
Abstract
MicroRNAs (miRNAs) are a class of small regulatory RNAs involved in gene regulation. The regulation is effected by either translational inhibition or transcriptional silencing. In vertebrates, the importance of miRNA in development was discovered from mice and zebrafish dicer knockouts. The miRNA-9 (miR-9) is one of the most highly expressed miRNAs in the early and adult vertebrate brain. It has diverse functions within the developing vertebrate brain. In this article, the role of miR-9 in the developing forebrain (telencephalon and diencephalon), midbrain, hindbrain, and spinal cord of vertebrate species is highlighted. In the forebrain, miR-9 is necessary for the proper development of dorsoventral telencephalon by targeting marker genes expressed in the telencephalon. It regulates proliferation in telencephalon by regulating Foxg1, Pax6, Gsh2, and Meis2 genes. The feedback loop regulation between miR-9 and Nr2e1/Tlx helps in neuronal migration and differentiation. Targeting Foxp1 and Foxp2, and Map1b by miR-9 regulates the radial migration of neurons and axonal development. In the organizers, miR-9 is inversely regulated by hairy1 and Fgf8 to maintain zona limitans interthalamica and midbrain-hindbrain boundary (MHB). It maintains the MHB by inhibiting Fgf signaling genes and is involved in the neurogenesis of the midbrain-hindbrain by regulating Her genes. In the hindbrain, miR-9 modulates progenitor proliferation and differentiation by regulating Her genes and Elav3. In the spinal cord, miR-9 modulates the regulation of Foxp1 and Onecut1 for motor neuron development. In the forebrain, midbrain, and hindbrain, miR-9 is necessary for proper neuronal progenitor maintenance, neurogenesis, and differentiation. In vertebrate brain development, miR-9 is involved in regulating several region-specific genes in a spatiotemporal pattern.
Introduction
MicroRNAs (miRNAs) are endogenously expressed 20–25 nucleotide-long RNA molecules whose identification began with lin-4 in Caenorhabditis elegans, where it was shown to be involved in gene regulation at the larval developmental stage. 1 Since then, miRNAs have been reported to regulate multiple biological processes from development to cell proliferation, growth regulation, and apoptosis. 2
MiRNAs are transcribed by RNA Pol II as primary transcripts (primary miRNA, pri-miRNAs), which are usually several kilobases long and contain local stem-loop structures. 3 The maturation of the miRNA begins with the cleavage of larger hairpins into smaller hairpins called the precursor miRNA (pre-miRNA) by Drosha. 4 In the case of Drosophila, an additional protein Pasha was identified and was shown to have pri-miRNA-processing capability. Pasha knockouts (KOs) showed the accumulation of pri-miRNA suggesting that Drosha works together with Pasha. 5 Post nuclear processing, the pre-miRNAs are transported into the cytoplasm where they are processed again by the enzyme dicer, generating the mature miRNA. 6 The miRNA duplexes are incorporated into a protein complex consisting of Argonaute and accessory proteins called the pre-miRNA-induced silencing complex (pre-RISC). 7 The removal of the passenger strand from the miRNA duplex results in the formation of the mature RISC complex, which then targets suitable m RNAs resulting in posttranscriptional repression or the destruction of the mRNA target (Fig. 1). 8 10

miRNA biogenesis.
miRNAs in Development
The miRNAs are involved in numerous cellular and biological events. 11 13 They are ideal candidates for the fine-tuning of gene expression due to their nature of having specific spatiotemporal expression patterns. 14 The importance of miRNAs in development is revealed by the loss of miRNA-processing enzymes and its machinery proteins (dicer, Drosha, dgcr8, Argonaute). 15 19
Dicer significance in development
Dicer is essential for the processing of mammalian miRNA. Dicer knockout (KO) studies in mice and zebrafish16,17 have provided insights into the role of miRNAs from a developmental perspective. Dicer ablation results in embryonic lethality in mice 16 and zebrafish; 17 thus, conditional KO (CKO) serves as a tool to study the role of dicer in the vertebrate system.
Conditional dicer KO in development
The functional implication for the necessity of miRNA in development comes from the zebrafish maternal-zygotic (MZy) dicer KO embryo, a CKO. The MZy dicer shows abnormalities in brain development with the reduction in the formation of brain vesicles. 20 In mice, the dicer CKOs were generated using cre-lox mechanism. The normal expression patterns of the brain-specific genes in which the CKOs were created are as follows: Foxg1-cre 21 at E8.0, Wnt1-cre 22 at E8.5, Emx1-cre at E9.5,23–26 Nestin-cre at E10.5, 27 Nex-cre28,29 and hGFAP-cre30,31 at E13.5, and CaMKII-α-cre at E15.5 32 (Table 1, Fig. 2A and B).

Time line of gene expression and gene expression pattern in the developing brain.
Summary of dicer Knockout studies.

The dicer CKOs from Foxg1-cre, Emx1-cre, and Nestin-cre show that miRNAs are necessary in the early forebrain development and Nex-cre, CAMKII-cre, and GFAP-cre in the postnatal forebrain development.28,33–36 The Wnt1-cre dicer demonstrates the necessity of the miRNAs in midbrain and hindbrain development. 37 Finally, Wnt1-cre and hGFAP-cre dicer shows the necessity of miRNAs in hindbrain development.30,37 Some of the CKO embryos died early or postnatal.28,35,38 The dicer CKOs provide valuable information on the role of miRNAs in development which include cell proliferation, neurogenesis, migration, maturation, and differentiation. The dicer KO studies are described in detail in the following sections in context with the role of miRNA-9 (miR-9) in brain development.
Role of miR-9 in development
The miR-9 is one of the highly expressed miRNAs in the developing vertebrate brain and involved in several cellular functions and development. 39–41 The miR-9 is conserved among the vertebrates and specifically expressed in the central and peripheral nervous system; 39 42 it has also been reported to be present in multiple copies (Table 2).41,43 The retention of multiple copies has also been linked to the differential expression of its paralogs and specialization in their function. 44 The first evidence of its function in the nervous system comes from mouse, 45 where it has been reported to be involved in the patterning of brain structures and in cellular events like proliferation, maturation, and differentiation of neurons in the brain. 46 49 The expression pattern of miR-9 in the vertebrates is summarized in Table 3.
miR-9 and their location in chromosomes of different species.
MiR-9 expression in the developing brain of different species.

Apart from its prominent role in development, miR-9 has been correlated with several other cellular roles and pathology.2,50 The most significant of these roles is its association with cancer, where aberrant levels of the miRNA have been identified in tumor formation or progression. 51 In the case of gastric cancer, miR-9 knockdowns have shown an inhibitory role in the proliferation of tumor cells. 52 Contrastingly, in the case of ovarian cancer cells, the overexpression of miR-9 was shown to be useful in the suppression of tumor proliferation. 53 The miR-9 has also been shown to regulate interferon-regulated genes and MHC class I molecules. 54 In this review, the role of miR-9 in early brain development is summarized.
Brain Development
The developing central nervous system (CNS) is regionalized into four distinct regions, namely prosencephalon (forebrain), mesencephalon (midbrain), rhombencephalon (hindbrain), and spinal cord. The vertebrate forebrain is further divided anteriorly into telencephalon (prospective cerebral hemisphere) and posteriorly into diencephalon (prospective thalamus and hypothalamus). The rhombencephalon is subdivided into metencephalon (prospective pons and cerebellum) and myel-encephalon (prospective medulla oblongata).55,56 The organizers mediate the anterior-posterior (AP) axis and dorsoventral (DV) axis development. The organizers are special group of cells that produce secreted molecules and act as long-lasting coordinators of many cellular events like cell fate, survival, proliferation, maturation, and differentiation. The organizers along the AP axis are anterior neural ridge (ANR)/anterior neural border (ANB), zona limitans interthalamica (ZLI), and isthmus or midbrain-hindbrain boundary (MHB).57,58
Cell biology of neurogenesis
In the early phase of development, the neural plate and neural tube are made of a single layer of neuroepithelial cells. The neuroepithelial cells are primary neural stem cells (NSCs) in the developing neural tube. These NSCs divide symmetrically resulting in massive expansion prior to neurogenesis. 59 The telencephalic neurogenesis in mice takes place between E10 and E17.33,60 In chicken, the telencephalic neurogenesis takes place between E4 (HH24) and E10, 61 while the neurogenesis of the optic tectum takes place between E3 (HH18) and E20 (HH45).62,63 In zebrafish, the telencephalic formation and the morphogenetic movement take place between 18-hour postfertilization (hpf) and 5 day postfertilization (dpf), 64 and the tectal neurogenesis starts by 24 hpf. 65 Cortical neurogenesis takes place between stage 22 and 36,66,67 and tectal neurogenesis between stage 35 and 58 in Xenopus. 68 In the early phase, the NSCs divide asymmetrically and form the NSC and Cajal-Retzius (CR) neurons. The generation of CR neurons is modulated by miR-9.47,60 The CR neurons69,70 migrate from the ventricular zone (VZ) to the basal lamina that forms the preplate. As the development progresses, the preplate forms the subventricular zone (SVZ), intermediate zone (IZ), the cortical plate (CP), and the marginal/mantle zone (MZ). 71 In later stages, the NSCs give rise to the neuronal progenitor cell types: the apical (cells from the VZ) and basal progenitors (cells from the SVZ) and neurons (reside in the MZ). The apical and basal progenitors consist of radial glia and intermediate progenitors. 72 These progenitors mature and form the neurons in the MZ (Fig. 3).59,73,74
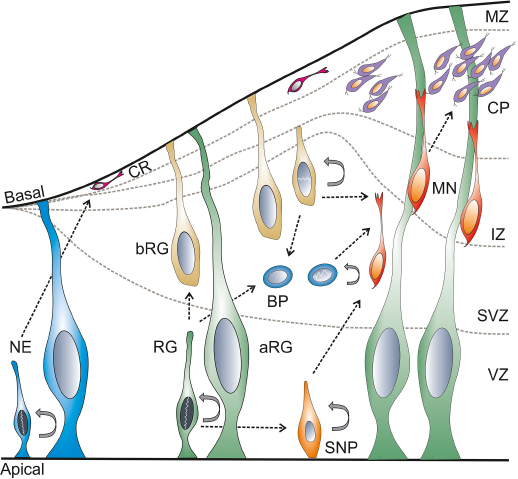
Cell biology of neurogenesis.
The miR-9 is expressed throughout the cellular development of neurons, ie, proliferation, neurogenesis, migration, maturation, and differentiation.45,74–78 The miR-9 is also one of the specific markers for neuronal cells. 79 It is expressed in the proliferating and differentiating cells in the telencephalon, diencephalon, and optic tectum. 39 The miR-9 regulates proliferation by targeting several transcription factors (Foxg1, Meis2, hairy1, hes1, her genes, etc.).46–49,80 The miR-9 suppresses TLX (human homolog of the tailless gene; also known as nuclear receptor subfamily 2, group E member 1 [Nr2e1]) expression to negatively regulate proliferation and accelerate neural differentiation. 75 Along with miR-124, miR-9 acts as a key factor in regulating neuronal differentiation. 77 Axon development in the developing cortex is regulated by miR-9 via targeting Map1b. 81 Neurite outgrowth/migration is also controlled by miR-9 through its regulation of Foxp1 and Foxp2.82,83
Telencephalon development
Telencephalon is derived from the rostral margin of the neural plate. 84 During early gastrulation, the formation of the neural plate (neural induction) is promoted by fibroblast growth factor (Fgf) and repressed by transforming growth factor-β and Wnt signaling. Following neural induction, the rostral margin neural plate cells known as ANR in mouse or the ANB in zebrafish which express Fgf8 is essential for the development of telencephalon.85–87
During the early telencephalon development, at the closing end of the neural tube, the telencephalic midline is formed. Within the midline, there are three signaling centers: the dorsal midline, the rostral midline, and the ventral midline. The dorsal midline expresses several bone morphogenetic protein (BMP) genes. 88 The ANR/ANB and the nearby area becomes the rostral midline that expresses several Fgf genes including Fgf8. 89 The ventral midline is defined by the expression of sonic hedgehog (SHH) signaling. 90
The DV patterning of telencephalon is the first step in the formation of the neocortex. The dorsal half forms the pallium, later develops into the cortex, and the ventral half forms the subpallium that later develops into the basal ganglia. 57 The pallium is divided into four main subregions: dorsal, medial, lateral, and ventral pallium. The DV patterning is defined by the generation of different cell types from the dorsal and ventral precursor cells; dorsally, they give rise to glutamatergic neurons and form the neocortex and hippocampus, and ventrally they give rise to GABAergic neurons and form the medial ganglionic eminence (MGE) and lateral ganglionic eminence (LGE), which form the striatum and globus pallidus. 91 The transcription factor, Pax6, is expressed in the dorsal telencephalon, whereas Gsh2 is expressed in the ventral telencephalon. The counteraction between Pax6 and Gsh2 forms the pallio-subpallial boundary (PSB; Fig. 4).92,93

Gene expression in the developing telencephalon.
Overall, the telencephalon is induced by the repression of Wnt signaling. The formation of the telencephalic midline involves the interaction of Shh ventrally, Fgf genes rostrally, Bmp laterally, and Wnt genes dorsally. The signaling centers regulate the expression of Emx2, Pax6, and Gsh2, which is necessary for the regulation of region-specific neurogenic genes such as Ngn1 and Mash1 on dorsal and ventral telencephalon, respectively (Fig. 4).91,94
Role of miR-9 in telencephalon development of mice
In mice, miR-9 has three variants such as miR-9-1, miR-9-2, and miR-9-3 that express the same mature product, miR-9. 95 The miR-9-2 is abundantly expressed in the telencephalon, while miR-9-1 and miR-9-3 are observed in the proliferative zone (Table 2). 80 The miR-9 expression is first observed at E9.0 stage in the telencephalic primordium. At E10.5, miR-9 is observed in optic vesicles and spinal cord, where they are dorsolaterally graded in the cortex with the subcortex sparsely expressing miR-9. The caudal pallium expresses miR-9 at E10.5; later at E11.5, in the medial pallium, and at E12.5, the expression was restricted to the medial pallium and retained in the MZ of the entire cortex including ventral pallium. 47 The hippocampus that is derived from the medial pallium expresses miR-9. 28 Histology shows that miR-9 is expressed throughout the proliferative zone of the developing telencephalon80,96 and in some differentiating neurons of the neocortex.49,96,97
The forkhead box protein G1 (Foxg1) is the earliest expression marker in the telencephalon and is expressed at E8.0. 98 The Foxg1-cre dicer KO shows that there are no obvious changes in the telencephalic markers: the pan-telencephalic marker such as FoxG1 and dorsal telencephalic markers such as Pax6, Emx2, and Ngn2, and ventral telencephalic markers such as Olig2 and Dlx2. The miR-9 was reduced and undetectable in the telencephalon at E11.5 stage, when neurogenesis occurs. The Nestin (a progenitor marker) in the VZ radial progenitor cells was reduced, with a loss of transcription factor, Sox9 (a co-enhancer of Nestin gene)99,100 and Erb2 receptor (mediator for the formation of radial glial cells via neuregulin 1 signaling). 101 The postmitotic cells were markedly reduced in E12.5 telencephalon. The CR cells have been depleted in this dicer KO. The proposition of basal progenitor Tbr2 cells has been increased in E11.5 ventral telencephalon and appears at the telencephalic wall, where these cells are not usually found. The telencephalon size was markedly reduced due to apoptosis from E12.5 onward. 33
Interestingly, Foxg1 is upregulated and miR-9 is downregulated in the olfactory and GnRH neurons in Dlx5 (distal-less homeobox gene) mutant mice. Thus, proving that miR-9 regulates Foxg1 and Dlx5 facilitates the production of miR-9. 102 Dlx5 is one of the earliest markers for the rostral ectoderm. The Dlx5 is expressed in the ANR at E7.5, ventral cephalic epithelium at E8.5 and from E9.5 onward in the telencephalon.103,104 However, in the telencephalon, the interaction between Dlx5 and miR-9 is yet to be established.
The Emx1 (empty spiracles homolog 1) is specifically expressed in the dorsal telencephalon and observed at E9.5.23–25The Emx1-cre dicer KO shows that the miR-9 is barely detectable from E10.5 telencephalon in the VZ neural progenitors cells. Detailed study indicates that the miR-9 expression is abrogated in the preplate (E11.5), CP (E12.5), and VZ of the telencephalon in comparison to the normal phenotype. 96 There is no difference in the apical and basal progenitor population, but there is a reduction in the cortical neuronal layer, which was attributed to apoptosis via depletion of miRNAs. Until E13.5, the apical (Pax6-expressing cells) and basal (Tbr2-expressing cells) progenitor cell division and progression were not affected; 96 however, at E14.5, there was a decrease in the mitotic apical and basal progenitors.36,96 At E15, the cortices had higher Tbr1 and Tuj1 expression indicating that early neurogenesis is altered. At E17.5, the cerebral cortices were smaller, and the VZ and IZ were reduced with slightly enlarged CP. The Emx1-cre dicer KO mice survived until postnatal day 30 (P30). 36
The Emx1-cre dicer KO also shows severe defects in the hippocampus with a reduction in miR-9 at E16.5. The Emx1-cre has reduced hippocampal neuroepithelial cells and dentate gyrus (DG) granular progenitors. 28 The early hippocampal marker, Lhx2, which helps in the expansion of the cortical hem 105 where CR neurons are formed, 106 was reduced in the hippocampus from E13.5 onward. At E16, the hippocampal marker such as Wnt7b and Lef1 had been reduced in the hippocampus and fimbria and Prox1 in the DG. 28 The CA1, CA3, and DG regions of hippocampus are formed at E18.5.107,108 The Tbr2 (progenitor marker) and Prox1 (neuronal marker) expression in the DG was absent. Ectopic expression of Tbr2 was observed in cortical progenitor zone of Emx1-cre dicer KOs. However, mature neurons were reduced in all the three regions. 28
The Emx1-cre dicer KO has increased Foxp2 expression in the dorsal telencephalon as well as in some neural progenitors of the VZ, 82 which was not observed in the wildtype. 109 Similarly in miR-9-2/3 double KO at E18.5, the Foxp2 was increased in the layer VI concluding that Foxp2 expression was increased in the absence of miR-9. 80 Foxp2 is a member of forkhead family of transcription factors. Foxp2 is expressed in the ganglionic eminence as early as E12.5, in the CP at E14.5, and from E16.5 onwards as a gradient from lateralhigh to mediallow cortex. They are expressed in the migratory or postmigratory neurons of the striatum. 109 The Foxp2 has a 3′ untranslated region (UTR) target site for miR-9. Experimental evidence proved that the increased Foxp2 in the dorsal telencephalon of Emx1-cre dicer is due to the lack of miR-9, 82 which is similar to miR-9–2/3 mutant. 80 Ectopic Foxp2 expression in the embryonic neocortex impairs radial migration of neurons. 82 In the dorsal telencephalon, miR-9 represses Foxp2 expression for proper dorsal telencephalon development that includes radial migration of neurons.
The Nestin expression starts at E9.5 in the developing cerebral cortex,110,111 The Nestin-cre dicer mice did not survive postnatally. At E15, the Tuj1 (a neurofilament marker) was found in IZ, early-born Tbr1 neurons in the CP, and the neurofilament was observed in the axons of IZ. This denotes early neurogenesis being unaffected. No difference in the expression of neuronal progenitor markers such as Pax6 and Tbr2 was observed. At E18, the brain size of the mice appears to be normal. However, histological sections show larger ventricles, reduced dividing cells in the VZ, SVZ and thinning of postmitotic neurons in the CP. This shows that the late-born neurons are affected, indicating the need for miRNA for proper function. 36
The Nestin-cre dicer KO has reduced hippocampal size, and miR-9 was undetectable in the cortex and hippocampus at E14.5. Similar to Emx1-cre, Nestin-cre also has reduced hippocampal neuroepithelial cells and DG granular progenitors. The early hippocampal marker Lhx2 was reduced from E18.5. The DG Tbr2 (progenitor marker) and Prox1 (neuronal marker) expression was reduced. Similar to Emx-cre dicer KO, Tbr2 was ectopically expressed. Early-born neurons expressing Tbr1 were increased in CA1 and DG regions, although mature neurons were reduced in all the three regions of the hippocampus. 28
McLoughlin et al 111 reported that Nestin-cre dicer mutant has cortical thinning and enlargement of lateral ventricles at E16. At E15.5, the nestin expression was distributed throughout the cortical parenchyma. The neural precursor cell proliferation was greatly reduced in the VZ and SVZ of E15.5 and E16 cortex due to delayed cell division. The thinning of cortex is also due to apoptosis. The CR neuron migration and differentiation was defective, which was observed as more CR neurons in the CP than on the MZ. The premature maturation of neurons was observed by the overproduction of DCX and Rnd2 (a maturation marker). Precocious astrocyte differentiation was also observed in the Nestin-cre dicer KO cortex. 111
The Nex expression starts at E13.5 in the hippocampus.28,29 The Nex-cre dicer KO showed smaller brains and died at P23. 38 The miR-9 was reduced at P22 in the hippocampus. The CA3 region is severely affected in Nex-cre dicer KO. The mature neurons were reduced in all the three regions. 28 The CaMKII-cre dicer KO resulted in microcephaly and reduction in the size of white matter tracts. 35 The CaMKII-cre was specific to forebrain structures such as the thalamus, cortex, and hippocampus with little emphasis on cerebellum, medulla, and pons.32,36 The hGFAP starts at E13.5. 31 The hGFAP-cre dicer KO shows that miR-9 is necessary for proper cerebral cortex development via Notch1 signaling. 34 However, there are no significant abnormalities during embryonic stages in these KOs (Nex-cre, CaMKII-cre, and hGFAP-cre). This might be due to the delayed dicer deletion. These studies indicate that dicer is necessary for the survival of postnatal neurons.
The conditional dicer KOs, namely Foxg1, Emx1, Nestin, Nex, GFAP, and CaMKII, show loss of miR-9 expression in the telencephalon.28,33,34,112 The conditional dicer KOs point out the influence of dicer in early and late neurogenesis in the developing telencephalon. However, it also provides indirect evidence for the necessity of miR-9 in telencephalic development; the role of miR-9 in the development of telencephalon comes from miR-9 KO study. The miR-9-2/3 double KO exhibits severe growth retardation with small cerebral hemispheres and olfactory bulbs, and they died within one week postnatal. Histology of postnatal miR-9–2/3 double KO showed reduction in cortical and VZ, with an expansion of lateral ventricles. The proliferating zone in subpallium is hyperplastic with reduced differentiation. The miR-9-2 or miR-9-3 mutant does not show any defect in neurogenesis in pallium and subpallium; however, the miR-9-2/3 double-mutant does. 80 This miR-9-2/3 KO shows that miR-9 is necessary for proper telencephalic development by controlling neural progenitor proliferation and differentiation. Overall, Foxg1-cre, Emx1-cre, Nestin-cre, Nex-cre, and hGFAP-cre dicer KOs point out that miR-9 is necessary for the early and late neurogenesis of the developing cortex and hippocampus.
The miR-9-2/3 double mutant shows the direct role of miR-9 in telencephalon development. The telencephalic markers such as Foxg1, Nr2e1/T1x, Pax6, Meis2 (myeloid ecotropic viral integration site 2), and Gsh2 have miR-9 target binding sites in their 3′UTR. 80 The miR-9 targets Foxg1 for the generation of CR neurons in the medial pallium. The miR-9 and Foxg1 genes are expressed in reciprocal gradient along the medial pallium at E11.5. 47 The Foxg1/BF1 is necessary for the early development of telencephalon.87,113 Foxg1 mutants displayed delayed cell cycle progression, reduction in proliferative progenitors and increased neurogenesis. Foxg1 controls proliferation through the regulation of Fgf signaling and differentiation through the regulation of BMP signaling. 114 The Foxg1-driven dicer mutant also shows that miR-9 is depleted and undetected at E11.5 in the cortex. 33 In the early stages of miR-9-2/3 double mutant, Foxg1 was increased in the pallium at E12.5–E14.5, but at E16.5 there was no significant increase. This signifies that miR-9 is necessary for Foxg1 regulation at the early stage for the differentiation of CR and early-born neurons. However, in the later stage, Elavl2 (RNA-binding protein) counteracts the suppression of miR-9 by binding to the U-rich region of Foxg1 3′UTR in the pallium. Both the Foxg1 dicer mutant and miR-9-2/3 double KO studies indicate that miR-9 and Foxg1 negatively regulate each other for proper pallial progenitor proliferation and CR cell differentiation (Fig. 5A).

Role of miR-9 in mice brain development.
The Foxg1 and Pax6 are expressed in the progenitors of the developing telencepahlon.87,115 Foxg1 regulates telencephalic progenitor proliferation by an autonomous mechanism as well as region-specific progenitor proliferation through the regulation of Pax6. 116 In E14.5, miR-9-2/3 double mutant, Pax6 is reduced in the pallium with an increase in Meis2. 80 Meis2 belongs to three amino acid loop extension superclass family of homeobox genes. 117 Meis2 is first observed at E10.5 in the ventrolateral telencephalon (ie, LGE and MGE), and at E12.5, it is observed in the VZ of the entire telencephalon. At E16.5, Meis2 is observed in the striatum and CP. 118 Meis2 acts as an essential Pax6 cofactor in adult olfactory bulb neurogenesis 119 and is known to regulate Pax6 expression. 120 It can be postulated that at later stages, miR-9 regulates Pax6 via Meis2 for progenitor proliferation in VZ and SVZ of the developing pallium.
However, Nr2e1 has 3′UTR binding site for miR-9; at E13.5, the Nr2e1 mRNA was upregulated in miR-9-2/3 double KO. 80 The introduction of miR-9 or let-7d into the VZ of E13.5 embryonic cortex results in the reduction in proliferative cells, and they migrate into the CP from the VZ at E15.5. The Tlx null mutant has upregulated pri-miR-9-1 and pri-miR-9-2.75,76 The let-7d inhibits TLX and promotes miR-9 expression. 76 This suggests that miR-9 helps in neuronal migration and differentiation through loop regulation of Tlx and miR-9 at early stages (E12.5–15.5). At E16.5, there is no reduction of Nr2e1 in the miR-9-2/3 double mutant. The presence of RNA-binding protein, Elav1 and Msi1, in the pallium may attenuate the suppression of Nr2e1 by miR-9. 80 Thus, miR-9 plays a role in the development of pallium by regulating Foxg1, Pax6, Nr2e1, and Meis2 (Fig. 5A).
In the E15.5 miR-9-2/3 mutant subpallium, Gsh2 and Foxg1 are increased. 80 The mechanistic role of Foxg1 is not clearly understood in subpallial progenitor proliferation and differentiation.114,121 Gsh2 is necessary for the identification of early LGE progenitor cells.122,123 The increase in Gsh2 and Foxg1 results in hyperplastic, where the proliferating cells expand into differentiating areas of the subpallium. This shows that miR-9 is necessary for proper subpallial development by regulating Gsh2 and Foxg1 expression (Fig. 5B).
Pax6 and Gsh2 have a role in specifying dorsal and ventral telencephalon, 92 and these cells by counteraction form the PSB.93,124 The Tlx/Nr2e1 is expressed in the VZ and spans along the PSB, with the exception of the dorsomedial and ventromedial regions. The Tlx mutant is similar to Pax6 mutants. The Tlx/Nr2e1 along with Pax6 regulates the establishment of PSB. 125 Similar to Tlx/Nr2e1 mutant, 125 the miR-9-2/3 double-mutant subpallium 80 shows that the development of basal ganglia including striatum and globus pallidus was suppressed, the PSB shifted dorsally, and the ventral pallidum was lost. This was substantiated by the inhibition of Pax6 via Meis2 and upregulation of Gsh2 and Nr2e1. Overall, this suggests that miR-9 is necessary for proper PSB formation by regulating Pax6, Gsh2, and Nr2e1 as well as for normal DV telencephalic development (Fig. 5C).
Role of miR-9 in telencephalon development of Xenopus
In Xenopus tropicalis, miR-9 is expressed from four variants that produce mature miR-9. The pre-miR-9 expression is similar in all the transcripts in comparison to the mature miR-9 expression. At stages 19, 30, and 36, the pre-miR-9 transcripts were observed in the forebrain, eye, midbrain, and hindbrain, but not observed in MHB (Table 2). The miR-9 expression is evident from stage 18/19 in the anterior neural plate stage of the prospective forebrain. At stage 23/24, mature miR-9 was observed in the forebrain, eye and retina, midbrain, and hindbrain with the exception of MHB.48,126 In the forebrain, miR-9 expression was widespread, in both neural progenitors (VZ) and postmitotic neurons (MZ). Morpholino (MO) miR-9 studies indicate the miR-9 was necessary for the survival of progenitors and neuronal differentiation. The miR-9 knockdown results in the inhibition of neurogenesis via apoptosis. It has been proposed that hairy1 is the main target for miR-9, where it affects proliferation via Fgf8 in the forebrain (Fig. 6A). 48 Zebrafish Her6, mammalian Hes1, Xenopus hairy1, and chicken hairy1 belong to the family of transcription factor, hairy/enhancer of split. The hairy/enhancer of split belongs to basic helix-loop-helix proteins of DNA-binding transcription factors. 127
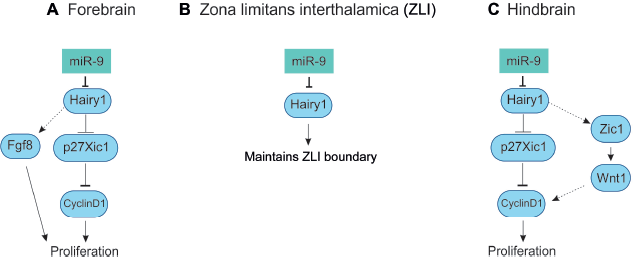
Role of miR-9 in Xenopus brain development.
Role of miR-9 in telencephalon development of zebrafish
In zebrafish, miR-9 expression is observed in the telencephalon by 24 hpf46,128 The zebrafish have seven miR-9 variants that express the same mature product, miR-9. 41 Each pri-miR-9 variant shows differential expression pattern (Table 2). At Prim6 stage 129 (25 hpf), the mature miR-9 is expressed only in the telencephalon. However, the pri-miR-9 variants 1–7 are expressed in the telencephalon with subtle differences; pri-miR-9-2 is expressed in the developing retina, pri-miR-9-4 in the posterior brain regions, and pri-miR-9-6 in the hypothalamus. 130 At 30 hpf, the miR-9 is expressed in the entire CNS, but spared MHB.46,128 At 48 hpf, all pri-miR-9 variants expressed throughout the CNS, but always spared MHB. Similarly, the mature miR-9 is expressed throughout the entire CNS. 130 In the telencephalon, miR-9 is regulated by her6 via feedback mechanism in the proliferative cells, but not in the differentiated cells. 128 The miR-9 regulates Fgf target genes (dusp6 and Pea3) in the telencephalon 46 (Fig. 7A). Limited information is available on the role of miR-9 in zebrafish telencephalon development; however, studies are needed to know how miR-9 is regulated, influences gene regulation and thereby promotes cellular changes in the zebrafish telencephalon.

Role of miR-9 in zebrafish brain development.
Role of miR-9 in telencephalon development of chick embryo
The early expression of miR-9 is observed in the area opaca at HH (Hamburger Hamilton stage) 131 4–5. At HH18, miR-9 is diversely expressed in all the developing brain vesicles and in the spinal cord. At HH20, it is expressed in the telencephalic vesicle, hindbrain, and spinal cord. From HH22–25 onward, miR-9 is expressed in the telencephalic vesicle, diencephalon, midbrain, hindbrain, and spinal cord (for detailed information, see GEISHA website).42,132 The chick has two pre-miRs (miR-9-1 and miR-9-2), which are processed by the internal dicer to produce the same mature product, miR-9. 83 So far no research reports are available on the role of miR-9 in chick telencephalon.
Diencephalon development
The diencephalon is the posterior part of the developing forebrain and develops into the thalamus and hypothalamus. 56 58 The ZLI acts as an organizer for diencephalic development. 133 136 It is a narrow strip of Shh-expressing cells in the diencephalon, transecting between prethalamus and thalamus (Fig. 8). 21 The neural tissues anterior to ZLI and in between MHB and ZLI can induce Shh. 135 The ZLI is formed at the interface between the anterior Zinc finger proteins of the Fez family and posterior homeodomain protein of Iroquois (Irx) family137,138 Like Gbx2 in the MHB, Irx3 positions the ZLI in the posterior forebrain. 139 The Six3 and Irx3 mutually repress each other and form the ZLI boundary. 140

Gene expression in developing chick embryo.
Role of miR-9 in the diencephalon development of mice
The miR-9 is not expressed in the diencephalon of E11.5 mouse. 33 No reports are available with regard to gene regulation by miR-9 expression in diencephalon.
Role of miR-9 in the diencephalon development of Xenopus
In Xenopus tropicalis, miR-9 is not expressed in the MHB and ZLI, where hairy1 and Fgf8 are expressed. The miR-9 MO and hairy1 target protection assay leads to the expansion of hairy1- and Fgf8-positive domain, 48 which indicates that miR-9 inversely regulates hairy1 expression via non-cell autonomous signaling pathway in ZLI (Fig. 6B). Another study has shown that hes1 in mouse, similar to hairy1 in Xenopus, inversely regulates miR-9 biogenesis. 49
Role of miR-9 in the diencephalon development of zebrafish
The miR-9 expression is observed in the diencephalon.46,128,141 However, there are no research reports available on gene regulation by miR-9.
Role of miR-9 in the diencephalon development of chick
There are no reports on the role of miR-9 on chick embryonic diencephalon. However, the miR-9 expression is observed in the diencephalon 42 from HH22 onward.
Mesencephalon/midbrain development
The M HB plays an important role for the AP development of midbrain. The midbrain consists of two main components along the DV axis: the dorsal tectum and the ventral tegmentum. The tectum develops from the alar plate, and the tegmentum develops from the basal plate. 142 The MHB arises from the morphological constriction between the midbrain and hindbrain. The organization of MHB has been divided into four stages: (1) positioning and establishment, (2) induction, (3) maintenance, 55 and (4) morphogenesis. 143 The Otx2 and Gbx2 are the critical factors for position and establishment of the MHB.144,145 These signals refine the position of MHB.146–148 The Otx2 and Gbx2 repress each other, from where the Fgf8 is expressed. 149 In zebrafish, her5 has been postulated to establish the midbrain and hindbrain cells, 150 and Pou2 is necessary for the establishment of MHB. 151 The induction of MHB is regulated by the members of Pax, Fgf, Wnt, and En families, where Fgf8 plays a crucial role. 152 In Xenopus, Xiro along with Otx2/Gbx2 is necessary for the induction of Fgf8 at MHB. 153 The maintenance of MHB is achieved by interdependent regulation of genes, namely Pax2/5, Wnt1, En1/2, and Fgf8 (Fig. 8A and B).55,143 Recently, it was shown that the maintenance and morphogenesis of MHB in zebrafish was governed by the transcription factor Grhl2b (grainy head-like 2b). 154
Role of miR-9 in the mesencephalon/midbrain development of mice
Foxg1 is also expressed in the MHB at E10.5. 98 However, there is no report on the role of miRNAs in the MHB of Foxg1-cre dicer KO.
The observation in Emx1-cre dicer KO shows that there is no change in midbrain structure and miR-9 expression in the mesencephalon, 96 which is obvious since Emx1 expression is observed only in the telencephalon. 26
The Pax2-cre dicer KO and Wnt1-cre dicer KO have similar impaired growth in the midbrain and hindbrain structures.37,155 These Pax2-cre mice died at E18.5. Pax2 is expressed in the MHB 156 and Wnt1 is expressed in the midbrain at E8. 157 The Wnt1-cre dicer KO shows several face malformation. The size of the tectum, rostral (superior colliculus), and caudal (inferior colliculus) tectum was clearly reduced, and the cerebellum was ablated in these KOs. The AP pattern was unaffected. The expression of genes such as Otx2, Gbx2, Wnt1, FgfS, and Lmx1b was not affected, and the MHB was correctly positioned. In comparison to the wild type, the miR-9, miR-124, and miR-218 were greatly eliminated in tectum, ventral midbrain, and rostral hindbrain of the Wnt1-cre dicer mutant. The dicer mutant points out that the TH expression was markedly reduced in the ventral midbrain. The TH and Nurr1-positive cells failed to properly differentiate into mDA (mesencephalic dopaminergic) neurons. 37 This shows that dicer mutant have mDA precursor cells, but needs miRNAs to differentiate into proper mDA neurons. It appears that the miRNAs (miR-9, miR-124, miR-218) eliminated in Wnt1-cre dicer KOs are necessary for the proper development of mDA neurons.
Role of miR-9 in the mesencephalon/midbrain development in Xenopus
At stages 30–36, the miR-9 is expressed in the midbrain as well as in the hindbrain with the exception of MHB. The miR-9 expression in the midbrain and hindbrain is restricted to the proliferating progenitor cells of the VZ. The primary miR-9 (miR-9a-1, miR-9a-2, miR-9-3) is expressed in the midbrain (Table 2).48,126 The hairy1 is expressed in the ventral midbrain that coincides with the expression of miR-9a-1. 48 It might be possible that miR-9 regulates hairy1 for neurogenesis and differentiation in the Xenopus midbrain.
Role of miR-9 in the mesencephalon/midbrain development in zebrafish
In zebrafish, miR-9 is not expressed in the MHB.46,128 The gain of function (GOF) of miR-9 experiment shows that overexpression leads to the loss of MHB and MHB marker gene expression. The GOF and loss of function (LOF) studies show that miR-9 has influence on the Fgf signaling pathway genes (FgfS, Fgfrl, and canopy1) and its target genes such as Pea3 and dusp6 are downregulated. Experimental evidence proves that Her5, her9, canopy1, fgf8, and fgfr1 are in vivo targets of miR-9. Independent of Fgf signaling interaction, miR-9 also promotes midbrain-hindbrain neurogenesis and neuronal differentiation in vivo via regulating her (her5 and her9) genes (Fig. 7B). 46 Overall, miR-9 is involved in neurogenesis, Fgf signaling, and maintenance of MHB.
Role of miR-9 in the mesencephalon/midbrain development in chick
The miR-9 is expressed in the midbrain; 42 however, no research reports are available on the role of miR-9 in gene regulation of midbrain and MHB genes.
Hindbrain development
The developing hindbrain is formed from the rudimentary segments of rhombomeres, where the cells move freely between rhombomeres before the boundary formation and after the boundary formation they expands within the boundaries. 158 The development of hindbrain depends on the retinoic acid gradient signals, tyrosine kinases, and several transcription factors along the rhombomere boundaries. 159 161 In zebrafish, several Wnt genes are expressed in the boundaries between the rhombomeres.162,163 In chick, Hox genes play a main role in the formation of rhombomere boundaries.159,160
Role of miR-9 in the hindbrain development of mice
In Wnt1-cre dicer mutant, notable defects were observed in the hindbrain that included the ablation of cerebellum. The miR-9 along with miR-124 and miR-218 expression was ablated in rostral hindbrain of the Wnt1-cre dicer mutant. There was no difference in the expression of Gbx2 expression in the hindbrain. The mitotic progenitors in the hindbrain were similar to the control, 37 indicating dicer KO does not affect proliferation in mice hindbrain. Further research is needed to pinpoint the role of miR-9 in the developing hindbrain.
In hGFAP-cre dicer mutant, the activity was observed as early as E13.5 at the rhombic lip of the cerebellar plate and in postnatal cerebellar cells including the Bergmann glia. Defective cerebellar foliation was observed at E16.5. In the Bergmann glia, miR-9 is needed for the maintenance of these cells via Notch1 signaling. 30
Role of miR-9 in the hindbrain development of Xenopus
Bonev et al reported that in Xenopus, miR-9 expression was restricted to the Sox3 (neural progenitor marker)-positive domain, ie, VZ cells of the developing hindbrain. The miR-9 is involved in the hindbrain development through hairy1 on proliferation via Wnt signaling (Fig. 6C). The MO experiment also shows that miR-9 is necessary for the differentiation of neuron. In comparison to forebrain, miR-9 is necessary for limiting progenitor proliferation in hindbrain and promotes the onset of neurogenesis. 48
Role of miR-9 in the hindbrain development in zebrafish
In zebrafish, at 30 hpf, the miR-9 expression is observed in the hindbrain with the absence of MHB and rhombomere boundary.41,46,128 In the hindbrain, radial glial cells in the VZ express miR-9, where the expression is segmentally pattern and stronger near the rhombomere boundaries. The miR-9 expression is also observed in the neurons of the MZ. The miR-9 regulates hindbrain neurogenesis by maintaining neural progenitor cells through zic5 and her6 and neuronal differentiation by targeting elavl3/HuC (Fig. 7C). The miR-9 was positively regulated by Notch signaling and negatively regulated by Notch target Her6. 128 The miR-9 inhibits Her9, thereby regulates neuronal progenitor maintenance. 46 Overall, miR-9 is involved in proper neuronal progenitor maintenance and neurogenesis in zebrafish.
Role of miR-9 in the hindbrain development of chick
From HH20 onward, the miR-9 expression is observed in the hindbrain. At HH24, it is strongly expressed in the margin of the roof plate and absent in the floor plate. 42 However, there are no research reports on gene regulation by miR-9 in the hindbrain.
Spinal cord development
The spinal cord develops from the posterior neural tube by the induction of cues from the dorsal ectoderm. The regional differentiation occurs along the DV and rostrocaudal axis. The retinoid synthesis by RALDH2 (retinaldehyde dehydrogenase 2) is critical for establishing the distinction of spinal cord neural cells. 164 The dorsally positioned neurons are induced by BMP signaling, and the ventrally positioned neurons are induced by the graded expression of SHH. The Hox genes govern the patterning of spinal cord. 165 The motor neurons (MNs) and interneurons (IN) are generated by the graded action of SHH. The graded SHH repress the class I homeodomain proteins (Pax6/7, Irx3, DBX1/2) and class II proteins (Nkx2.2 and Nkx6.1) for the generation of five distinct class of neurons (ventral IN–-V0-3 and MN) from the ventral progenitor cells along DV axis of the ventral neural tube. 166 The combinatorial action of Nkx6.1, Nkx2.2, and Irx3 restricts the generation of MN to a single progenitor domain.164,166 The median motor column (MMC) neurons innervate axial muscles and lateral motor column (LMC) innervate limb muscles. 164
Role of miR-9 in the spinal cord development of chick
The miR-9 is expressed in the VZ of developing chick spinal cord from HH11 to HH29.42,83 In the MN, the miR-9 expression was low at HH20, increased at HH24 and undetectable at HH29. At HH24, miR-9 overlaps with the MN markers (FoxP1 and Isl1/2) in the LMC; however, they do not overlap with MN markers (Lhx3, HB9) of the MMC neurons. The overexpression of miR-9 results in switching of LMC into MMC neurons by repressing the production of LMC neurons by downregulating FoxP1 and Isl1/2 and promoting the differentiation of MMC neurons by upregulating Lhx3 and HB9. The overlapping expression of miR-9 and FoxP1 along with experimental evidence shows that miR-9 regulates FoxP1 expression in LMC in a dose-dependent manner. In chick, preganglionic motor column (PGC) is positioned along the thoracic level innervate axons to the sympathetic target. The overexpression of miR-9 shows that there is a reduced expression of PGC MN marker, Smad and Isl1/2. 83 In the LMC, the GoF-miR-9 shows that FoxP1 is reduced and vice versa in the LoF-miR-9. Likewise, the reduction and increase in FoxP1 are inversely proportional to the Lhx3 MN marker in the LMC. 167 Overall, miR-9 finely tunes FoxP1 expression for normal LMC (Fig. 9) and PGC neuron specification.
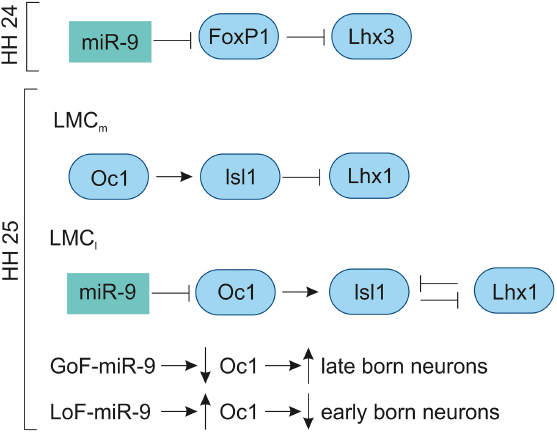
Role of miR-9 in chick spinal cord development.
At HH25, the miR-9 expression is increased in the intermediate and VZs of the spinal cord. At HH28, miR-9 is expressed in specific subpopulations of the MNs; the MN migration is completed by this stage, where the MMC (marker: Lhx3), medial LMC (marker: Isl1/FoxP1), and lateral LMC (marker: Lhx1/Hb9) can be clearly distinguished. The transcription factor OC1 (Onecut 1) is expressed in the lateral aspect of the ventral horn. At HH25, the OC1 is co-localized with Isl1/Lhx3 of early-born neurons of MMC, in the medial LMC with Isl1/FoxP1 and in the lateral LMC with Lhx1/Hb9. At HH27, the lateral MMC shows less OC1 compared to the medial MMC, while the miR-9 is reversed in strength. The medial LMC has strong OC1 expression with no miR-9, and lateral LMC has strong miR-9 with reduced OC1. The GoF-miR-9 results in the reduction in lateral MMC and medial LMC, while the medial MMC and lateral LMC are increased. The opposite reactions are observed in LoF-miR-9. The GoF-miR-9 results in the downregulation of OC1 and an increase in late-born neurons. Likewise, LOF-miR-9 results in the upregulation of OC1 and an increase in early-born neurons. 168 Overall, miR-9 regulates Onecut 1 to promote the switching of early-born to late-born MN population (Fig. 9).
Conclusion
In the developing brain, miR-9 influences gene expression, proliferation, neurogenesis, maturation, migration, and differentiation in a spatiotemporal pattern. The studies point out that miR-9 is expressed in neuronal cells which are proliferative and differentiated cells. The endogenous miR-9 expression pattern also differs from species to species; in zebrafish and Xenopus, miR-9 is excluded from the MHB, but in mice and chicken there is no such distinction in the expression pattern. The miR-9 is necessary for the maintenance of boundaries: PSB, ZLI, and MHB.
Although the studies on dicer KO mice show the lack of miR-9, one cannot ignore the plethora of other miRNAs involvement in brain development. At the same time, we could not reject the experimental evidence showing the necessity of miR-9 and its role in vertebrate brain development. The other dicer KOs have similar developmental defects in telencephalon in comparison to miR-9-2/3 double mutant. Combining these facts with other experimental evidence like GOF/LOF, we can conclude that miR-9 plays a role in the vertebrate telencephalon development in mice. One among such facts is the regulation of CR neurons by miR-9. The miR-9 modulates the generation, maturation, differentiation, and migration of CR neurons in the developing telencephalon. The miR-9 is also governed by several genes, like Notch, Hes/hairy1, Tlx, Dlx5, etc. There are also feedback loop mechanisms for Tlx, Foxg1, and Foxp1 with miR-9 that regulates cellular events within the developing vertebrate brain.
Several hairy/enhancer of split genes (Xenopus hairy1, zebrafish her5, her9, her6) are regulated by miR-9. Most of these are proneural genes involve in the regulation of progenitor pools. In these studies, we found that miR-9 regulates proliferation and differentiation via targeting her genes. Even within the species, the miR-9 is differentially regulated. For example, hairy1 mediates the effects of miR-9 on proliferation through Fgf8 signaling in the forebrain and via Wnt signaling in the hindbrain.
Further studies are required to completely understand the role of miR-9 in vertebrate brain development. For instance, miR-9 expression has been observed in the chick diencephalon from HH22, and reports on the role of miR-9 in the embryonic diencephalon are lacking. Similarly, miR-9 expression has been reported in the zebrafish diencephalon, but data on its gene regulatory role are not yet available. In the case of the chick midbrain and hindbrain, no data are available on the role of miR-9 in both these regions. This situation is also prevalent in the case of the chick telencephalon, allowing much more scope for understanding the role of miR-9. This may also provide an insight into the role played by miR-9 from an evolutionary standpoint. To conclude, miR-9 is necessary for the development of the embryonic brain.
Author Contributions
Conceived and designed the review, analyzed the data, contributed equally in writing the manuscript: BR and AAPA. Agree with manuscript results and conclusions: BR and AAPA. Drawn figures for the manuscript using graphic software: BR. Jointly developed the structure and arguments for the paper: BR and AAPA. Made critical revisions and approved final version: AAPA. All authors reviewed and approved of the final manuscript.