaroD., López-EnríquezL., HernándezM.smcL as a novel diagnostic marker for quantitative detection of Listeria ivanovii in biological samples. J Appl Microbiol.2010; 109(3): 863–872.
93.
LiuD., LawrenceM.L., AinsworthA.J., AustinF.W.Species-specific PCR determination of Listeria seeligeri. Res Microbiol.2004; 155: 741–746.
94.
LiuD., AinsworthA.J., AustinF.W., LawrenceM.L.Identification of Listeria innocua by PCR targeting a putative transcriptional regulator gene. FEMS Microbiol Lett.2003; 203: 205–210.
95.
Rodríguez-LázaroD., HernándezM., ScorttiM., EsteveT., Vázquez-BolandJ.A., PlaM.Quantitative detection of Listeria monocytogenes and Listeria innocua by real-time PCR: assessment of hly, iap, and lin02483 targets and AmpliFluor technology. Appl Environ Microbiol.2004; 70: 1366–1377.
96.
LiuD., AinsworthA.J., AustinF.W., LawrenceM.L.Identification of a gene encoding a putative phosphotransferase system enzyme IIBC in Listeria welshimeri and its application for diagnostic PCR. Lett Appl Microbiol.2004; 38: 151–157.
LiuD., LawrenceM.L., AinsworthA.J., AustinF.W.Isolation and PCR amplification of a species-specific, oxidoreductase-coding gene region in Listeria grayi. Can J Microbiol.2005; 51: 95–98.
99.
DoumithM., CazaletC., SimoesN.New aspects regarding evolution and virulence of Listeria monocytogenes revealed by comparative genomics and DNA arrays. Infect Immun.2004; 72: 1072–1083.
100.
ChenJ., JiangL., ChenQ.lmo0038 is involved in acid and heat stress responses and specific for Listeria monocytogenes lineages I and II, and Listeria ivanovii. Foodborne Pathog Dis.2009; 6: 365–376.
101.
DunnK.A., BielawskiJ.P., WardT.J., UrquhartC., GuH.Reconciling ecological and genomic divergence among lineages of listeria under an “extended mosaic genome concept”. Mol Biol Evol.2009; 26(11): 2605–2615.
102.
McLauchlinJ., HamptonM.D., ShahS., ThrelfallE.J., WienekeA.A., CurtisG.D.Subtyping of Listeria monocytogenes on the basis of plasmid profiles and arsenic and cadmium susceptibility. J Appl Microbiol.1997; 83(3): 381–388.
103.
BibbW.F., SchwartzB., GellinB.G., PlikaytisB.D., WeaverR.E.Analysis of Listeria monocytogenes by multilocus enzyme electrophoresis and application of the method to epidemiologic investigations. Int J Food Microbiol.1989; 8: 233–239.
104.
BroschR., ChenJ., LuchanskyJ.B.Pulsed-field fingerprinting of listeriae: identification of genomic divisions for Listeria monocytogenes and their correlation with serovar. Appl Environ Microbiol.1994; 60: 2584–2592.
105.
FarberJ., AddisonC.RAPD typing for distinguishing species and strains in the genus Listeria. J Appl Bacteriol.1994; 77: 242–250.
106.
GravesL.M., SwaminathanB., ReevesM.W.Comparison of ribotyping and multilocus enzyme electrophoresis for subtyping of Listeria monocytogenes isolates. J Clin Microbiol.1994; 32: 2936–2943.
107.
O'DonoghueK., BowkerK., McLauchlinJ., ReevesD.S., BennettP.M., MacGowanA.P.Typing of Listeria monocytogenes by random amplified polymorphic DNA (RAPD) analysis. Int J Food Microbiol.1995; 27: 245–252.
108.
WiedmannM., BruceJ.L., KnorrR.Ribotype diversity of Listeria monocytogenes strains associated with outbreaks of listeriosis in ruminants. J Clin Microbiol.1996; 34: 1086–1090.
109.
LomonacoS., NuceraD., ParisiA., NormannoG., BotteroM.T.Comparison of two AFLP methods and PFGE using strains of Listeria monocytogenes isolated from environmental and food samples obtained from Piedmont, Italy. Int J Food Microbiol.2011; 149: 177–82.
110.
JersekB., GilotP., GubinaM.Typing of Listeria monocytogenes strains by repetitive element sequence-based PCR. J Clin Microbiol.1999; 37(1): 103–109.
111.
RipabelliG., McLauchinJ., ThrelfallE.J.Amplified fragment length polymorphism (AFLP) analysis of Listeria monocytogenes. Syst Appl Microbiol.2000; 23: 132–136.
112.
FranciosaG., TartaroS., Wedell-NeergaardC., AureliP.Characterization of Listeria monocytogenes strains involved in invasive and noninvasive listeriosis outbreaks by PCR-based fingerprinting techniques. Appl Environ Microbiol.2001; 67: 1793–1799.
113.
JeffersG.T., BruceJ.L., McDonoughP.L., ScarlettJ., BoorK.J., WiedmannM.Comparative genetic characterization of Listeria monocytogenes isolates from human and animal listeriosis cases. Microbiology.2001; 147: 1095–1104.
114.
GuerraM.M., BernardoF., McLauchlinJ.Amplified fragment length polymorphism (AFLP) analysis of Listeria monocytogenes. Syst Appl Microbiol.2002; 25: 456–461.
115.
JaradatZ.W., SchutzeG.E., BhuniaA.K.Genetic homogeneity among Listeria monocytogenes strains from infected patients and meat products from two geographic locations determined by phenotyping, ribotyping and PCR analysis of virulence genes. Int J Food Microbiol.2002; 76: 1–10.
116.
Keto-TimonenR.O., AutioT.J., KorkealaH.J.An improved amplified fragment length polymorphism (AFLP) protocol for discrimination of Listeria isolates. Syst Appl Microbiol.2003; 26: 236–244.
117.
ZhangC., ZhangM., JuJ.Genome diversification in phylogenetic lineages I and II of Listeria monocytogenes: identification of segments unique to lineage II populations. J Bacteriol.2003; 185(18): 5573–5584.
118.
MiyaS., TakahashiH., KamimuraC., NakagawaM., KudaT., KimuraB.Highly discriminatory typing method for Listeria monocytogenes using polymorphic tandem repeat regions. J Microbiol Methods.2012; 90(3): 285–291.
WardT.J., GorskiL., BoruckiM.K., MandrellR.E., HutchinsJ., PupedisK.Intraspecific phylogeny and lineage group identification based on the prfA virulence gene cluster of Listeria monocytogenes. J Biotechol.2004; 186: 4994–5002.
121.
WardT.J., UsgaardT., EvansP.A targeted multilocus genotyping assay for lineage, serogroup, and epidemic clone typing of Listeria monocytogenes. Appl Environ Microbiol.2010; 76(19): 6680–6684.
122.
RobertsA., NightingaleK., JeffersG., FortesE., KongoJ.M., WiedmannM.Genetic and phenotypic characterization of Listeria monocytogenes lineage III. Microbiology.2006; 152: 685–693.
123.
den BakkerH.C., DidelotX., FortesE.D., NightingaleK.K., WiedmannM.Lineage specific recombination rates and microevolution in Listeria monocytogenes. BMC Evol Biol.2008; 8: 277.
124.
MüllerA.A., SchmidM.W., MeyerO., MeussdoerfferF.G.Listeria seeligeri isolates from food processing environments form two phylogenetic lineages. Appl Environ Microbiol.2010; 76(9): 3044–3047.
125.
ChenJ., FangC., ZhuN.Genetic organization of ascB-dapE internalin cluster serves as a potential marker for Listeria monocytogenes sublineages IIA, IIB, and IIC. J Microbiol Biotechnol.2012; 22(5): 575–584.
126.
ZhaoH., ChenJ., FangC.Deciphering the biodiversity of Listeria monocytogenes lineage III strains by polyphasic approaches. J Microbiol.2011; 49(5): 759–767.
127.
ChenJ., ChenQ., JiangL.Internalin profiling and multilocus sequence typing suggest four Listeria innocua subgroups with different evolutionary distances from Listeria monocytogenes. BMC Microbiol.2010; 10: 97.
128.
BroschR., BrettM., CatimelB., LuchanskyJ.B., OjeniyiB., RocourtJ.Genomic fingerprinting of 80 strains from the WHO multicenter international typing study of Listeria monocytogenes via pulsed-field gel electrophoresis (PFGE). Int J Food Microbiol.1996; 32: 343–355.
NadonC.A., WoodwardD.L., YoungC., RodgersF.G., WiedmannM.Correlations between molecular subtyping and serotyping of Listeria monocytogenes. J Clin Microbiol.2001; 39: 2704–2707.
131.
DattaA.R., LaksanalamaiP., SolomotisM.Recent developments in molecular sub-typing of Listeria monocytogenes. Food Addit Contam Part A Chem Anal Control Expo Risk Assess.2012, Oct 12. [Epub ahead of print].
132.
JinnemanK.C., HillW.E.Listeria monocytogenes lineage group classification by MAMA-PCR of the listeriolysin gene. Curr Microbiol.2001; 43: 129–133.
DoumithM., JacquetC., Gerner-SmidtP.Multicenter validation of a multiplex PCR assay for differentiating the major Listeria monocytogenes serovars 1/2a, 1/2b, 1/2c and 4b: toward an international standard. J Food Prot.2005; 68(12): 2648–2650.
135.
ZhangW., KnabelS.J.Multiplex PCR assay simplifies serotyping and sequence typing of Listeria monocytogenes associated with human outbreaks. J Food Prot.2005; 68(9): 1907–1910.
136.
NightingaleK., BovellL., GrajczykA., WiedmannM.Combined sigB allelic typing and multiplex PCR provide improved discriminatory power and reliability for Listeria monocytogenes molecular serotyping. J Microbiol Methods.2007; 68(1): 52–59.
137.
KérouantonA., MaraultM., PetitL., GroutJ., DaoT.T., BrisaboisA.Evaluation of a multiplex PCR assay as an alternative method for Listeria monocytogenes serotyping. J Microbiol Methods.2010; 80(2): 134–137.
138.
EvansM.R., SwaminathanB., GravesL.M.Genetic markers unique to Listeria monocytogenes serotype 4b differentiate epidemic clone II (hot dog outbreak strains) from other lineages. Appl Environ Microbiol.2004; 70(4): 2383–2390.
139.
ChengY., SeletzkyR.M., KathariouS.Genomic divisions/lineages, epidemic clons, and population structure. In: LiuD. (Ed). Handbook of Listeria monocytogenes.2008; Boca Raton: CRC Press; 337–357.
140.
Chenal-FrancisqueV., LopezJ., CantinelliT.Worldwide distribution of major clones of Listeria monocytogenes. Emerg Infect Dis.2011; 17(6): 1110–1112.
141.
KathariouS.Listeria monocytogenes virulence and pathogenicity, a food safety perspective. J Food Prot.2002; 65: 1811–1829.
142.
ChenY., ZhangW., KnabelS.J.Multi-virulence-locus sequence typing identifies single nucleotide polymorphisms which differentiate epidemic clones and outbreak strains of Listeria monocytogenes. J Clin Microbiol.2007; 45(3): 835–846.
143.
ChenY., KnabelS.J.Prophages in Listeria monocytogenes contain single-nucleotide polymorphisms that differentiate outbreak clones within epidemic clones. J Clin Microbiol.2008; 46(4): 1478–1484.
144.
ChenY., KumarN., SiddiqueN.Development and evaluation of a real-time polymerase chain reaction assay targeting iap for the detection of Listeria monocytogenes in select food matrices. Foodborne Pathog Dis.2011; 8(10): 1063–1069.
145.
KnabelS.J., ReimerA., VergheseB.Canadian Public Health Laboratory Network (CPHLN). Sequence typing confirms that a predominant Listeria monocytogenes clone caused human listeriosis cases and outbreaks in Canada from 1988 to 2010. J Clin Microbiol.2012; 50(5): 1748–1751.
146.
Chenal-FrancisqueV., DiancourtL., CantinelliT.Optimized Multilocus Variable-Number Tandem-Repeat Analysis Assay and Its Complementarity with Pulsed-Field Gel Electrophoresis and Multilocus Sequence Typing for Listeria monocytogenes Clone Identification and Surveillance. J Clin Microbiol.2013; 51(6): 1868–1880.
147.
LomonacoS., KnabelS.J., DalmassoA., CiveraT., BotteroM.T.Novel multiplex single nucleotide polymorphism-based method for identifying epidemic clones of Listeria monocytogenes. Appl Environ Microbiol.2011; 77(17): 6290–6294.
148.
LomonacoS., PattiR., KnabelS.J., CiveraT.Detection of virulence-associated genes and epidemic clone markers in Listeria monocytogenes isolates from PDO Gorgonzola cheese. Int J Food Microbiol.2012; 160(1): 76–79.
149.
LomonacoS., VergheseB., Gerner-SmidtP.Novel epidemic clones of Listeria monocytogenes, United States, 2011. Emerg Infect Dis.2013; 19(1): 147–150.
150.
PiffarettiJ.C., KressebuchH., AeschbacherM.Genetic characterization of clones of the bacterium Listeria monocytogenes causing epidemic disease. Proc Natl Acad Sci USA.1989; 86(10): 3818–3822.
151.
YildirimS., LinW., HitchinsA.D.Epidemic clone I-specific genetic markers in strains of Listeria monocytogenes serotype 4b from foods. Appl Environ Microbiol.2004; 70(7): 4158–4164. Erratum in: Appl Environ Microbiol. 2004; 70(12): 7581.
152.
FranciosaG., ScalfaroC., MauglianiA.Distribution of epidemic clonal genetic markers among Listeria monocytogenes 4b isolates. J Food Prot.2007; 70(3): 574–581.
153.
LomonacoS., ChenY., KnabelS.J.Analysis of additional virulence genes and virulence gene regions in Listeria monocytogenes confirms the epidemiologic relevance of multi-virulence-locus sequence typing. J Food Prot.2008; 71(12): 2559–2566.
154.
den BakkerH.C., FortesE.D., WiedmannM.Multilocus sequence typing of outbreak-associated Listeria monocytogenes isolates to identify epidemic clones. Foodborne Pathog Dis.2010; 7(3): 257–265.
155.
HuangB., FangN., DimovskiK., WangX., HoggG., BatesJ.Observation of a new pattern in serogroup-related PCR typing of Listeria monocytogenes 4b isolates. J Clin Microbiol.2011; 49(1): 426–429.