Bacillus sp strain IBA 33 metabolites, isolated from decaying lemon fruits, were evaluated for the control of pathogenic and non-pathogenic fungi (Penicillium digitatum, Geotrichum candidum, Penicillium expansum, Aspergillus clavatus, Aspergillus flavus, Aspergillus niger, and Fusarium moniliforme). These metabolites were recovered from Landy medium (LM) without aminoacids. In order to optimize metabolites production the LM was modified by adding different concentrations and sources of amino acids and carbohydrates at different culture conditions.
Bacillus sp strain IBA 33 metabolites efficacy to control fungi were evaluated with in vitro and in vivo assays. A. flavus growth inhibition was 52% with the metabolites of Bacillus sp strain IBA 33 recovered from LM (MBLM) in vitro assays. MBLM supplemented with 0.5% glutamic acid, inhibited the growth of P. digitatum, G. candidum, A. clavatus, A. niger and F. moniliforme by 65%, 88.44%, 84%, 34% and 92% respectively. The highest inhibition of P. expansum was 45% with MBLM supplemented with 0.5% aspartic acid. Similar results were obtained in vivo assays. These results showed that Bacillus sp strain IBA 33 metabolites specificity against fungi depended on the composition of the LM.
One of the greatest causes of loss in the food industry is postharvest diseases of fruits and vegetables.1 According to U.S.A. estimations this loss reaches 24%2 whereas it may climb to 45%–50% in developing countries,3 but loss has generally been considered to be approximately 10% to 40% depending on packinghouse technology.4
Fungi are the principal decaying agents in fruit kept in cold storage chambers for long periods.5
Postharvest lemon diseases are mainly caused by Penicillium italicum (blue mold), Penicillium digitatum (green mold), Geotrichum candidum (sour rot), Alternaria citri (black mold) and Fusarium sp.6
Postharvest fruit diseases are controlled with careful manipulation practices and synthetic fungicides like 2–4 thiazalil benzimidazole and imidazole. This method is more widely used against fungal decay because of its low cost and easy application. However, it presents manifold objections since prolonged use generates resistance to synthetic fungicides by major postharvest pathogens7,8 and increases chemical remainders in fruits with the consequent potentiality engendering iatrogenic diseases.9 In addition, there is strong pressure from the consumers who demand products of greater quality, but with a smaller amount of chemical agents and a major environmental protection.10 A desirable alternative to the use of synthetic fungicides is the application of a biological control of phytopathogenic fungi.
There are many microorganisms like Pseudomona, Gliocladium, Trichoderma, Criptococcus, Rhodotorula and Bacillus11 with antagonistic activity against phytopathogenic fungi.
Currently there are several formulations of Bacillus subtilis in the market to eradicate postharvest diseases of rice, beans, tomato and avocado pears,11 but there are no data about biological control of lemon phytopathogenic fungi using microorganisms of the region where such citric fruits are grown.
In this work we studied the production and optimization of metabolites by a strain of Bacillus sp IBA 33 with antifungal activity and their effectiveness in controlling decaying lemon fruit fungi. The microorganism was isolated from the soil of lemon plantations of Tucumán, Argentina.
We also carried out in vivo studies to determine if the in vitro effect reappeared when the metabolites were applied to the lemons.
Materials and Methods
Microorganisms
Bacterial strain
The Bacillus strain used in this paper was isolated from the soil of a lemon plantation of Tucumán, Argentina and maintained in bevel tubes with nutrient agar pH 7.2. The microorganism was taxonomically identified in genus by phenotypic tests (Gram stain, morphology, catalase activity, nitrate reduction, indole production, motility, growth at 7% NaCl, acid production, gelatinase and starch hidrolysis). The bacilli identification was performed by the standard tests described in the Bergey's Manual of Determinative Bacteriology12 Microscopic disposition, growth at 65 °C were assayed to identify this group.
Genomic DNA from Bacillus strain IBA 33 was isolated, from LB cultures, using a lysozyme method for Gram positive bacteria.13 Bacterial 16S rRNA was amplified by polymerase chain reaction (PCR) from DNA extracted from cultured cells. The primers used were PLB (5′-AGAGTTTGATCCTGGCTCAG-3′) and MLB (5′-GGCTGCTGGCACGTAGTTAG-3′) (The Great American Gene Company, Ramona, CA). PCR contents for a 50 μl volume were 0.3 mg/ml template DNA, 10 μM forward (PLB) and reverse (MLB) primers, 2.5 mM deoxynucleoside triphosphates, 5 μl of 10 × Taq polymerase buffer (50 mM KCl, 10 mM Tris HCl pH 8.3), and 5 U/μl of Taq DNA polymerase (Promega, Madison, WI). The thermal cycling conditions used were 1 min denaturation step at 94 °C, then 1 min at 94 °C, 1 min at 50 °C and 30 cycles at 72 °C for 1 min, followed by a final 7 min extension at 72 °C. All reactions were carried out in a tetrad thermal cycler (Gene Cycler, Bio-Rad). All PCR products were purified with the PCR-Clean up gel extraction MACHEREYNAGEL. PCR products were examined by electrophoresis in a 1% (wt/vol) agarose gel visualized by staining with SYBR Safe DNA Gel Stain (Invitrogen) on a UV transilluminator (Consort, B2300) (Fig. 1) and sequenced in (Ruralex/Fagos, Buenos Aires, Argentina). The obtained sequence was identified through the Genbank database by using the BLAST algorithm.14
Agarose gel electrophoresis of PCR products obtained from DNA extracted from Bacillus sp. IBA 33 cells (Lines 1 and 2). Line 3 DNA size markers (1 Kb DNA Ladder, Gibco BRL).
Fungi strains
Phytopathogenic and saprophytic fungi Penicillium digitatum, Geotrichum candidum, Penicillium expansum, Aspergillus clavatus, Aspergillus flavus, Aspergillus niger and Fusarium moniliforme were isolated from ill lemons harvested in the Tucumán citric zone, identified15 and belong to the stock strains of our Institute. They were grown in bevel tubes with potato dextrose agar pH 5.0 (PDA) at 28 °C and stored on the same medium at 4 °C.
Culture medium and growth conditions of Bacillus sp strain IBA 33 for the production of antifungal metabolites
Growth kinetic of Bacillus sp strain IBA 33
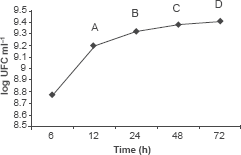
25 ml of Bacillus sp strain IBA 33 activated inoculum (108 CFU ml-1) were placed in 250 ml of nutrient broth pH 7.2 and incubated in a rotary shaker (250 rpm) at 28 °C and sampled every 3 h. Data were presented as CFU ml-1.
Production of antifungal metabolites and effect of culture conditions
Bacillus sp strain IBA 33 was activated for 3 h in GNB media pancreatic peptone 22.5 g l-1, glucose 1 g l-1, K2HPO4 2 g l-1, NaCl 3 g l-1 pH 6.0, at 28 °C and 250 rpm.16
After activation, Bacillus sp strain IBA 33 was cultured on basal Landy medium (glucose 20 g l-1, Fe2 (SO4)3 H2O 0.15 mg l-1, MnSO4 H2O 5 mg l-1, CuSO4 H2O 0.16 mg l-1 K2HPO4 0.5 g l-1, KCl 0.25 g l-1 and MgSO4 0.25 g l-1 pH 6.0).16 In order to optimize the production of metabolites, the Landy medium (LM) was modified by adding different concentrations and sources of amino acids and carbohydrates at different culture conditions.
The aminoacids studied were glutamic acid (0.25%; 0.5% and 1%); aspartic acid (0.5%); glutamic acid and aspartic acid (0.5% respectively) and leucine (0.5%). The carbohydrates assayed were glucose and mannitol (1.5%; 2% and 2.5%).17
Culture conditions were pH 5.0; 6.0 and 7.0 at incubation temperatures of 20 °C, 28 °C and 37 °C.
250 ml of the resulting medium were cultured with 25 ml of active inoculum of Bacillus sp strain IBA 33 (108 cells ml-1), incubated at the studied temperature in a shaker at 250 rpm. Samples were taken as indicated in Figure 2. After centrifugation (10000 rpm, 15 min), the cell-free supernatants were concentrated to 57% by dialysis using a membrane of 100 °A and carboxymethylcellulose. The metabolites sterilization were carried out by: a) filtration using a cellulose nitrate filter pore: 0.22 μm and b) autoclaving at 121 °C for 15 min.
Times of isolation of Bacillus sp strain IBA 33 culture filtrates, A: after 12 h (logarithmic phase); B: after 24 h (end of logarithmic phase); C: after 48 h (stationary phase) and D: after 72 h (end phase).
Antifungal activity bioassays
In vitro assays, solid media
Petri dishes were filled with 10 ml of PDA and 0.1 ml of concentrated metabolites of each time period. After the plates were cooled, a defined piece of agar (0.7 cm of diameter and 0.2 cm of thickness) with grown mycelia was placed onto an agar surface. They were incubated for 5 days at 28 °C and the diameter of the mycelia growth was measured at the end of this period.
In vitro assays, liquid media
45 ml of potato dextrose broth pH 5.0; 2.5 ml spore suspension of each studied fungi (106 spores ml-1) and 2.5 ml of the concentrated metabolites (sterilized by filtration and autoclaving) were placed in 250 ml Erlenmeyer flasks. Flasks were incubated at 28 °C and 250 rpm for 5 days. Dry weight determinations were performed and the results expressed as percentage of inhibition growth.
Control assays were also made in 250 ml Erlenmeyer flasks that did not contain the concentrated metabolites.
In vivo assays
In the “in vivo assay,” % of green mold and sour rot in harvested lemons was performed to evaluate the fruit phytosanitary state. Lemons were harvested in July and August 2006 and received the same treatment they are given in packinghouses: they were washed with hot water (40 °C) for 5 min, submerged in a sodium bicarbonate solution at a concentration of 200 ppm for 2 min and sprayed with imidazole (Imazalil), 2,4 thiazalil benzimidazole (Thiabendazole) and wax. We worked with three batches of 36 lemons distributed in two boxes of 18 fruits per group. Group 1: lemons with the same treatment as the one received in packinghouses; Group 2: lemons treated with hot water, sodium bicarbonate and sprayed with metabolites of Bacillus sp strain IBA 33 and wax; Group 3: lemons without any treatment.
MBLM supplemented with 0.5% glutamic acid and corresponding to 72 h of culture was used.
The fruits were placed in boxes that contained moistened paper towels to maintain 85% humidity and were finally covered with plastic. They were kept for 3 weeks at 4 °C and 1 week at room temperature to reproduce the storage conditions to which the fruits are submitted until reaching their final destination. The results were expressed as % of diseased lemons.
Statistical Analysis
The data were analyzed by two-way analysis of variance (ANOVA) and the significant means were analyzed by Tukey's test and Kolmogorov-Smirnov test. Comparison of means was performed by Kruskal-Wallis's multiple range tests. Statistical significance was assessed at the level of p = 0.05.
Results
Identification of baterial strain
The biochemical profiling obtained were bacilli endospore forming, Gram (+), positive catalase activity, positive nitrate reduction, positive indole production, positive motility, growth at 7% NaCl, positive acid production, positive gelatinase and starch hidrolysis.
The 16S rRNA gene sequence determined for the strain consisted of 1028 nucleotides. On the basis of 16 S rRNA sequence analysis (Figs. 3 and 4), this strain appeared belong to the genus Bacillus (Group 1), and being closely related to B. cereus and B. thurigiensis.18
Sequence obtained from 16S RNA of Bacillus sp strain IBA 33 with the PLB primer.
Sequence obtained from 16S RNA of Bacillus sp strain IBA 33 with MLB primer.
In vitro assays
Growth inhibitions were higher in liquid than in solid media (data not shown), probably due to a smaller diffusion of the Bacillus sp strain IBA 33 metabolites in the solid culture media.
Its metabolites proved to be heat resistant because inhibition percentages remained the same after filtration and autoclaving.
Bacillus sp strain IBA 33 metabolite specificity towards fungi was found to depend on the composition of the LM. Because the LM contains glucose, which could exert some catabolic repression, we replaced that sugar with mannitol. However, the metabolites obtained with this last carbohydrate did not inhibit the growth of any of the fungi studied in the concentrations assayed. Only 2% glucose generated the results shown (all Figures).
Different incubation culture conditions (pH and temperature) showed the best results at pH 6.0 and 28 °C for the production of metabolites (all Figures).
According to references we assumed the peptide nature of metabolites produced by Bacillus sp.19 The effects of aspartic acid, that is structurally similar to glutamic acid and leucine, which in few cases is an activator of the protein synthesis,17 were studied upon the MBLM production.
The addition of leucine to LM did not show any effect on the synthesis of metabolites with antifungal activity (data not shown).
The major fungal growth inhibitions were obtained with metabolites of 72 h culture. Hence, we evaluated the antifungal activity of metabolites of 96 h, these metabolites did not produce growth inhibitions.
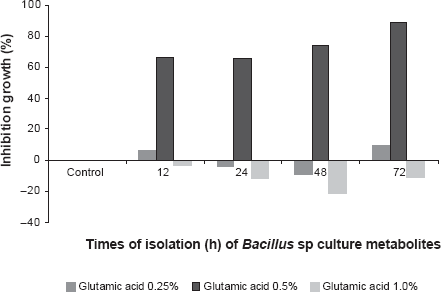
The most sensitive pathogenic fungus was G. candidum since MBLM with 0.5% glutamic acid of 12, 24, 48 and 72 h produced inhibitions of 65.58%, 66.55%, 72.61% and 88.44% respectively (Fig. 5). There is a highly significant difference in glutamic acid values (p < 0.01) according to variance analysis while Tukey's test (p < 0.05) detects the optimum value at 0.5%, but there is no statiscally significant time difference (p < 0.05). Aspartic acid and the mixture of glutamic acid and aspartic acid did not have any incidence on the synthesis of metabolites.
Percentage of inhibition growth of Geotrichum candidum in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM supplemented with glutamic acid (0.25%, 0.5% and 1.0%).
In the case of P. digitatum the greatest inhibition (65%) was observed when metabolites were recovered from LM (72 h) added with 0.5% of glutamic acid (Fig. 6). There is a statiscally significant difference (p < 0.01) between times according to variance analysis while Tukey's test detects the optimum (p < 0.05) at 72 h. Aspartic acid did not produce any effect and the same happened to the combination of glutamic acid and aspartic acid.
Percentage of inhibition growth of Penicillium digitatum in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM supplemented with glutamic acid (0.5%).
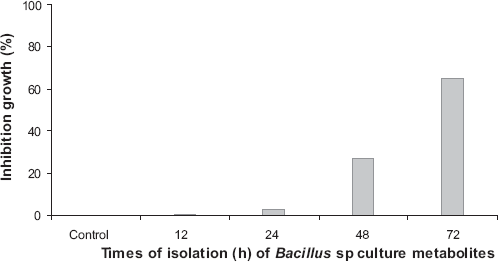
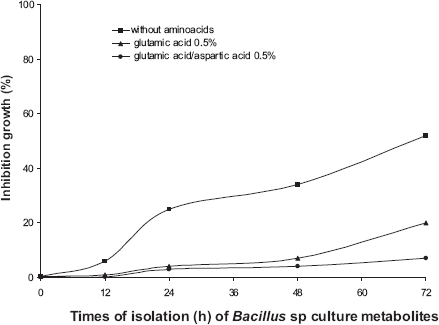
The growth of A. flavus was progressively inhibited by metabolites of Bacillus sp strain IBA 33 recovered from LM (without amino acids). At 72 h of culture, inhibition was 52%. Only 0.5% glutamic acid produced metabolites that inhibited A. flavus growth; metabolites from a 72 h culture produced an inhibition of 20%. Metabolites from LM supplemented with 0.5% glutamic acid and 0.5% aspartic acid (72 h culture) produced a 7% inhibition of A. flavus growth (Fig. 7). Glutamic acid (0 and 0.5%) has a significant difference (p = 0.01 <0.05) according to variance analysis. Glutamic acid and aspartic acid only have a significant difference between 12 and 72 h, reaching the optimum at 72 h, according to the Kruskal-Wallis range variance analysis.
Percentage of inhibition growth of Aspergillus flavus in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM without amino acids; LM supplemented with glutamic acid (0.5%), and glutamic acid/aspartic acid (0.5%).
Figure 8 shows that A. clavatus growth was inhibited by MBLM supplemented with 0.5% glutamic acid. The inhibition was evident (51%) with metabolites from a 24 h culture and was more intense with metabolites from a 72 h culture, where an inhibition of 87% was observed. There is a highly significant difference (p < 0.01) according to variance analysis.
Percentage of inhibition growth of Aspergillus clavatus in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM supplemented with glutamic acid (0.5%).
Metabolites from LM supplemented with 0.5% glutamic acid produced a 34% growth inhibition of A. niger (data not shown). When they were recovered from LM supplemented with 0.5% glutamic acid and 0.5% aspartic acid, the inhibition was 37% (Fig. 9). There is a significant difference only between 12 and 72 h, reaching the optimum at 72 h according to the Kruskal-Wallis range variance analysis. These results show that both amino acids are needed to partially inhibit A. niger growth.
Percentage of inhibition growth of Aspergillus niger in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM supplemented with glutamic acid/aspartic acid (0.5%).
In the case of P. expansum the inhibition was different according to the supplements incorporated to LM. When Bacillus sp strain IBA 33 grew in LM supplemented with 0.5% glutamic acid the highest inhibition was only 18%, whereas when it was cultivated with 0.5% aspartic acid the greatest inhibition was 45% (Fig. 10). There is a highly significant inhibition difference (p < 0.01) between glutamic acid (0.5%) and aspartic acid (0.5%) according to the Kolmogorov-Smirnov test. Statiscally there is a higher inhibition with aspartic acid (0.5%).
Percentage of inhibition growth of Penicillium expansum in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM supplemented with glutamic acid (0.5%) and aspartic acid (0.5%).
Growth inhibition of F. moniliforme was 92% when it was faced with metabolites produced in LM with 0.5% glutamic acid, but when LM was supplemented with glutamic acid and aspartic acid 0.5%, the major inhibition was only 73% (Fig. 11). There is a significant difference only between 12 and 72 h, reaching the optimum at 72 h according to the Kruskal-Wallis range variance analysis. There is a statiscally significant inhibition difference (p < 0.05) between glutamic acid and the combination glutamic and aspartic acids. Glutamic acid shows higher inhibition.
Percentage of inhibition growth of Fusarium moniliforme in presence of Bacillus sp strain IBA 33 culture metabolites recovered from LM supplemented with glutamic acid (0.5%) and glutamic/aspartic acid (0.5%).
In vivo assays
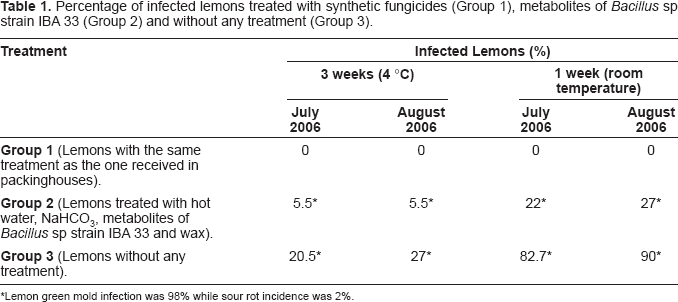
The evaluation of the phytosanitary state of lemons demonstrated that metabolites of Bacillus sp strain IBA 33 protected the fruit against green mold and sour rot (Table 1). Group 1 of lemons was healthy all through the month. Group 2, treated with metabolites, had 5.5% diseased fruit when it was retired from the cold chamber. After one week at room temperature the disease percentage increased to 22% and 27% (July and August, respectively). When the lemons did not receive any treatment (Group 3), disease incidence was between 20.5% and 27% in the cold chamber and 82.7%–90% after one week at room temperature. The disease incidence was greater in August than in July, probably due to high temperatures.
Percentage of infected lemons treated with synthetic fungicides (Group 1), metabolites of Bacillus sp strain IBA 33 (Group 2) and without any treatment (Group 3).
Treatment
Infected Lemons (%)
3 weeks (4 °C)
1 week (room temperature)
July 2006
August 2006
July 2006
August 2006
Group 1 (Lemons with the same treatment as the one received in packinghouses).
0
0
0
0
Group 2 (Lemons treated with hot water, NaHCO3, metabolites of Bacillus sp strain IBA 33 and wax).
Lemon green mold infection was 98% while sour rot incidence was 2%.
Discussion
The results of the present study show that Bacillus sp strain IBA 33 produces one or more metabolites with antifungal action against different phytopathogenic and non pathogenic fungi. The metabolites were heat-resistant because they conserved the same antifungal activity after sterilization at 121 °C for 10 min. That fact was reported in previews works.20
In this work it was demonstrated that Bacillus sp strain IBA 33 culture media composition and culture conditions (pH and temperature) are important factors to consider for the production of metabolites with antifungal action, especially with the addition of amino acids. The results are consistent with the ones obtained in previous studies, where the aggregate lipopeptides, exogenous to the culture media, generated a higher production of biosurfactant and therefore a greater antifungal activity. The highest growth inhibitions occurred in the presence of glucose (2%). Besides, there was an important biosurfactant production by B. subtilis DM-03 and DM-04.21
Many alternatives to chemical control have been investigated, but none, when used alone, provides the level of control of synthetic fungicides.
The metabolites of Bacillus sp srtain IBA 33 with a smaller antifungal activity could be combined with other methods to increase their effectiveness. The beneficial effect of treatments with hot water for Penicillium rot control was reported in citrus22,23 because it was demonstrated that heat inhibits spore germination and slows down P. digitatum growth. Recently the effect of hot water and bicarbonate with Bacillus subtilis was evaluated in order to control green mold and blue mold of citrus.24 A combination of the methods described above may be used to overcome the shortcomings of each.
In addition, in vivo tests demonstrated a good protective effect of the metabolites of Bacillus sp srtain IBA 33 against P. expansum, A. clavatus, P. digitatum and G. candidum. These results are important because there are bibliographical data that show that in vitro inhibitions can not be reproduced at in vivo assays.25
Gram positive bacteria, like Bacillus sp, however, have been studied less intensively than Gram-negative bacteria.26 The potential of Bacillus sp as a biological control agent to eliminate the use of synthetic fungicides has been previously reported,27 and it involves the production of cyclic lipopeptides including surfactin, iturin, fengycin and lichenycin.21Bacillus sp acts by producing an antibiotic of the iturin peptide family that affects membrane surface tension causing pore formation and resulting in the leakage of K+ and other vital ions causing cell death.28 It has been registered that a member of the family of the iturins, Bacillomicin F inhibits the growth of A. niger,29 while Iturin A inhibits the growth of A. flavus and F. moniliforme.30 Previous reports showed that Bacillus subtilis produced iturin in LM.31
This is the first study of Bacillus sp strain IBA 33 as a biological control agent against several species of fungi. This is important considering that Bacillus sp strain IBA 33 belongs to the same ecological niche of the studied fungi. Some of them are pathogenic and are under study from the point of view of biological control, while the saprophytic (species of Mucor, Fusarium, Aspergillus and others) are often resistant to the treatments applied.
Fungi presence has no effect on fruit quality and is not an indication of imminent decay. It is rather an indication of handling and storage conditions. The cotton- like material occasionally found on the stem-end of stored citrus fruits results from growth of saprophytic non-pathogenic fungi on senescing buttons (caliz and disk). High humidity conditions that favor growth of saprophytic fungi are actually very good conditions for maintaining fruit quality.32
Considering the above facts, the use of Gram-positive bacteria that have a natural formulation to suppress these two diseases would immensely help in overcoming the problems produced by the use of synthetic fungicides.
At present we are carrying out studies to determine the chemical nature and mode of action of Bacillus sp strain IBA 33 metabolites. The knowledge of the antagonism mechanisms in biocontrol may improve their effectiveness.
Footnotes
Abbreviations
The authors report no conflicts of interest.
References
1.
Vero MendezS., MondinoP.Control biológico poscosecha en Uruguay Horticultura Internacional.1999; 7: 26.
2.
WilsonC.L., GhaouthA., ChalutzE.Potential of induced Resistance to Control postharvest diseases of fruits and vegetables. Plant Dis.1994; 78: 837–844.
VeroS., MondinoP., BurgueñoJ., SoubesM., WisniewskiM.Caracterization of biocontrol activity of two yeast strains from Uruguay against ble mold of apple. Postharvest and Technology.2002; 23: 191–98.
5.
TeixidóN., UsallJ., PalauL., AsensioA., NunesC., ViñasI.Improving control of green and blue molds of oranges by combining Pantoea agglomerans (CPA-2) and sodium bicarbonate. Eur J Plant Pathol.2001; 107: 685–694.
6.
WilsonC.L., WisniewskiM.E.Biological control of postharvest diseases of citrus and vegetables an emerging technology. Ann Phytopathol.1989; 27: 425–441.
7.
WilsonC., WisniewskiM.F.Biological control of postharvest diseases of Fruits and Vegetables-Theory and Practice.CRC PressBoca Raton. FL.1994.
8.
FogliataG.M., TorresL.G.J., PloperL.D.Detection of imazalil-resistant strains of Penicillium digitatum sacc. In citrus packing-houses of Tucumán Province (Argentina) and their behaviour against current employed and alternative fungides. Rev Ind Agric Tucumán.2001; 77: 71–75.
9.
LingkW.Health risk evaluation of pesticide contaminations in drinking water. Gesunde Pflangen.1991; 43: 21–25.
10.
RasgaleN.N., SislerH.D.Social and Political implications of managing plant diseases with decreased availability of fungicides in the United States. Ann Rev Phytopathol.1994; 32: 45–557.
11.
Fernandez Larrea VegaO.Microorganismos antagonistas para el control fitosanitario. Manejo Integrado de Plagas (Costa Rica).2001; 62: 96–100.
12.
BergeyD.H., HoltJ.G.Bergey's manual of determinative bacteriology.Lippincott Williams and Wilkins, Baltimore, MD, U.S.A.1994.
13.
Popiech and Neumann A.1995.
14.
ZhangZ., SchwartzS., WagnerL., MillerW.A Greedy algorithm for aligning DNA secuences. J Comput Biol.2000; 200: 7(1–2): 203–214.
15.
MaldonadoM.C., RuncoR.S., NavarroA.R.Isolation, identification and antifungal susceptibility of lemon pathogenic and non pathogenic fungi. Rev Iberoam Micol.2005; 22: 57–59.
16.
DolejS., BochowH.Studies of the mode of action of Bacillus subtilis culture filtrates in the model pathosystem tomato seedling-Fusarium oxysporum. F. SP: Radicis-Lycopersici. Med Fac Landbouww Univ Gent.1996; 61/2b.
17.
RuncoR., NavarroA., MaldonadoM.C.Regulation of the production of poligalacturonase by Aspergillus terreus. World J Microbial and Biothecnol.2001; 17: 487–491.
18.
AshC., FarrowJ.A.E., WallbanksS., CollinsM.D.Phylogenetic heterogenity of the genus Bacillus revealed by comparative analysis of small-subunit-ribosomal RNA sequence. Letters of Applied Microbiology.1991; 13: 202–206.
19.
BechardJ., EastwellK.C., SholbergP.L., MazzaG., SkuraB.Isolation and partial chemical characterization of an antimicrobial peptide produced by a strain of Bacillus subtilis.J Agric Food Chem.1998; 46: 5355–5361.
MukherjeeA.K., DasK.Correlation between diverse cyclic lipopeptides production and regulation of growth and substrate utilization by Bacillus subtilis strains in a particular habitat. FEMS Microbiology Ecology.2005; 54: 479–489.
22.
PoratR., DausA., CohenL., FallikE., DrobyS.Reduction of postharvest decay in organic citrus fruit by a short hot water brushing treatment. postharvest Biol Technol.2000; 18: 151–157.
23.
AuretE.A.Control strategies for citrus postharvest diseases. Msc thesis, University of Pretoria, Pretoria, South Africa. 2000.
24.
SinghV., DaverallS.J.Bacillus subtilis as a control agent against fungal pathogens of citrus fruit. Trans Br Mycol Soc.1984; 83: 487–490.
25.
AgarryO.O., OshoB.I.In-vitro and In-vivo Inhibition of Aspergillus fumigatus by Pseudomonas fluorescens used as a microbial antagonist. Pakistan Journal of Nutrition.2005; 4: 371–37.
26.
San-LangW., ShihI.L., WangC.H., TsengK.C., ChangW.T.Production of antifungal compounds from chitin by Bacillus subtilis.Enzyme and Microbial Technology.2002; 31: 321–328.
27.
ObagwuJ., KorstenL.Intregrated control of citrus green and blue molds using Bacillus subtilis in combination with sodium bicarbonate or hot water. Postharvest Biology and Technology.2003; 28: 187–194.
28.
ThimonL., PeypouxF., Maget-DanaR., MichelG.Surfactive properties of antifungal lipopeptides produced by Bacillus subtilis.J Am Oil Chem Soc.1992; 69: 92–93.
29.
De LuccaA., WalshT.Antifungal peptides: Novel Therapeutic Compounds against emerging pathogens. Antimicrob Agents Chemother.1999; 1–11.
30.
KlichM.A., LaxA.R., BandJ.M.Inhibition of some mycotoxigenic fungi by Iturin A, a peptidolipid produced by Bacillus subtilis.Mycopathol.1991; 116: 77–80.
31.
AkpaE., JacquesP., WatheletB.Influence of culture conditions on lipopeptide production by Bacillus subtilis. Applied Biochemistry and Biotechnology.2001; 91–93.
32.
WhitesideJ.O., GarnseyS.M., TimmierL.W.Compendium of citrus diseases.The American Phytopathological Society Press, St. Paul, MN-80 pp. 1988.
33.
LoganN.A., BerkeleyR.C.W.Identification of Bacillus strains using the API system. J Gen Microbiol.1984; 130: 1871–1882.