
Abstract
Obesity has been linked to more aggressive characteristics of several cancers, including breast and prostate cancer. Adipose tissue appears to contribute to paracrine interactions in the tumor microenvironment. In particular, cancer-associated adipocytes interact reciprocally with cancer cells and influence cancer progression. Adipokines secreted from adipocytes likely form a key component of the paracrine signaling in the tumor microenvironment. In vitro coculture models allow for the assessment of specific adipokines in this interaction. Furthermore, micronutrients and macronutrients present in the diet may alter the secretion of adipokines from adipocytes. The effect of dietary fat and specific fatty acids on cancer progression in several in vivo model systems and cancer types is reviewed. The more common approaches of caloric restriction or diet-induced obesity in animal models establish that such dietary changes modulate tumor biology. This review seeks to explore available evidence regarding how diet may modulate tumor characteristics through changes in the role of adipocytes in the tumor microenvironment.
Introduction
The prevalence of obesity is a significant health challenge in most of the developed world. Evidence increasingly suggests that obesity mediates a more aggressive phenotype in certain cancers via a paracrine effect. The increased prevalence of both obesity and cancer highlights the importance of elucidating the basic mechanisms through which obesity may impact cancer aggressiveness.1,2
Adipocytes are recognized to form part of the tumor microenvironment in a variety of cancers.3–7 In both breast and prostate cancers, obesity is associated with higher grade disease.8,9 Other tumor sites in which obesity is linked to poorer outcomes include the kidney, pancreas, esophagus, thyroid, and endometrium.10–12 Notably, all of these sites have appreciable amounts of surrounding adipose tissue or stromal adipocytes.1,2,9,13–15 In in the context of hepatocellular carcinoma, lipid stores in the liver increase under obese conditions, contributing to alterations in local tumor microenvironment. 16 Both the quantity and quality of the surrounding adipose tissue may influence the biological and secretory behavior of adipocytes.17,18 This review article will explore how diet influences the relationship between adipocytes and cancer cells in the tumor microenvironment.
Tumor Microenvironment
In addition to adipocytes, other components of the tumor microenvironment include tumor-associated macrophages, fibroblasts (ie, cancer-associated fibroblasts), and endothelial cells. The cells of the tumor microenvironment are heterogenous and appear to differ from similar noncancer associated cells. 1 The cross talk between the stromal and the tumor cells is bidirectional, and a further understanding of this signaling may yield future therapeutic targets. While other cells of the tumor microenvironment mediate paracrine effects on tumor cells, 19 the unique aspect of adipocytes is their modifiable relationship with dietary intake. Therefore, diet (and exercise) may represent a potentially inexpensive and modifiable means to favorably alter the tumor microenvironment and subsequent tumor behavior. A schematic of the role of adipocytes and the influence of dietary factors is presented in Figure 1.
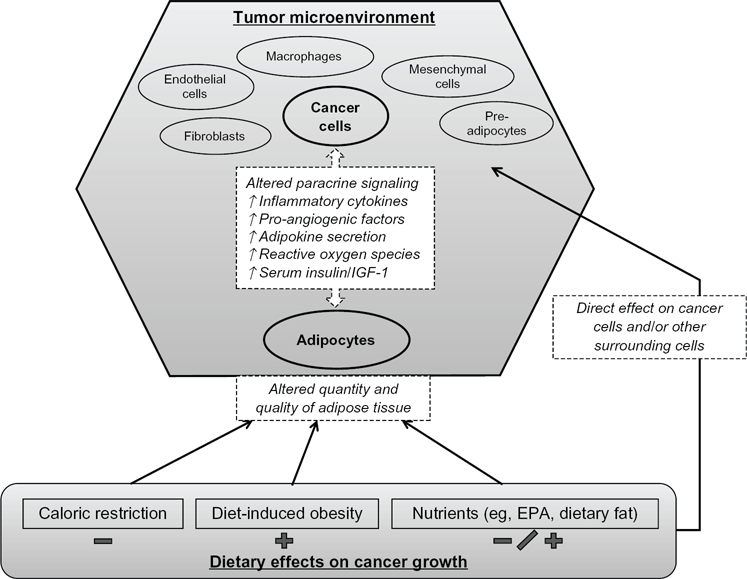
Dietary effects on adipocytes and cancer cells within the tumour microenvironment.
Several current therapeutic strategies currently target the tumor microenvironment. These include inhibitors of angiogenesis such as tyrosine kinase inhibitors and monoclonal antibodies against vascular endothelial growth factor (VEGF). In prostate cancer, denosumab inhibits the Receptor activator of nuclear factor kappa-B ligand (RANK-L) activation of osteoclasts in the bone microenvironment. The renewed interest in immunotherapy also represents a treatment that targets the tumor microenvironment instead of directly impacting the tumor.20–22
Cancer-Associated Adipocytes
A key question that remains unanswered at the cellular level relates to how adipocytes or obesity leads to the development of more aggressive cancer. Obesity represents increased lipid storage in white adipose tissue (WAT). White adipose tissue is comprised of adipocytes (forming the adipose fraction) as well as numerous other cell types of the stroma vascular fraction (SVF). 23 The smaller and more heterogenous SVF contains pre-adipocytes, mesenchymal stem cells, macrophages, endothelial cells, and fibroblasts.
The term cancer-associated adipocytes has been used to describe the abnormal adipocytes found adjacent to cancer cells.24,25 These adipocytes demonstrate decreased lipid content, a decrease in more differentiated adipose markers (eg, HSL, APN, resistin) and overexpression of several inflammatory cytokines (eg, IL6, IL-1β). 25 This peritumoral dysmorphism is not unique to adipocytes, with cancer-associated fibroblasts also showing an altered phenotype.26,27 These peritumoral cells form reciprocal signaling pathways with the tumor cells. The functional role of cancer-associated adipocytes in cancer initiation, progression, and resistance continues to be elucidated.
Evidence is now accumulating that adipocytes in the tumor microenvironment appear to promote a more aggressive cancer phenotype in some cancers.16,24,28 One recent study suggests endogenous WAT may serve as a reservoir of adipocytes and progenitors that populated the tumor microenvironment. 29 A more local origin is suggested by histologic evidence indicating a predominance of adipocytes at the invasive front of breast cancers. 30 Notably, the presence of invasive adipocytes at the margin of breast tumor growth indicates more aggressive disease. 31 Similarly, in a study utilizing prostate cancer specimens, infiltration of adipocytes was highly correlated with grade of cancer, with ≫90% of high grade specimens demonstrating infiltrative fat versus <20% in low grade cancers. 32 Thus, it appears that adipocytes form a critical role in the tumor microenvironment, particularly among aggressive tumors.33,34
Cancer-associated adipocytes have been shown to play a role in radioresistance to breast cancer cells 17 and are also implicated in chemoresistance. 34 Furthermore, these cancer-associated adipocytes appear to have altered gene expression and production of matrix metalloproteinases (MMPs), which may impact cancer invasiveness.35,36 Evidence also suggests that cancer-associated adipocytes may differentiate into cancer-associated fibroblasts. 37 Finally, cancer-associated adipocytes may also act as an energy source, coupling with the tumor cells. 38
Though it remains unclear whether cancer-associated adipocytes in humans are affected by diet in the same way as other adipocytes, in vitro studies suggest that they still maintain the basic functions of adipocytes such as lipid storage. 24 It is possible that through interactions with the cancer cells, their responsiveness to diet is significantly reduced. However, many of the experiments mentioned below do not support that view, assuming that adipocytes do indeed play a key functional role in the tumor microenvironment. Alternatively, diet-induced factors may alter the propensity of adipocytes to migrate into the tumor microenvironment to become cancer-associated adipocytes. Thus, obesity and diet may alter the relative supply of adipocytes that can migrate to the tumor microenvironment and there promote a more aggressive cancer.
Does Diet Influence Adipokine Secretion?
There is particular interest in the role adipokines may play in mediating the effects of obesity on cancer progression. Adipokines are a large heterogeneous group of molecules secreted by adipocytes. Adipokines affect many cellular functions including inflammation, cell growth, apoptosis, and motility. While it is well recognized that some epithelial cancers (eg, squamous cell carcinoma) can be caused by chronic inflammation and irritation, it is less clear if obesity-related inflammation plays a similar role in carcinogenesis, progression, and metastasis. Not all adipokines are implicated in cancer, and some studies describe the preventative role of particular adipokines.39,40 For instance, adiponectin is an anti-inflammatory adipokines of significant research interest in this regard.
Studies point to the fact that secreted factors released by adipocytes are critical in the regulation of tumor growth and spread. 5 For example, Park et al found that endotropins secreted from adipocytes increased angiogenesis, fibrosis, and inflammation in the mammary microenvironment through recruitment of macrophages and endothelial cells. 41 This was demonstrated to be mediated in part through NF-KB. The expression of endotropin was associated with both higher growth and increased metastases in breast cancer. Some of the other adipokines that have been studied in relation to cancer include, resistin, visfatin, β-NGF (nerve growth factor), HGF (hepatocyte growth factor), TNF-α, CCL2 (Monocyte chemotactic protein 1), IL-6, and osteopontin.
Recent research has demonstrated that diet can induce changes in the release of adipokines and in turn influence adjacent tissue function.18,42 Furthermore, dietary interventions can decrease serum IL-6 levels in humans 43 as well as alter serum levels of several other adipokines. 44 The secretion of adipokines is understood to change with changes in adipocyte size; 24 adipocyte size is in turn modulated by caloric intake. Notably, there is significant variability in the measurement of adipokines in human serum, which makes establishing a consistent correlation between diet and serum adipokines difficult. 45 Further research is still needed to determine if dietary changes may influence paracrine-adipokine signaling within the tumor microenvironment. Adipoparacrinology is a developing field of study that deals with the effects of adipokine secretion on adjacent tissue. 46
In Vitro Models
Modeling and testing the effect of adipokines on cancer progression in vitro has several challenges. Adipocyte hypertrophy and lipid droplet accumulation, which occur as a result of increased energy intake, are not reproducible in cell culture models. 24 In 3T3-L1 cell lines differentiated into adipocytes, multiple lipid droplets of triglyceride storages form instead of the one large droplet seen in vivo. 24 This altered phenotype may modify adipokine secretion, as seen with leptin in one study. 24 However, lipid droplet size is likely only one of many adipocyte and environmental factors regulating adipokine secretion. Therefore, simplified coculture models may help elucidate some of the reciprocal mechanisms between adipocytes and cancer cells.
Coculture models allow for isolated analysis of the effect of secreted molecules by adipocytes on cancer cells and vice versa. To date, most studies use the murine 3T3-L1 cells, which are differentiated into adipocytes. These cells appear to demonstrate results comparable to human tissue correlates. 25 For example, MCF-7 breast cancer cells have increased viability when grown in coculture with 3T3-L1 differentiated adipocytes. 44 Interestingly, cancer cell growth increased when the adipocytes were grown in higher glucose conditions; this effect appeared to be related to higher secretion of IL-8 and IGF-1. 44 Moreover, adipocyte-derived conditioned media (CM) from obese individuals resulted in increased MCF-7 cell proliferation, as well as higher IGF-1 production compared with adipocyte-derived CM from nonobese individuals, 44 suggesting that adipocytes in obese individuals have altered secretion of adipokines that promote cancer growth. Moreover, cancer cell growth was further regulated through culturing adipocytes in different glucose and fatty acid environments. 18 Taken together, these experiments validate the utility of coculture models to explore adipocyte-cancer cell interactions and also suggest that specific nutrient environments may alter these reciprocal interactions.
Dietary Fat
Dietary fat comprises a large part of the Westernized diet, which results in increased adipose tissue via adipocyte hypertrophy and hyperplasia. Similar to quantitative alterations, dietary fat influences the qualitative function of adipocytes. Dietary fatty (unsaturated vs. saturated) acids may influence adipokine release through their influence on phospholipid membrane fluidity, transcription factors, or other unknown factors. 47 This represents one of the methods by which dietary fat may influence cancer progression. Other research has suggested that a high-fat diet may also be involved in cancer initiation. 48
Several animal studies have evaluated diets containing different fat content on prostate cancer progression. Animal models are commonly used to study the effects of dietary interventions on cancer progression. Genetically engineered mouse models are particularly useful due to a lack of immunosuppression and the spontaneous, orthotopic development of tumors. In the transgenic adenocarcinoma of the mouse prostate (TRAMP) model, 0/17 mice fed a low-fat diet (8% dietary fat) developed tumors by 18 weeks compared with 7/20 in the high-fat group (20% dietary fat). There were a similar number of tumors found at 24 weeks, suggesting that a low-fat diet delays cancer progression in the TRAMP model. 49 Another study utilizing the TRAMP mouse found that when increasing epididymal fat (a marker of visceral fat) by the implementation of a Westernized high-fat diet, a subsequent increase in the number of more aggressive tumors resulted. 50 Similar results have also been observed in xenograft models.51,52
There is some evidence that specific fatty acids may impact adipokine secretion. While mechanisms are mostly unknown, it is likely that secretion of adipokines is regulated in part through a complex network involving dietary fatty acids. Oleic acid, a monounsaturated fatty acid, is one of the most abundant fatty acids in adipose tissue. Its tissue levels correlate with dietary intake 53 and appear to alter adipokine secretion, which may promote cancer progression.54,55 Palmitic acid is another fatty acid implicated in altered metabolic conditions that occur with increased adiposity. For example, in mice fed high-fat diets of either soybean oil or palmitic acid, IL-6 levels in adipose tissue were three-fold higher in those mice fed the palmitic acid. 56 This suggests that some cancer-promoting cytokines may increase with higher levels of fatty acids seen in obesity. Conversely, eicosapentaenoic acid (EPA) appears to lower inflammatory cytokine secretion from adipocytes. Perez-Matute et al 57 demonstrated in mice that EPA appears to block the increases in TNF-α induced by a high-fat diet. The omega-3 fatty acid EPA has also been shown in mice to increase levels of the anti-inflammatory adipokine, adiponectin. 58
Caloric Restriction
Dietary caloric restriction appears to have an effect on cancer progression. 59 Whether this is via direct effects on adipocytes or due to other systemic factors remains unclear. Some studies have found that caloric restriction can exert an anticancer effect through alterations in systemic IGF-1 and NF-κB levels.60,61 In the TRAMP model for prostate cancer, 62 intermittent calorie restriction was observed to increase survival time and delay prostate cancer detection. 63 Other in vivo models have demonstrated caloric restriction induced decreases in adipose tissue levels of IL-6, TNF-α and adiponectin. 18 However, these diet-induced changes varied between the adipose depot locations in the mice, suggesting that a tissue-specific response to diet alterations exists. 18 Alternatively, some studies now suggest that caloric restriction may also regulate tumor cells directly through modulation of the PI3K/Akt/mTOR pathway.64,65
Diet-Induced Obesity
There are many studies implicating diet-induced obesity (DIO) to correlate with larger tumors in murine xenografts models, although some study results conflict (Table 1). The exact mechanism of action of the observed effects of DIO on cancer is unclear, though locally secreted factors implicated include, leptin, VEGF, TNF-α, and IL-6. 66 Increased tumor cell invasiveness may represent another surrogate of altered modulation of the tumor microenvironment. Blando et al, showed that DIO increased the incidence of aggressive adenocarcinoma with stromal invasion at 6 months. 67 Increased levels of NFκB in the ventral prostate suggest inflammatory changes in the tumor environment occur as a result of DIO. Additionally, the mRNA levels for IL-1α, IL-1β, IL-6, IL-7, IL-23, IL-27, NFκB1, TNFα, and VEGF family members were also significantly increased with DIO. Further, increased vascularity and significant infiltration of the tumor microenvironment with adipocytes and inflammatory cells was noted in DIO mice. 67 These results suggest that the level of pro-inflammatory and pro-angiogenic factors are increased via dietary changes, though the cell of origin is not definitely implied. 67
Reported impact of diet-induced obesity on murine tumor growth.

Excess dietary intake induces a positive energy balance and may also facilitate adipocytes to induce metabolic stress in coordination with immune cells. 68 This metabolic stress, including excess production of reactive oxygen species, hyperinsulinemia, and elevated adipokine secretion may favor increased tumor growth. In particular, the insulin pathway appears to be essential for tumor growth in DIO mouse models of colon cancer. 69 Similar to prostate cancer, inflammatory cytokines were also increased in DIO colon cancer models.70,71 Taken together, it appears that DIO results in changes in the tumor microenvironment that increase inflammation and hyperinsulinemia and promote tumor growth and aggressiveness.
Other studies also further support the view that it is the positive energy balance independent of diet that results in these phenotypic changes in cancer. Zhang et al 29 prefed mice with low- or high-fat diets and then switched them over to a normal chow for 1 month before implantation of tumor cells. They found increased tumor growth in obese mice compared with lean mice, suggesting the effect may be due to the presence of white adipose tissue. Further, the tumor growth rate was not significantly different in DIO-resistant mice versus lean mice, suggesting that overfeeding alone does not induce greater tumor growth. 29 Finally, their experiments suggested that endogenous adipocyte stem cells may populate the tumor microenvironment and exert a paracrine effect to induce tumor growth. Similarly, results from TRAMP mice fed an ad libitum diet revealed that the temperature at which the mice were housed influenced prostate cancer progression. 72 Overall, it appears that diet-induced obesity impacts tumor aggressiveness via indirect effects through metabolic and inflammatory changes associated with excess adipose tissue.
Summary
Elucidating the role of diet in cancer progression is challenging, given the complexity and variability of biological systems. Dietary changes cannot easily be modeled in vitro. Moreover, animal models do not respond uniformly to dietary changes. Overall, a review of the current literature is indicative that the effects of diet on the host metabolism may be more important than any direct effect imparted on cancer cells. Subsequently, it is easier to correlate the outcome of interest (eg, cancer progression) with the effects of diet (eg, obesity) than to the diet itself. Nonetheless, in addition to modifications in caloric intake, there are suggestions that specific dietary fats may alter the function of adipocytes and subsequently influence the tumor microenvironment. Thus, the importance of adipocytes and their changes with diet continues to be a fascinating area relevant to cancer progression.
Footnotes
Author Contributions
Conceived and designed the concept: PT, VV. Analyzed the data: PT, BCM, VV. Wrote the first draft of the manuscript: PT, BCM. Contributed to the writing of the manuscript: PT, BCM, VV. Agree with manuscript results and conclusions: PT, BCM, VV. Made critical revisions and approved final version: PT, BCM, VV. All authors reviewed and approved of the final manuscript.
Funding
Author(s) disclose no funding sources.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.